
Effects of Topographic and Soil Factors on Woody Species Assembly in a Chinese Subtropical Evergreen Broadleaved Forest

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description

2.2. Plot Installation and Investigation
2.3. Topographic and Soil Data Collection
2.4. Data Analysis
2.4.1. Stand Characteristics
2.4.2. Spatial Patterns and Associations
2.4.3. Topography and Soil Contribution
3. Results
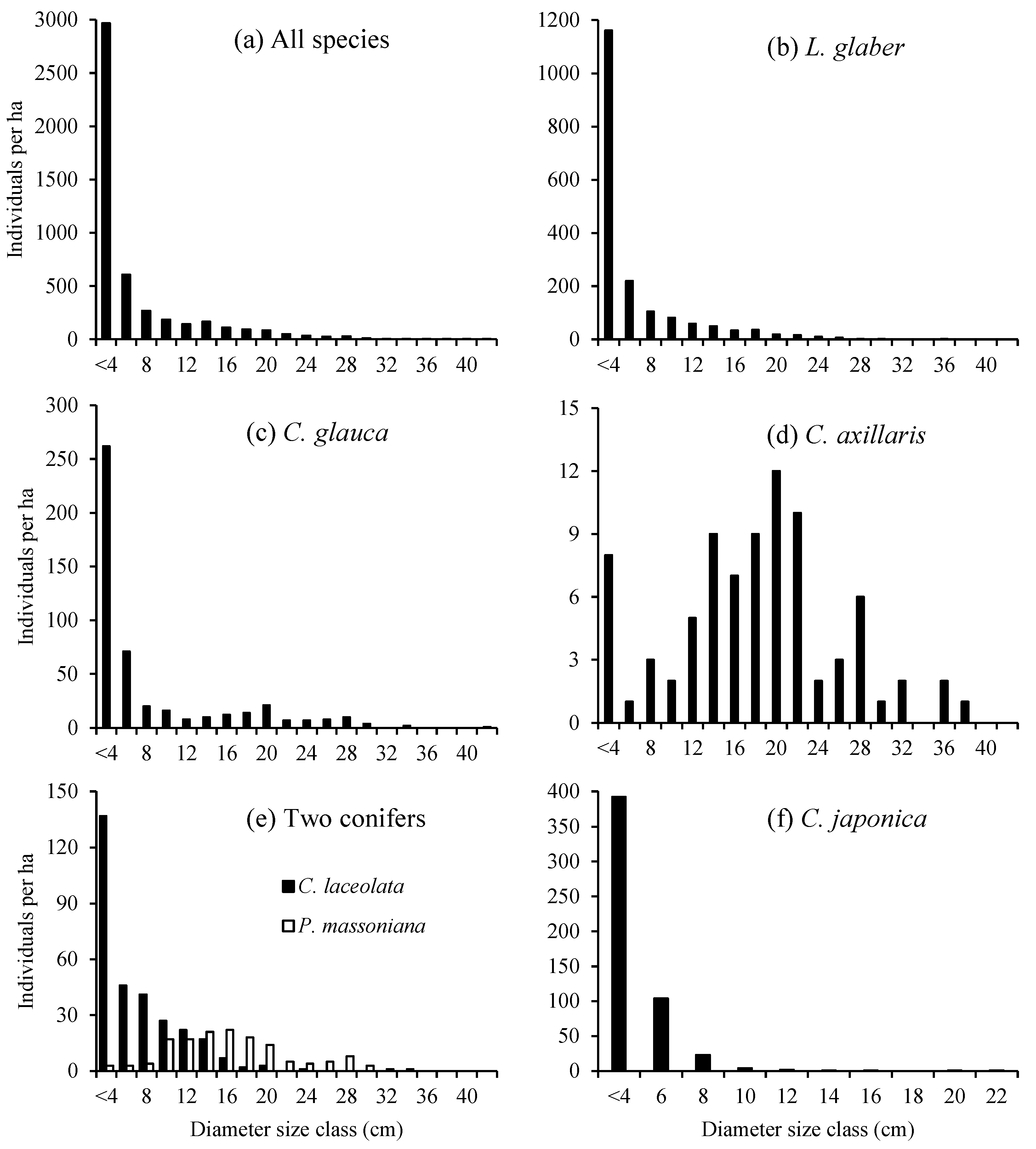
3.1. Stand Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Density (Stems ha−1) | BA (m2 ha−1) | Height (m) | dbh (cm) | Frequency (%) | RIV |
|---|---|---|---|---|---|---|
| L. glaber | 1802 | 6.4 | 5.5 | 5.4 | 94 | 25.3 |
| C. glauca | 473 | 3.8 | 5.5 | 5.8 | 62 | 11.1 |
| P. massoniana | 146 | 3.1 | 12.6 | 16.5 | 45 | 7.2 |
| C. japonica | 529 | 0.6 | 3.7 | 3.3 | 57 | 6.6 |
| C. lanceolata | 303 | 1.2 | 5.8 | 6.0 | 51 | 5.7 |
| C. axillaris | 83 | 2.3 | 11.5 | 18.4 | 38 | 5.3 |
| Loropetalum chinense (R. Br.) Oliver | 185 | 0.1 | 3.3 | 2.5 | 47 | 3.2 |
| Eurya muricata Dunn | 181 | 0.1 | 2.7 | 2.1 | 46 | 3.1 |
| Phyllostachys edulis (Carrière) J. Houz. | 94 | 1.1 | 12.2 | 12.2 | 9 | 2.5 |
| C. henryi | 26 | 1.2 | 12.1 | 22.5 | 16 | 2.5 |
| Other 63 species | 975 | 3.0 | 4.6 | 4.5 | 95 | 27.5 |
| Total 73 species | 4797 | 22.9 | 6.1 | 5.5 | 100 | 100.0 |
| Species | Density (Stems ha−1) | BA (m2 ha−1) | Height (m) | dbh (cm) | Frequency (%) | RIV |
|---|---|---|---|---|---|---|
| L. glaber | 641 | 5.9 | 8.2 | 10.0 | 90 | 26.4 |
| C. glauca | 211 | 3.7 | 8.3 | 10.8 | 50 | 12.6 |
| P. massoniana | 143 | 3.1 | 12.6 | 16.6 | 45 | 10.2 |
| C. lanceolata | 166 | 1.1 | 7.6 | 8.7 | 42 | 7.4 |
| C. axillaris | 75 | 2.3 | 12.0 | 19.2 | 37 | 7.1 |
| C. japonica | 137 | 0.4 | 5.2 | 5.9 | 34 | 5.2 |
