
Differential Performance between Two Timber Species in Forest Logging Gaps and in Plantations in Central Africa

Abstract
:1. Introduction
2. Methods
2.1. Study Area
2.2. Study Species

2.3. Silvicultural Experiments
2.3.1. Enrichment in Logging Gaps
2.3.2. Plantations
2.4. Data Analysis
3. Results and Discussion
3.1. Differential Performance between the Two Species


{kind=link}
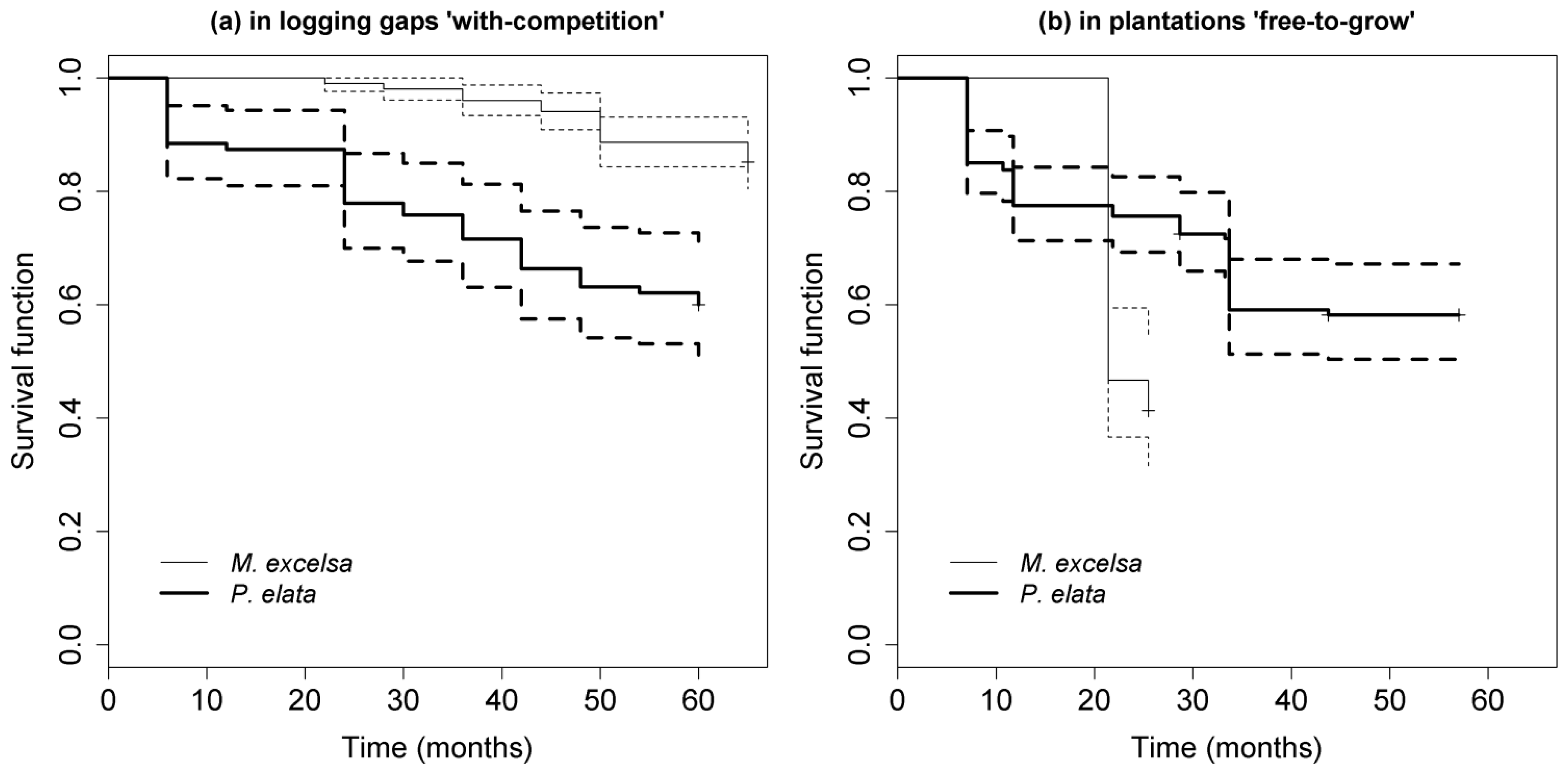{kind=link}
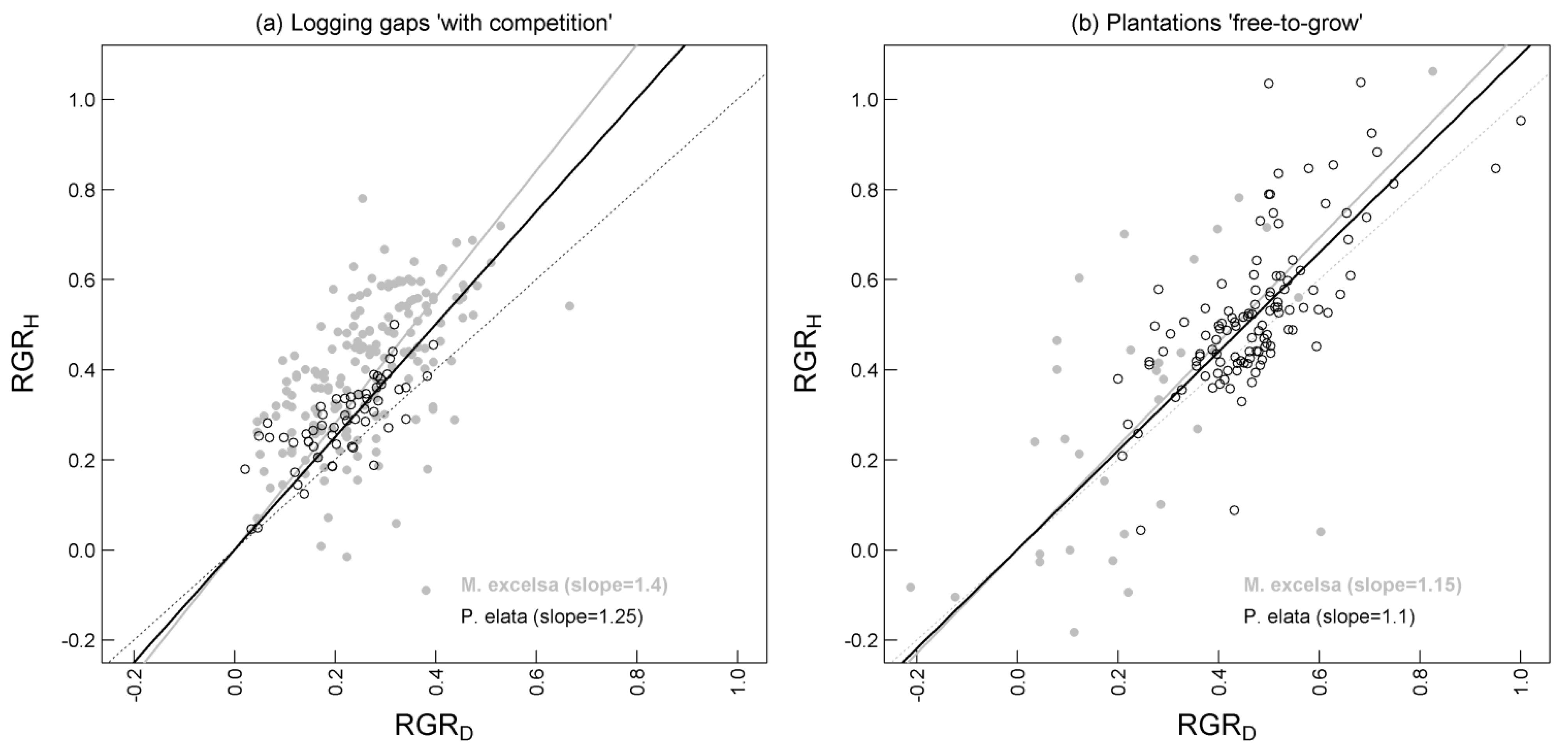{kind=link}
| Growth Variable | Milicia excelsa | Pericopsis elata | T Value | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | N | Mean | SD | N | |||
| (a) Logging Gaps-with-Competition | ||||||||
| Height increment (cm·yr−1) | 39.96 a | 30.76 | 172 | 45 a | 23 | 57 | −1.29 | 0.20 |
| Diameter increment (mm·yr−1) | 2.87 a | 2.02 | 172 | 2.79 a | 1.37 | 57 | 0.32 | 0.75 |
| RGRH (cm cm−1·yr−1) | 0.40 a | 0.16 | 172 | 0.29 b | 0.09 | 57 | 6.81 | <0.0001 |
| RGRD (mm mm−1·yr−1) | 0.26 a | 0.11 | 172 | 0.21 b | 0.09 | 57 | 3.41 | 0.0009 |
| (b) Plantations-Free-to-Grow | ||||||||
| Height increment (cm·yr−1) | 19.96 a | 20.87 | 33 | 86.43 b | 42.96 | 115 | −12.29 | <0.0001 |
| Diameter increment (mm·yr−1) | 1.70 a | 1.78 | 33 | 9.08 b | 4.56 | 115 | −14.03 | <0.0001 |
| RGRH (cm cm−1·yr−1) | 0.31 a | 0.31 | 33 | 0.53 b | 0.17 | 115 | −3.83 | 0.0005 |
| RGRD (mm mm−1·yr−1) | 0.24 a | 0.21 | 33 | 0.47 b | 0.13 | 115 | −6.15 | <0.0001 |
3.2. Tree Architecture in Logging Gaps and in Plantations

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Wasseige, C.; Devers, D.; de Marcken, P.; Eba’a Atyi, R.; Nasi, R.; Mayaux, P. (Eds.) Les Forêts du Bassin du Congo—Etat des Forêts 2008; Office des publications de l’UE: Luxembourg, Luxembourg, 2009.
- De Wasseige, C.; de Marcken, P.; Bayol, N.; Hiol Hiol, F.; Mayaux, P.; Desclée, B.; Billand, A.; Nasi, R. (Eds.) Les Forêts du Bassin du Congo—Etat des Forêts 2010; Office des publications de l’UE: Luxembourg, Luxembourg, 2012.
- Nasi, R.; Billand, A.; van Vliet, N. Managing for timber and biodiversity in the Congo Basin. For. Ecol. Manag. 2012, 268, 103–111. [Google Scholar]
