Population Spatial Dynamics of Larix potaninii in Alpine Treeline Ecotone in the Eastern Margin of the Tibetan Plateau, China


Abstract
:1. Introduction
2. Materials and Methods
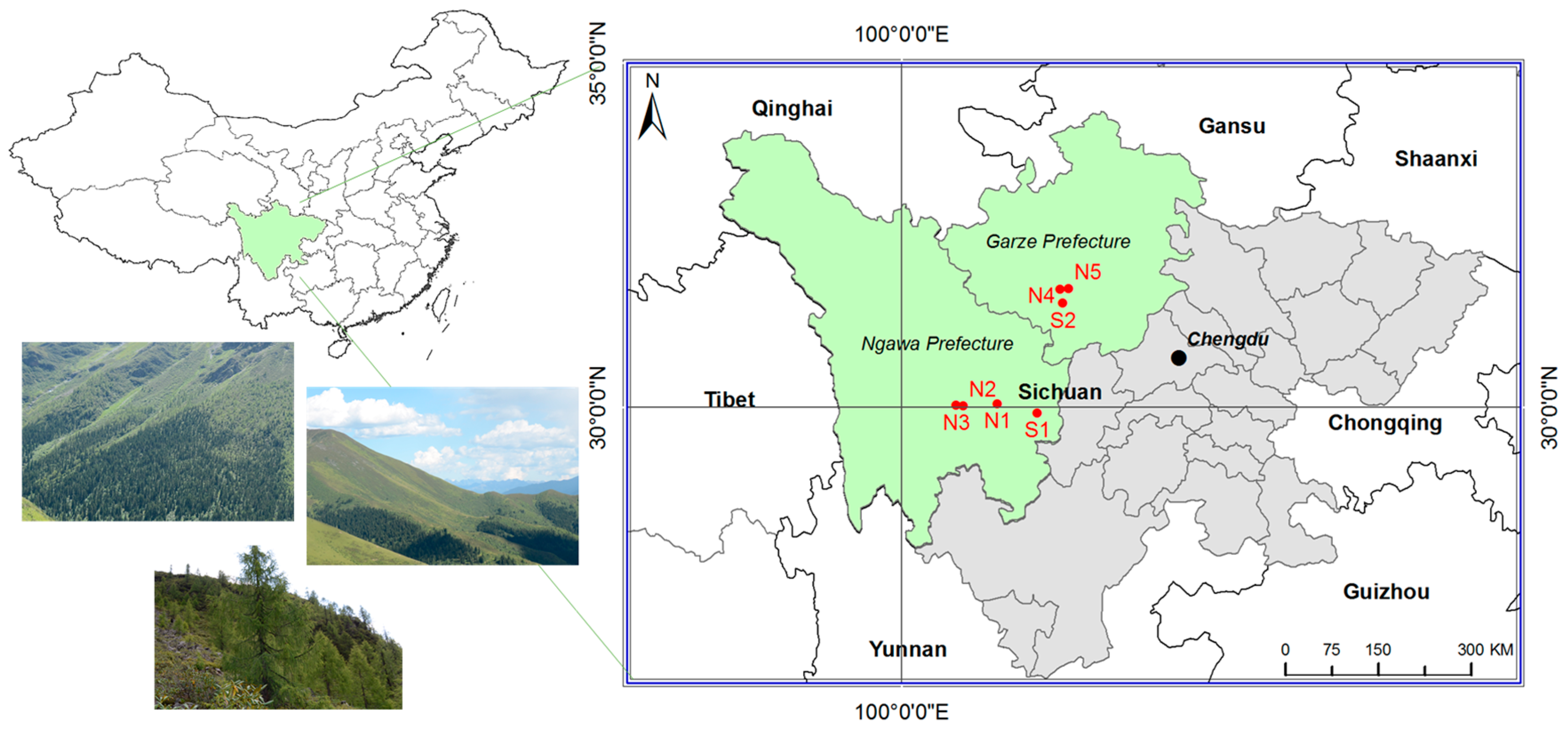