| P. edulis | 90 | 1.1 | 12.5 | 12.5 | 9 | 3.8 |
| C. henryi | 26 | 1.2 | 12.1 | 22.4 | 16 | 3.3 |
| Quercus fabri Hance | 27 | 0.5 | 10.9 | 15.3 | 19 | 2.5 |
| Sassafras tzumu (Hemsl.) Hemsl. | 21 | 0.7 | 12.4 | 19.6 | 14 | 2.3 |
| Other 35 species | 294 | 1.6 | 6.6 | 8.3 | 81 | 19.2 |
| Total 45 species | 1831 | 21.6 | 8.3 | 9.6 | 100 | 100.0 |

| Species | Individuals (Stems ha−1) | Mean dbh (cm) | Mean Height (m) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| O | M | U | O | M | U | O | M | U | |
| L. glaber | 191 | 527 | 1084 | 14.3 | 6.4 | 2.3 | 12.1 | 6.9 | 3.3 |
| C. glauca | 86 | 117 | 270 | 19.8 | 7.1 | 2.5 | 13.2 | 6.8 | 3.4 |
| P. massoniana | 110 | 36 | 0 | 16.2 | 12.3 | - | 13.5 | 8.3 | - |
| C. lanceolata | 24 | 124 | 155 | 13.1 | 7.9 | 3 | 12 | 7.2 | 3.3 |
| C. japonica | 2 | 90 | 437 | 12.8 | 5.4 | 2.8 | 12.2 | 5.9 | 3.2 |
| C. axillaris | 52 | 20 | 11 | 20.6 | 14.2 | 3.5 | 13.3 | 8.2 | 3.1 |
| 6 species | 465 | 914 | 1957 | 16.4 | 7 | 2.5 | 12.8 | 6.9 | 3.3 |
| Total | 630 | 1174 | 2993 | 16.0 | 6.9 | 2.5 | 12.9 | 6.9 | 3.2 |
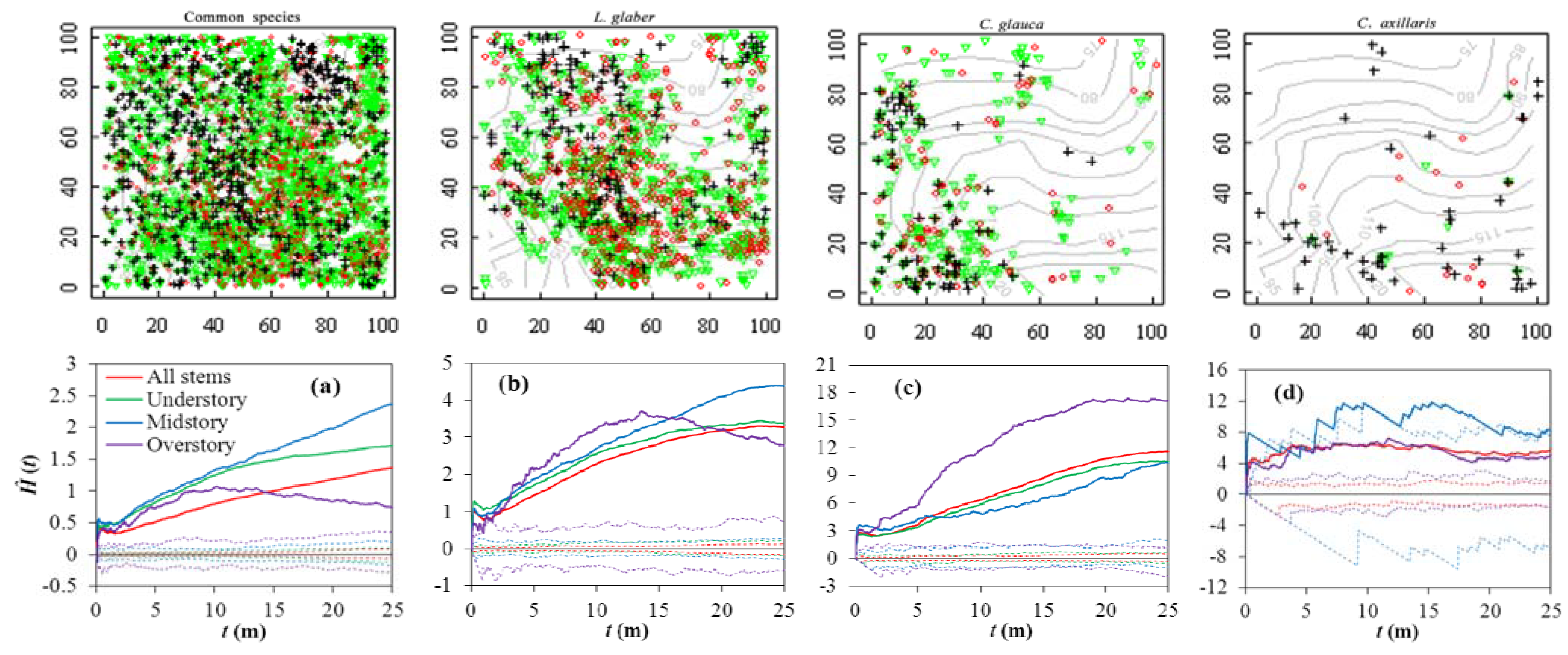
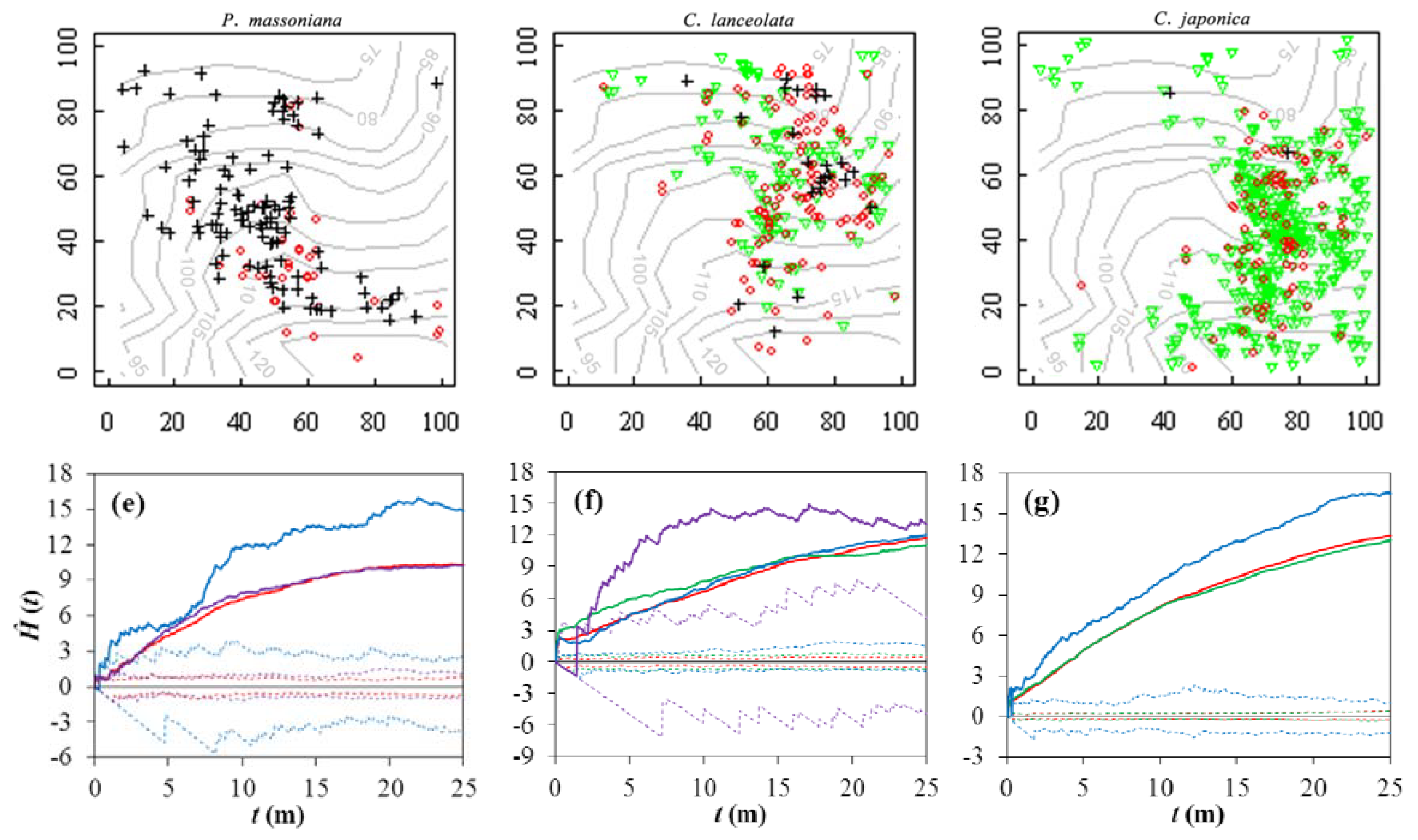
3.2. Spatial Patterns


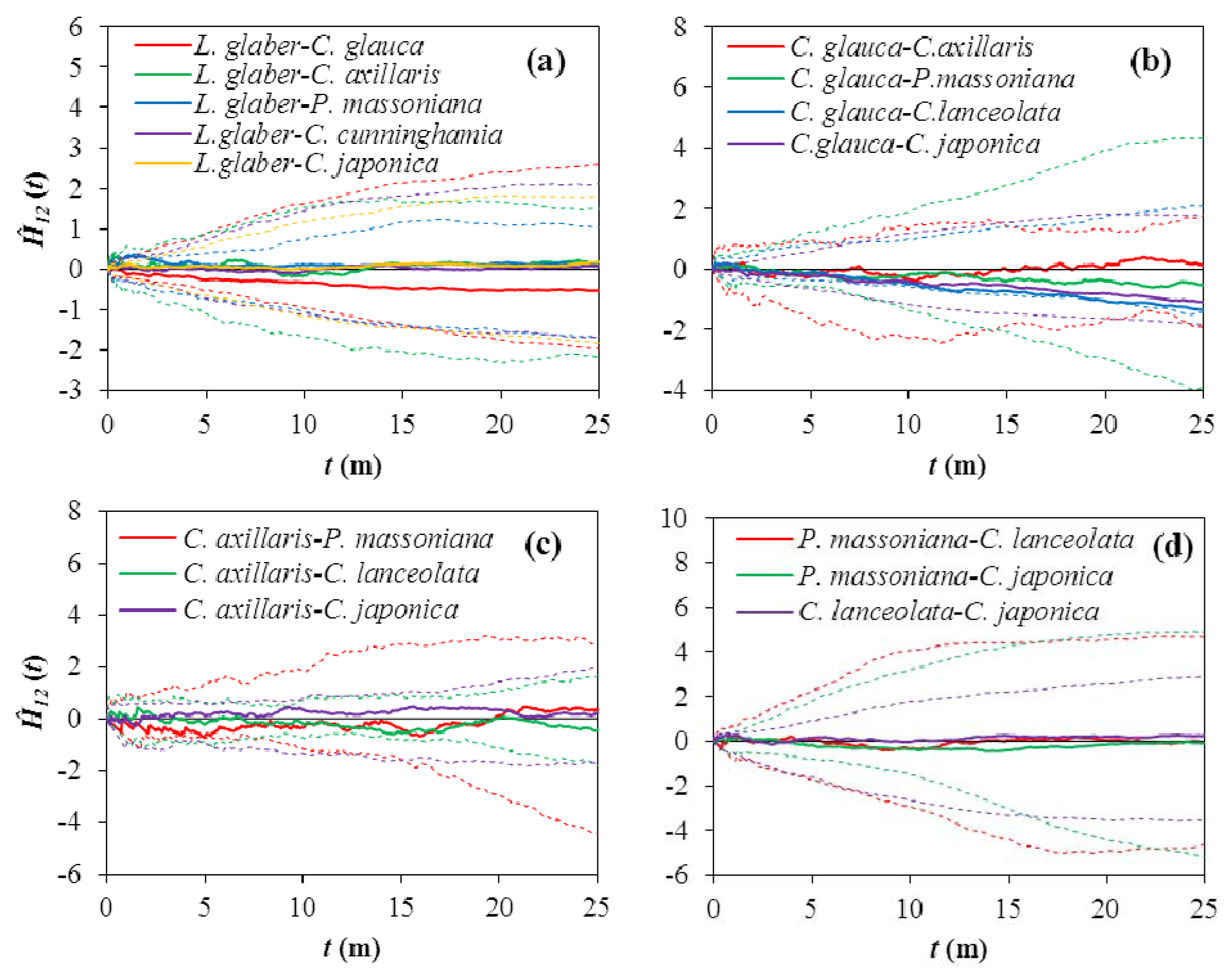
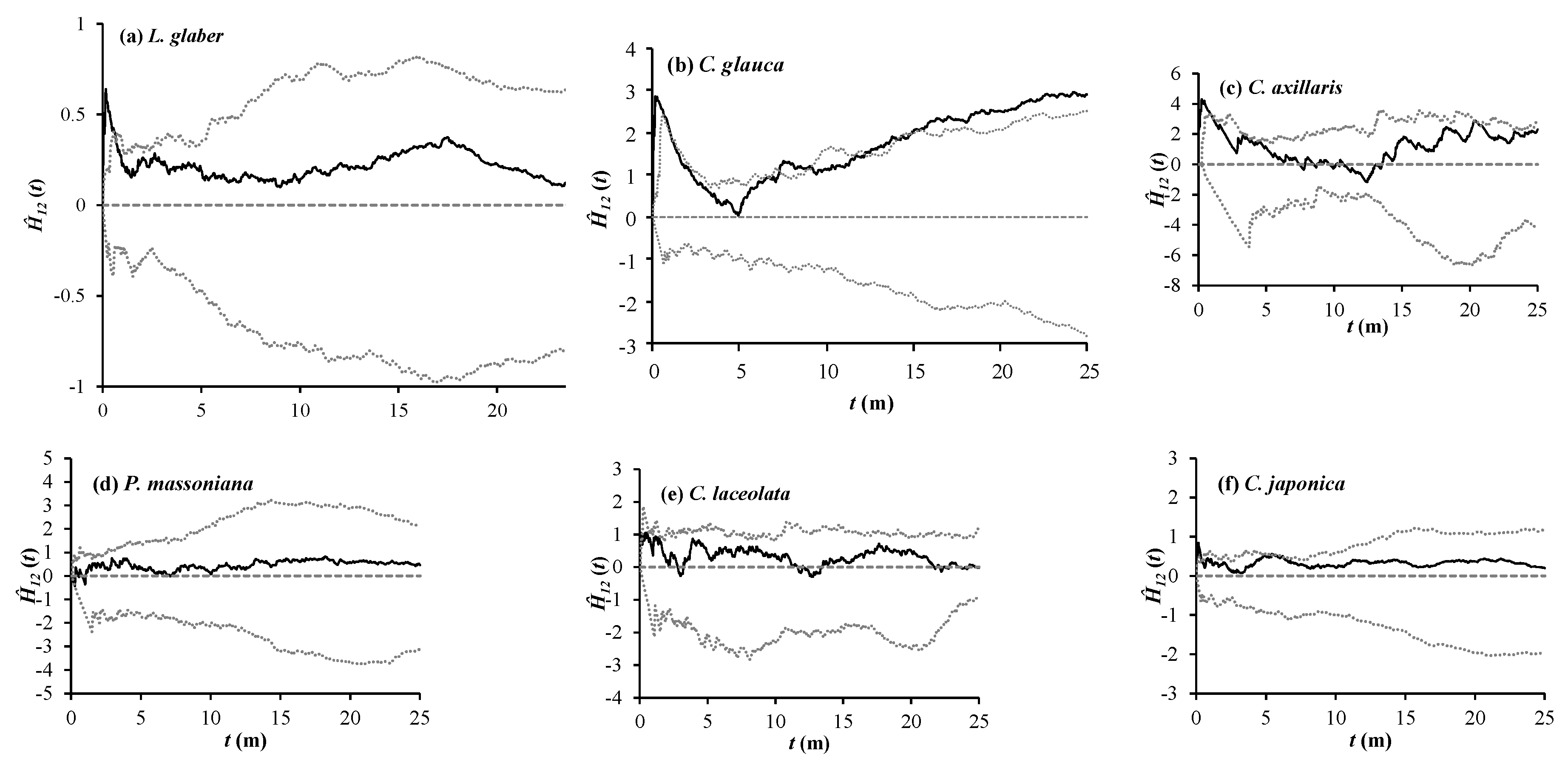
3.3. Spatial Associations


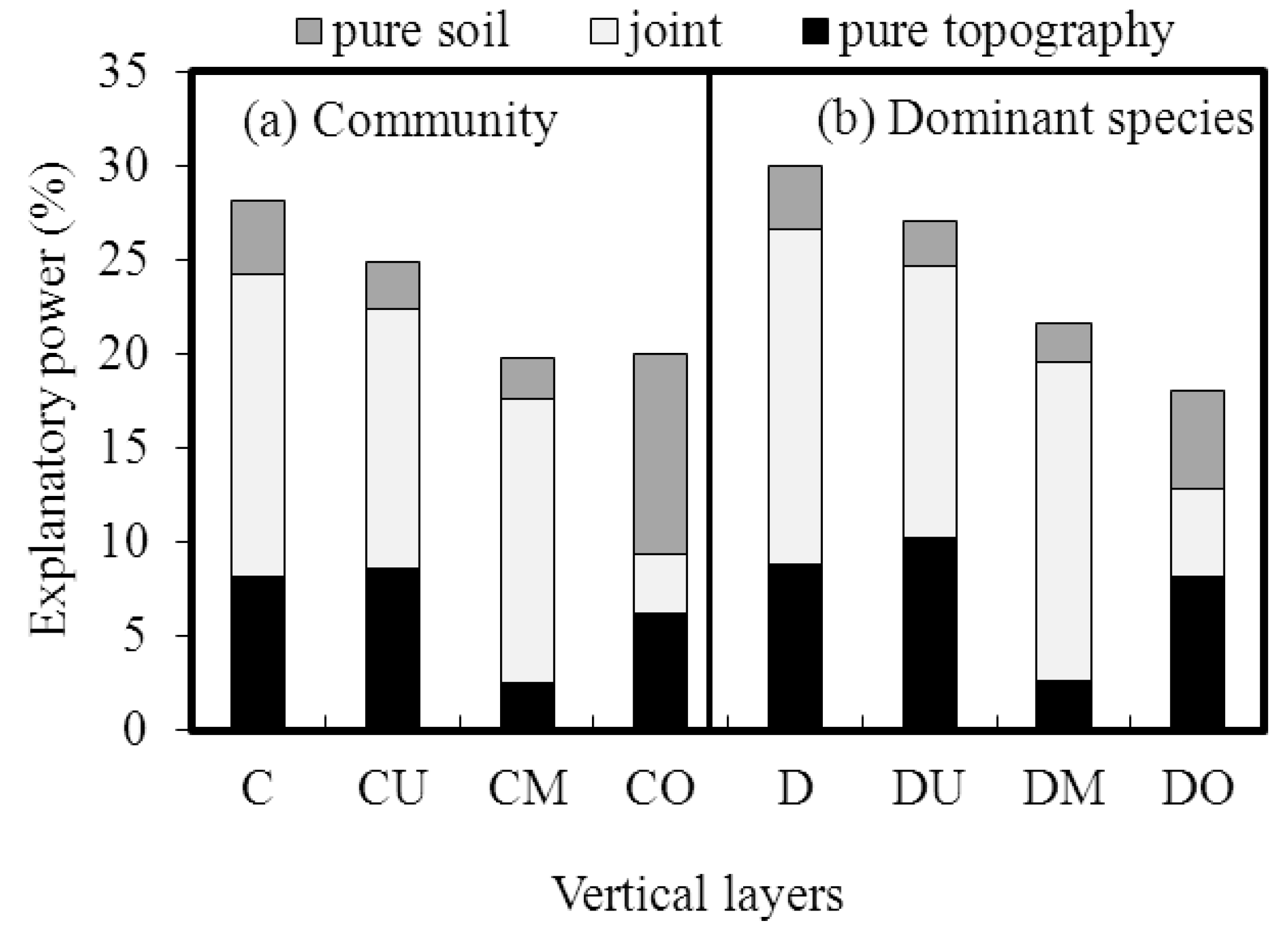
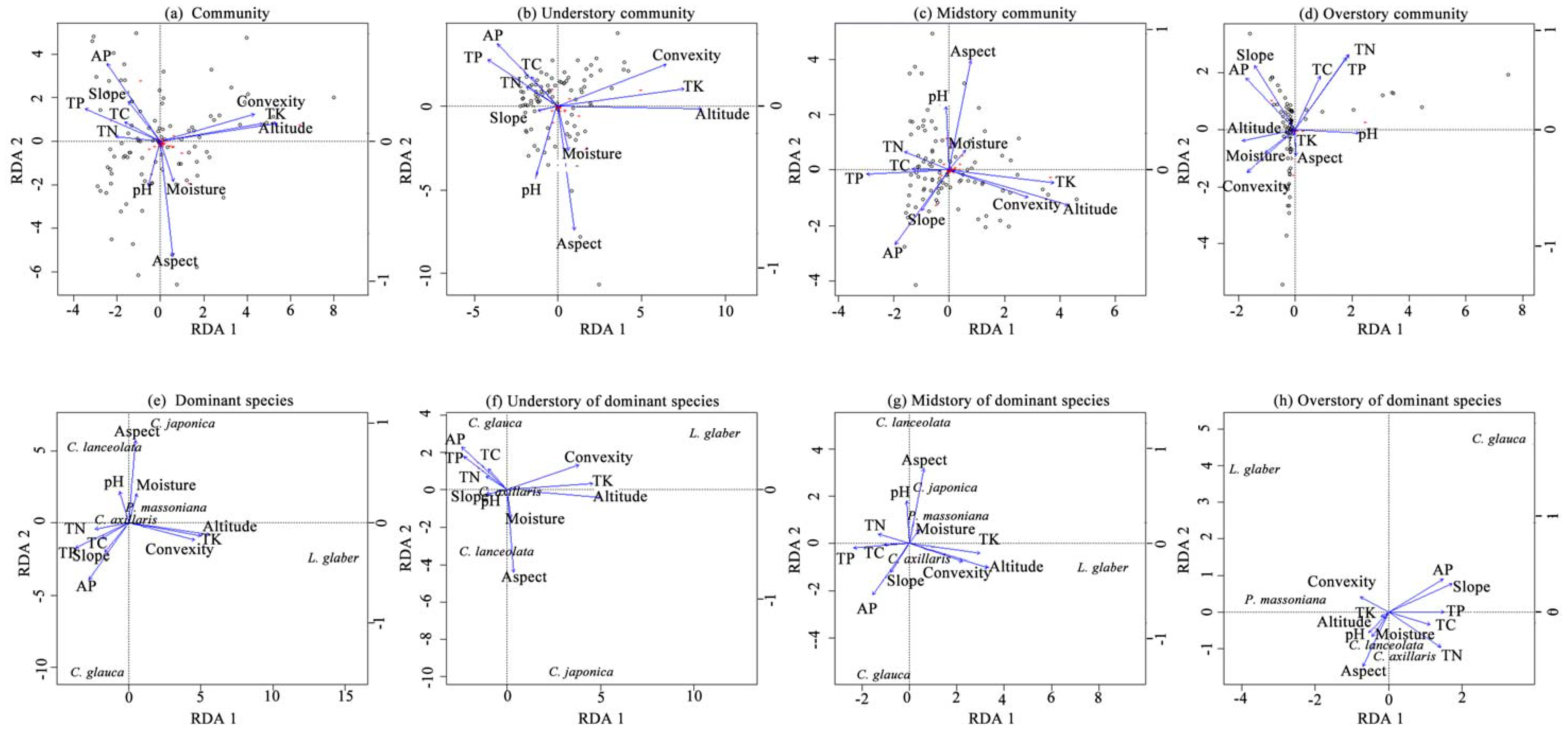
3.4. Contributions of Topographic and Soil Variables

3.5. Main Driving Variables Across Vertical Layers

4. Discussion
4.1. Stand Characteristics and Regeneration
4.2. Implications of Spatial Patterns and Associations for Species Assembly
4.3. Relative Contribution of Topography and Soil