- Ruiz Pérez, M.; Ezzine de Blas, D.; Nasi, R.; Sayer, J.A.; Sassen, M.; Angoué, C.; Gami, N.; Ndoye, O.; Ngono, G.; Nguinguiri, J.C. Logging in the Congo Basin: A multi-country characterization of timber companies. For. Ecol. Manag. 2005, 214, 221–236. [Google Scholar]
- Hall, J.S.; Harris, D.J.; Medjibe, V.; Ashton, P.M.S. The effects of selective logging on forest structure and tree species composition in a Central African forest: Implications for management of conservation areas. For. Ecol. Manag. 2003, 183, 249–264. [Google Scholar] [CrossRef]
- Fargeot, C.; Forni, É.; Nasi, R. Réflexions sur l’aménagement des forêts de production dans le bassin du Congo. Bois For. Trop. 2004, 281, 19–34. [Google Scholar]
- Karsenty, A.; Gourlet-Fleury, S. Assessing sustainability of logging practices in the Congo Basin’s managed forests: The issue of commercial species recovery. Ecol. Soc. 2006, 11, 26. [Google Scholar]
- Bongers, F.; Poorter, L.; Rompaey, R.S.A.R.; Parren, M.P.E. Distribution of twelve moist forest canopy tree species in Liberia and Côte d’Ivoire: Response curves to a climatic gradient. J. Veg. Sci. 1999, 10, 371–382. [Google Scholar] [CrossRef]
- Hawthorne, W.D. Ecological Profiles of Ghanaian Forest Trees; Tropical Forestry Papers; Oxford Forestry Institute, Department of Plant Sciences, University of Oxford: Oxford, UK, 1995. [Google Scholar]
- Doucet, J.-L. L’alliance délicate de la gestion forestière et de la biodiversité dans les forêts du centre du Gabon. Ph.D. Thesis, Faculté Universitaire des Sciences Agronomiques de Gembloux, Gembloux, Belgique, 2003. [Google Scholar]
- Makana, J.-R.; Thomas, S.C. Effects of Light Gaps and Litter Removal on the Seedling Performance of Six African Timber Species. Biotropica 2005, 37, 227–237. [Google Scholar] [CrossRef]
- Sist, P.; Brown, N. Silvicultural intensification for tropical forest conservation: A response to Fredericksen and Putz. Biodivers. Conserv. 2004, 13, 2381–2385. [Google Scholar] [CrossRef]