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bruelheide, H.; Böhnke, M.; Both, S.; Fang, T.; Assmann, T.; Baruffol, M.; Bauhus, J.; Buscot, F.; Chen, X.Y.; Ding, B.Y.; et al. Community assembly during secondary forest succession in a Chinese subtropical forest. Ecol. Monogr. 2011, 81, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H.; Slatyer, R.O. Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Veblen, T.T.; Ashton, D.H.; Schlegel, F.M. Tree regeneration strategies in a lowland Nothofagus-dominated forest in south-central Chile. J. Biogeogr. 1979, 6, 329–340. [Google Scholar] [CrossRef]
- Zenner, E.K.; Lahde, E.; Laiho, O. Contrasting the temporal dynamics of stand structure in even- and uneven-sized Picea abies dominated stands. Can. J. For. Res. 2011, 41, 289–299. [Google Scholar] [CrossRef]
- Baraloto, C.; Hardy, O.J.; Paine, C.E.T.; Dexter, K.G.; Cruaud, C.; Dunning, L.T.; Gonzalez, M.A.; Molino, J.F.; Sabatier, D.; Savolainen, V.; et al. Using functional traits and phylogenetic trees to examine the assembly of tropical tree communities. J. Ecol. 2012, 100, 690–701. [Google Scholar] [CrossRef]
- Li, L.; Huang, Z.L.; Ye, W.H.; Cao, H.L.; Wei, S.G.; Wang, Z.G.; Lian, J.Y.; Sun, I.F.; Ma, K.P.; He, F. Spatial distributions of tree species in a subtropical forest of China. Oikos 2009, 118, 495–502. [Google Scholar] [CrossRef]
- Manabe, T.; Nishimura, N.; Miura, M.; Yamamoto, S. Population structure and spatial patterns for trees in a temperate old-growth evergreen broad-leaved forest in Japan. Plant Ecol. 2000, 151, 181–197. [Google Scholar] [CrossRef]
- Shen, G.C.; He, F.L.; Waagepetersen, R.; Sun, I.F.; Hao, Z.Q.; Chen, Z.S.; Yu, M.J. Quantifying effects of habitat heterogeneity and other clustering processes on spatial distributions of tree species. Ecology 2013, 94, 2436–2443. [Google Scholar] [CrossRef] [PubMed]
- Seidler, T.G.; Plotkin, J.B. Seed dispersal and spatial pattern in tropical trees. PLoS Biol. 2006, 4, e344. [Google Scholar] [CrossRef] [PubMed]
- Condit, R.; Ashton, P.S.; Baker, P.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial patterns in the distribution of tropical tree species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.H.; Mi, X.C.; Liu, C.R.; Ma, K.P. Spatial patterns and associations in a Quercus- Betula forest in northern China. J. Veg. Sci. 2004, 15, 407–414. [Google Scholar]
- Chen, L.Z.; Chen, W.L. Chinese Degraded Ecological System Research; Science Press: Beijing, China, 1995. [Google Scholar]
- Yamada, T.; Zuidema, P.A.; Itoh, A.; Yamakura, T.; Ohkubo, T.; Kanzaki, M.; Tan, S.; Ashton, P.S. Strong habitat preference of a tropical rain forest tree does not imply large differences in population dynamics across habitats. J. Ecol. 2007, 95, 332–342. [Google Scholar] [CrossRef]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- John, R.; Dalling, J.W.; Harms, K.E.; Yavitt, J.B.; Stallard, R.F.; Mirabello, M.; Hubbell, S.P.; Valencia, R.; Navarrete, H.; Vallejo, M.; et al. Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. USA 2007, 104, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P.; Mi, X.; Ren, H.B.; Ma, K.; Yu, M.; Sun, I.F.; He, F. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hu, G.; Zhu, J.; Ni, J. Aggregated spatial distributions of species in a subtropical karst forest, southwestern China. J. Plant Ecol. 2013, 6, 131–140. [Google Scholar] [CrossRef]
- Chang, L.W.; Zelený, D.; Li, C.F.; Chiu, S.T.; Hsieh, C.F. Better environmental data may reverse conclusions about niche-and dispersal-based processes in community assembly. Ecology 2013, 94, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Y.; Zhao, Y.Z.; Zhao, X.H.; Klaus, V.G. Species-Habitat Associations in a Northern Temperate Forest in China. Silva Fenn. 2012, 46, 501–519. [Google Scholar]