- Van Gemerden, B.S.; Olff, H.; Parren, M.P.E.; Bongers, F. The pristine rain forest? Remnants of historical human impacts on current tree species composition and diversity. J. Biogeogr. 2003, 30, 1381–1390. [Google Scholar] [CrossRef]
- Engone Obiang, N.L.; Ngomanda, A.; Hymas, O.; Chézeaux, É.; Picard, N. Diagnosing the demographic balance of two light-demanding tree species populations in central Africa from their diameter distribution. For. Ecol. Manag. 2014, 313, 55–62. [Google Scholar] [CrossRef]
- Grogan, J.; Landis, R.M.; Ashton, M.S.; Galvão, J. Growth response by big-leaf mahogany (Swietenia macrophylla) advance seedling regeneration to overhead canopy release in southeast Pará, Brazil. For. Ecol. Manag. 2005, 204, 399–412. [Google Scholar] [CrossRef]
- Coates, K.D.; Burton, P.J. A gap-based approach for development of silvicultural systems to address ecosystem management objectives. For. Ecol. Manag. 1997, 99, 337–354. [Google Scholar] [CrossRef]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of Degraded Tropical Forest Landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M. Technical and financial analysis of enrichment planting in logging gaps as a potential component of forest management in the eastern Amazon. For. Ecol. Manag. 2008, 255, 866–879. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Palik, B.J.; Dumroese, R.K. Contemporary forest restoration: A review emphasizing function. For. Ecol. Manag. 2014, 331, 292–323. [Google Scholar] [CrossRef]
- Keefe, K.; Schulze, M.D.; Pinheiro, C.; Zweede, J.C.; Zarin, D. Enrichment planting as a silvicultural option in the eastern Amazon: Case study of Fazenda Cauaxi. For. Ecol. Manag. 2009, 258, 1950–1959. [Google Scholar] [CrossRef]
- Korpelainen, H.; Ådjers, G.; Kuusipalo, J.; Nuryanto, K.; Otsamo, A. Profitability of rehabilitation of overlogged dipterocarp forest: A case study from South Kalimantan, Indonesia. For. Ecol. Manag. 1995, 79, 207–215. [Google Scholar] [CrossRef]
- Ashton, M.S.; Goodale, U.M.; Bawa, K.S.; Ashton, P.S.; David Neidel, J. Restoring working forests in human dominated landscapes of tropical South Asia: An introduction. For. Ecol. Manag. 2014, 329, 335–339. [Google Scholar] [CrossRef]
- Doucet, J.-L.; Kouadio, Y.L.; Monticelli, D.; Lejeune, P. Enrichment of logging gaps with moabi (Baillonella toxisperma Pierre) in a Central African rain forest. For. Ecol. Manag. 2009, 258, 2407–2415. [Google Scholar] [CrossRef]
- Brienen, R.J.; Zuidema, P.A. Lifetime growth patterns and ages of Bolivian rain forest trees obtained by tree ring analysis. J. Ecol. 2006, 94, 481–493. [Google Scholar] [CrossRef]
- Goodale, U.M.; Ashton, M.S.; Berlyn, G.P.; Gregoire, T.G.; Singhakumara, B.M.P.; Tennakoon, K.U. Disturbance and tropical pioneer species: Patterns of association across life history stages. For. Ecol. Manag. 2012, 277, 54–66. [Google Scholar] [CrossRef]
- Brunck, F.; Grison, F.; Maitre, H.F. L’Okoumé: Monographie; Centre Technique Forestier Tropical (CIRAD-Forêt): Montpellier, France, 1990. [Google Scholar]
- Koumba Zaou, P.; Nze Nguema, S.; Mapaga, D.; Deleporte, P. Croissance de 13 essences de bois d’œuvre plantées en forêt gabonaise. Bois For. Trop. 1998, 256, 21–33. [Google Scholar]
- Ndongo, P.-A.O.; Peltier, R.; Linjouom, I.; Louppe, D.; Smektala, G.; Beligné, V.; Njoukam, R.; Tieche, B.; Temgoua, L. Plantations de bois d’oeuvre en zone équatoriale africaine: Cas de l’arboretum de l’Enef de Mbalmayo au sud du Cameroun. Bois For. Trop. 2009, 299, 37–48. [Google Scholar]
- Bourland, N.; Kouadio, Y.L.; Fétéké, F.; Lejeune, P.; Doucet, J.-L. Ecology and management of Pericopsis elata (Harms) Meeuwen (Fabaceae) populations: A review. Biotechnol. Agron. Société Environ. Biotechnol. Agron. Soc. Environ. BASE 2012, 16, 486–498. [Google Scholar]
- Bourland, N.; Kouadio, L.; Lejeune, P.; Sonké, B.; Philippart, J.; Daïnou, K.; Fétéké, R.F.; Doucet, J.L. Ecology of Pericopsis elata (Fabaceae), an Endangered Timber Species in Southeastern Cameroon. Biotropica 2012, 44, 840–847. [Google Scholar] [CrossRef]
- Baraloto, C.; Goldberg, D.E.; Bonal, D. Performance trade-offs among tropical tree seedlings in contrasting microhabitats. Ecology 2005, 86, 2461–2472. [Google Scholar] [CrossRef]