- Comita, L.S.; Condit, R.; Hubbell, S.P. Developmental changes in habitat associations of tropical trees. J. Ecol. 2007, 95, 482–492. [Google Scholar] [CrossRef]
- Hao, Z.; Zhang, J.; Song, B.; Ye, J.; Li, B. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. For. Ecol. Manag. 2007, 252, 1–11. [Google Scholar] [CrossRef]
- Qi, C.J. Hunan Vegetation; Hunan Science and Technology Press: Changsha, China, 1990; pp. 51–341. [Google Scholar]
- Xiang, W.H.; Liu, S.H.; L, X.D.; Frank, S.C.; Tian, D.L.; Wang, G.J.; Deng, X.W. Secondary forest floristic composition, structure, and spatial pattern in subtropical China. J. For. Res. 2013, 18, 111–120. [Google Scholar] [CrossRef]
- Yasmi, Y.; Broadhead, J.; Enters, T.; Genge, C. Forestry Policies, Legislation and Institutions in Asia and the Pacific: Trends and Emerging Needs for 2020, Asia-Pacific Forestry Sector Outlook Study II; Food and Agriculture Organization of the United Nations: Bangkok, Thailand, 2010. [Google Scholar]
- Diaci, J. Nature-Based Forestry in Central Europe: Alternatives to Industrial forestry and Strict Preservation; Biotechnical Faculty, University of Ljubljana: Ljubljana, Slovenia, 2006. [Google Scholar]
- Lai, J.S. Species Habitat Associations and Species Coexistence on Evergreen Broad-Leaved Forest in Gutianshan, Zhejiang. Ph.D. Thesis, Chinese Academy of Science, Beijing, China, June 2008. [Google Scholar]
- Xie, Y.B.; Ma, Z.P.; Yang, Q.S.; Fang, X.F.; Zhang, Z.G.; Yan, E.R.; Wang, X.H. Coexistence mechanisms of evergreen and deciduous trees based on topographic factors in Tiantong region, Zhejiang Province, eastern China. Biodivers. Sci. 2012, 20, 159–167. [Google Scholar]
- Zhao, L.J.; Xiang, W.H.; Li, J.X.; Deng, X.W.; Liu, C. Composition, structure and phytogeographic characteristics in a Lithocarpus glaber-Cyclobalanopsis glauca forest community in mid-subtropical China. Sci. Silvae Sin. 2013, 49, 10–17. [Google Scholar]
- Tonon, G.; Panzacchi, P.; Grassi, G.; Gianfranco, M.; Cntoni, L.; Bagnaresi, U. Spatial dynamics of late successional species under Pinus nigra stands in the northern Apennines (Italy). Ann. For. Sci. 2005, 62, 669–679. [Google Scholar] [CrossRef]
- Lan, G.Y.; Hu, Y.H.; Cao, M.; Zhu, H. Topography related spatial distribution of dominant tree species in a tropical seasonal rain forest in China. For. Ecol. Manag. 2011, 262, 1507–1513. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Galindo-Jaimes, L.; González-Espinosa, M.; Quintana-Ascencio, P.; García-Barrios, L. Tree composition and structure in disturbed stands with varying dominance by Pinus spp. in the highlands of Chiapas, Mexico. Plant Ecol. 2002, 162, 259–272. [Google Scholar] [CrossRef]
- Ripley, B.D. Modelling spatial patterns. J. R. Stat. Soc. B 1977, 39, 172–212. [Google Scholar]
- Haase, P. Spatial pattern analysis in ecology based on Ripley’s K-function: Introduction and methods of edge correction. J. Veg. Sci. 1995, 6, 575–582. [Google Scholar] [CrossRef]
- Ripley, B.D. Test of randomness for spatial point patterns. J. R. Stat. Soc. B 1979, 41, 368–374. [Google Scholar]
- Besag, J.; Diggle, P.J. Simple Monte Carlo test for spatial pattern. J. Appl. Stat. 1977, 26, 327–333. [Google Scholar] [CrossRef]
- Salas, C.; LeMay, V.; Núñez, P.; Pacheco, P.; Espinosa, A. Spatial patterns in an old-growth Nothofagus obliqua forest in south-central Chile. For. Ecol. Manag. 2006, 231, 38–46. [Google Scholar] [CrossRef]