- Kobe, R.K.; Pacala, S.W.; Silander, J.A., Jr.; Canham, C.D. Juvenile tree survivorship as a component of shade tolerance. Ecol. Appl. 1995, 5, 517–532. [Google Scholar] [CrossRef]
- Walters, M.B.; Reich, P.B. Are shade tolerance, survival, and growth linked? Low light and nitrogen effects on hardwood seedlings. Ecology 1996, 77, 841–853. [Google Scholar] [CrossRef]
- Goodale, U.M.; Berlyn, G.P.; Gregoire, T.G.; Tennakoon, K.U.; Ashton, M.S. Differences in Survival and Growth among Tropical Rain Forest Pioneer Tree Seedlings in Relation to Canopy Openness and Herbivory. Biotropica 2014, 46, 183–193. [Google Scholar] [CrossRef]
- Coomes, D.A.; Grubb, P.J. A comparison of 12 tree species of Amazonian caatinga using growth rates in gaps and understorey, and allometric relationships. Funct. Ecol. 1998, 12, 426–435. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Jones, A.; Breuning-Madsen, H.; Brossard, M.; Dampha, A.; Deckers, J.; Dewitte, O.; Gallali, T.; Hallett, S.; Jones, R.; Kilasara, M.; et al. Soil Atlas of Africa; Publications Office of the European Union: Luxembourg, Luxembourg, 2013. [Google Scholar]
- De Namur, C. Aperçu sur la végétation de l’Afrique centrale atlantique. Paysages Quat. L’Afrique Cent. Atl. ORSTOM Paris 1990, 60–67. [Google Scholar]
- Letouzey, R. Notice de la Carte Phytogéographique du Cameroun au 1/500,000; Institut de la carte internationale de la végétation: Toulouse, France, 1985. [Google Scholar]
- Royal Botanic Gardens Seed Information Database (SID). Version 7.1. Available online: http://data.kew.org/sid/ (accessed on 15 December 2014).
- Daïnou, K.; Laurenty, E.; Mahy, G.; Hardy, O.J.; Brostaux, Y.; Tagg, N.; Doucet, J.-L. Phenological patterns in a natural population of a tropical timber tree species, Milicia excelsa (Moraceae): Evidence of isolation by time and its interaction with feeding strategies of dispersers. Am. J. Bot. 2012, 99, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Data from: Towards a worldwide wood economics spectrum. Dryad Digital Repository. Dryad Digital Repository. Available online: http://datadryad.org/resource/doi:10.5061/dryad.234 (accessed on 15 December 2014).