- Baddeley, A.; Turner, R. Spatstat: An R package for analyzing spatial point patterns. J. Stat. Softw. 2005, 12, 1–42. [Google Scholar]
- Core Development Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009. Available online: http://www.R-project.org (accessed on 22 April 2006).
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package, R package version 1.14-12; Available online: http://vegan.r-forge.r-project.org/ (accessed on 3 June 2008).
- Takahashi, K.; Homma, K.; Vetrova, V.P.; Florenzev, S.; Hara, T. Stand structure and regeneration in a Kamchatka mixed boreal forest. J. Veg. Sci. 2001, 12, 627–634. [Google Scholar] [CrossRef]
- Plotkin, J.B.; Potts, M.D.; Leslie, N.; Manokaran, N.; LaFrankie, J.; Ashton, P.S. Species- area curves, spatial aggregation, and habitat specialization in tropical forests. J. Theor. Biol. 2000, 207, 81–99. [Google Scholar] [CrossRef] [PubMed]
- He, F.L.; Legendre, P.; LaFrankie, J.V. Distribution patterns of tree species in a Malaysian tropical rain forest. J. Veg. Sci. 1997, 8, 105–114. [Google Scholar] [CrossRef]
- Liu, Q.J. Structure and dynamics of the subalpine coniferous forest on Changbai mountain, China. Plant Ecol. 1997, 132, 97–105. [Google Scholar] [CrossRef]
- Hubbell, S.P. Neutral theory and the evolution of ecological equivalence. Ecology 2006, 87, 1387–1398. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Gazol, A.; Wang, X.; Lin, F.; Ye, J.; Bai, X.; Li, B.; Hao, Z. Scale specific determinants of tree diversity in an old growth temperate forest in China. Basic Appl. Ecol. 2011, 12, 488–495. [Google Scholar] [CrossRef]