- Ouédraogo, D.-Y.; Fayolle, A.; Daïnou, K.; Demaret, C.; Bourland, N.; Lagoute, P.; Doucet, J.-L. Enrichment of Logging Gaps with a High Conservation Value Species (Pericopsis elata) in a Central African Moist Forest. Forests 2014, 5, 3031–3047. [Google Scholar] [CrossRef]
- Harrell, F.E. Regression Modeling Strategies: With Applications to Linear Models, Logistic Regression, and Survival Analysis; Springer-Verlag: New York, NY, USA, 2001. [Google Scholar]
- Therneau, T.M. A Package for Survival Analysis in S. Available online: http://CRAN.R-project.org/package=survival (accessed on 15 December 2014).
- Agyeman, V.K.; Swaine, M.D.; Thompson, J. Responses of tropical forest tree seedlings to irradiance and the derivation of a light response index. J. Ecol. 1999, 87, 815–827. [Google Scholar] [CrossRef]
- Kyereh, B.; Swaine, M.D.; Thompson, J. Effect of light on the germination of forest trees in Ghana. J. Ecol. 1999, 87, 772–783. [Google Scholar] [CrossRef]
- Poorter, L. Growth responses of 15 rain-forest tree species to a light gradient: The relative importance of morphological and physiological traits. Funct. Ecol. 1999, 13, 396–410. [Google Scholar] [CrossRef]
- Denslow, J.S. Gap partitioning among tropical rainforest trees. Biotropica 1980, 12, 47–55. [Google Scholar] [CrossRef]
- Wagner, M.R.; Cobbinah, J.R.; Bosu, P.P. Forest Entomology in West Tropical Africa: Forest Insects of Ghana; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Bosu, P.P.; Cobbinah, J.R.; Nichols, J.D.; Nkrumah, E.E.; Wagner, M.R. Survival and growth of mixed plantations of Milicia excelsa and Terminalia superba 9 years after planting in Ghana. For. Ecol. Manag. 2006, 233, 352–357. [Google Scholar] [CrossRef]
- Nichols, J.D.; Ofori, D.A.; Wagner, M.R.; Bosu, P.; Cobbinah, J.R. Survival, growth and gall formation by Phytolyma lata on Milicia excelsa established in mixed-species tropical plantations in Ghana. Agric. For. Entomol. 1999, 1, 137–141. [Google Scholar] [CrossRef]
- King, D.A. Allometry of Saplings and Understorey Trees of a Panamanian Forest. Funct. Ecol. 1990, 4, 27–31. [Google Scholar] [CrossRef]
- Vanclay, J.K. Assessing site productivity in tropical moist forests: A review. For. Ecol. Manag. 1992, 54, 257–287. [Google Scholar] [CrossRef]
- Midgley, J.J. Do mixed-species mixed-size indigenous forests also follow the self-thinning line? Trends Ecol. Evol. 2001, 16, 661–662. [Google Scholar] [CrossRef]
- Parrotta, J.A.; Turnbull, J.W.; Jones, N. Catalyzing native forest regeneration on degraded tropical lands. For. Ecol. Manag. 1997, 99, 1–7. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fayolle, A.; Ouédraogo, D.-Y.; Ligot, G.; Daïnou, K.; Bourland, N.; Tekam, P.; Doucet, J.-L. Differential Performance between Two Timber Species in Forest Logging Gaps and in Plantations in Central Africa. Forests 2015, 6, 380-394. https://doi.org/10.3390/f6020380
Fayolle A, Ouédraogo D-Y, Ligot G, Daïnou K, Bourland N, Tekam P, Doucet J-L. Differential Performance between Two Timber Species in Forest Logging Gaps and in Plantations in Central Africa. Forests. 2015; 6(2):380-394. https://doi.org/10.3390/f6020380
Chicago/Turabian StyleFayolle, Adeline, Dakis-Yaoba Ouédraogo, Gauthier Ligot, Kasso Daïnou, Nils Bourland, Patrice Tekam, and Jean-Louis Doucet. 2015. "Differential Performance between Two Timber Species in Forest Logging Gaps and in Plantations in Central Africa" Forests 6, no. 2: 380-394. https://doi.org/10.3390/f6020380
APA StyleFayolle, A., Ouédraogo, D.-Y., Ligot, G., Daïnou, K., Bourland, N., Tekam, P., & Doucet, J.-L. (2015). Differential Performance between Two Timber Species in Forest Logging Gaps and in Plantations in Central Africa. Forests, 6(2), 380-394. https://doi.org/10.3390/f6020380