- Lin, G.J.; Stralberg, D.; Gong, G.Q.; Huang, Z.L.; Ye, W.H.; Wu, L.F. Separating the effects of environment and space on tree species distribution: From population to community. PLoS One 2013, 8, e56171. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.M.; Tuomisto, H.; Borcard, D.; Legendre, P.; Clark, D.B.; Olivas, P.C. Explaining variation in tropical plant community composition: Influence of environmental and spatial data quality. Oecologia 2008, 155, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Punchi-Manage, R.; Getzin, S.; Wiegand, T.; Kanagaraj, R.; Savitri Gunatilleke, C.V.; Nimal Gunatilleke, I.A.U.; Wiegand, K.; Huth, A.; Zuidema, P. Effects of topography on structuring local species assemblages in a Sri Lankan mixed dipterocarp forest. J. Ecol. 2013, 101, 149–160. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Xiang, W.; Li, J.; Lei, P.; Deng, X.; Fang, X.; Peng, C. Effects of Topographic and Soil Factors on Woody Species Assembly in a Chinese Subtropical Evergreen Broadleaved Forest. Forests 2015, 6, 650-669. https://doi.org/10.3390/f6030650
Zhao L, Xiang W, Li J, Lei P, Deng X, Fang X, Peng C. Effects of Topographic and Soil Factors on Woody Species Assembly in a Chinese Subtropical Evergreen Broadleaved Forest. Forests. 2015; 6(3):650-669. https://doi.org/10.3390/f6030650
Chicago/Turabian StyleZhao, Lijuan, Wenhua Xiang, Jiaxiang Li, Pifeng Lei, Xiangwen Deng, Xi Fang, and Changhui Peng. 2015. "Effects of Topographic and Soil Factors on Woody Species Assembly in a Chinese Subtropical Evergreen Broadleaved Forest" Forests 6, no. 3: 650-669. https://doi.org/10.3390/f6030650
APA StyleZhao, L., Xiang, W., Li, J., Lei, P., Deng, X., Fang, X., & Peng, C. (2015). Effects of Topographic and Soil Factors on Woody Species Assembly in a Chinese Subtropical Evergreen Broadleaved Forest. Forests, 6(3), 650-669. https://doi.org/10.3390/f6030650