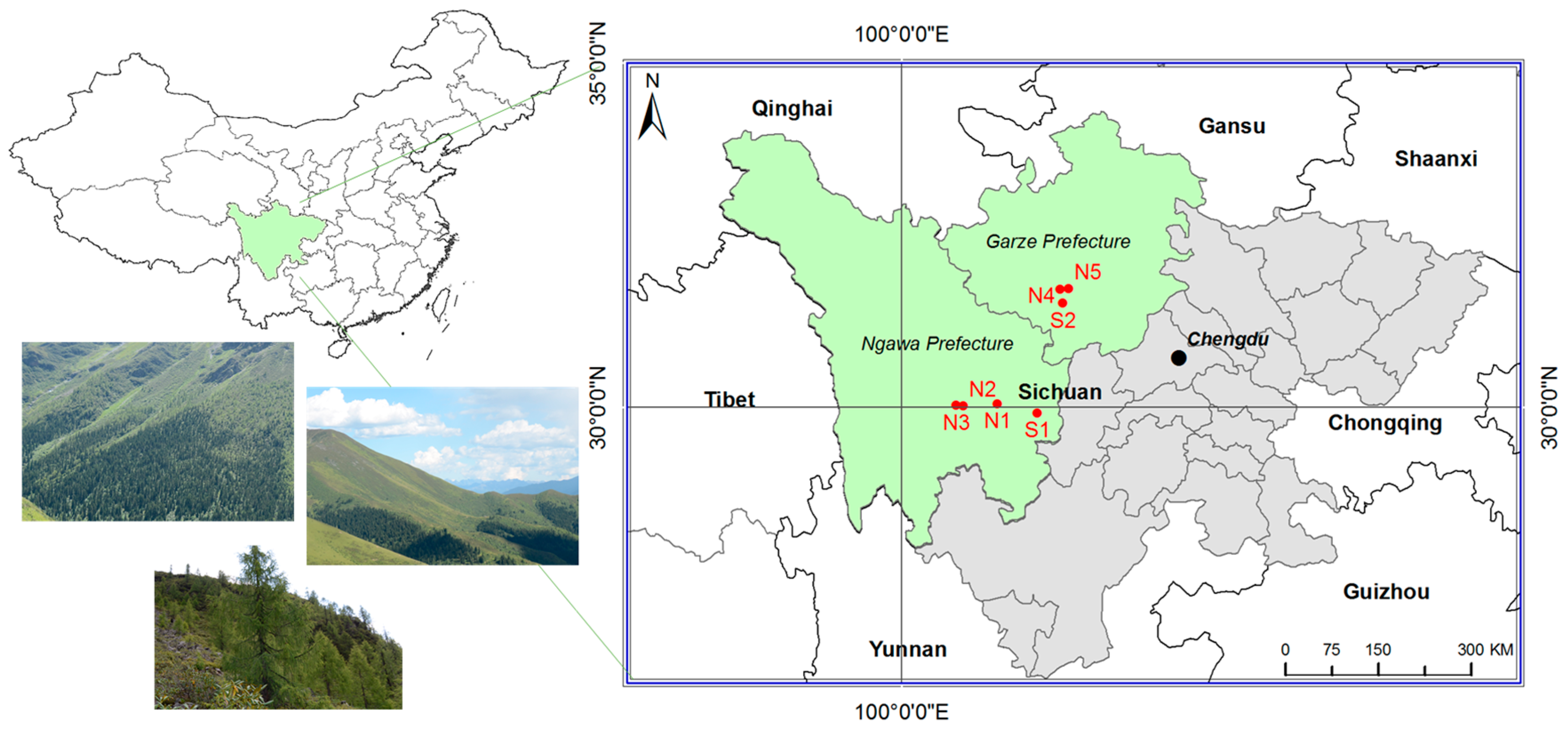
2.1. Study Area and Site Selection
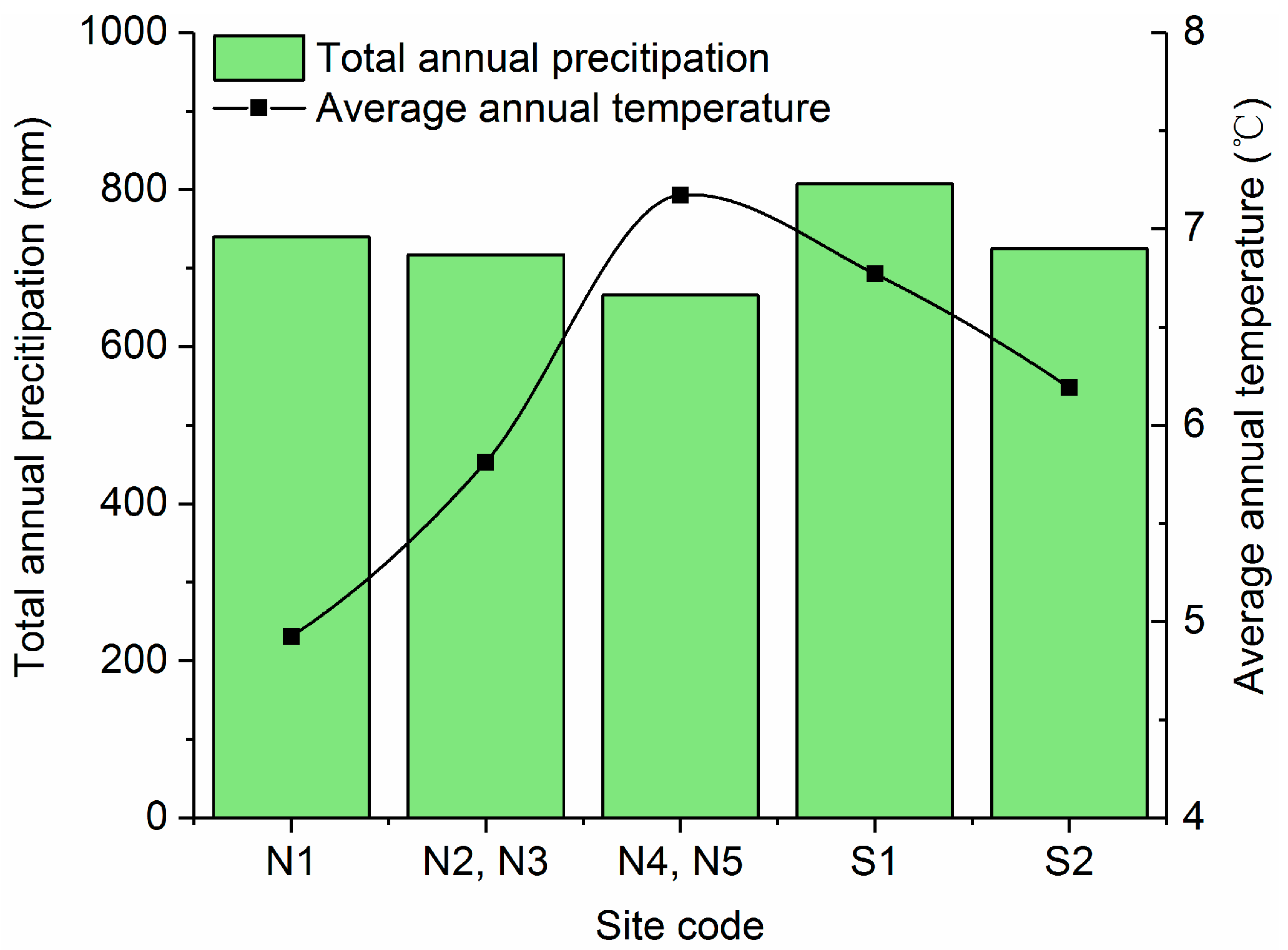
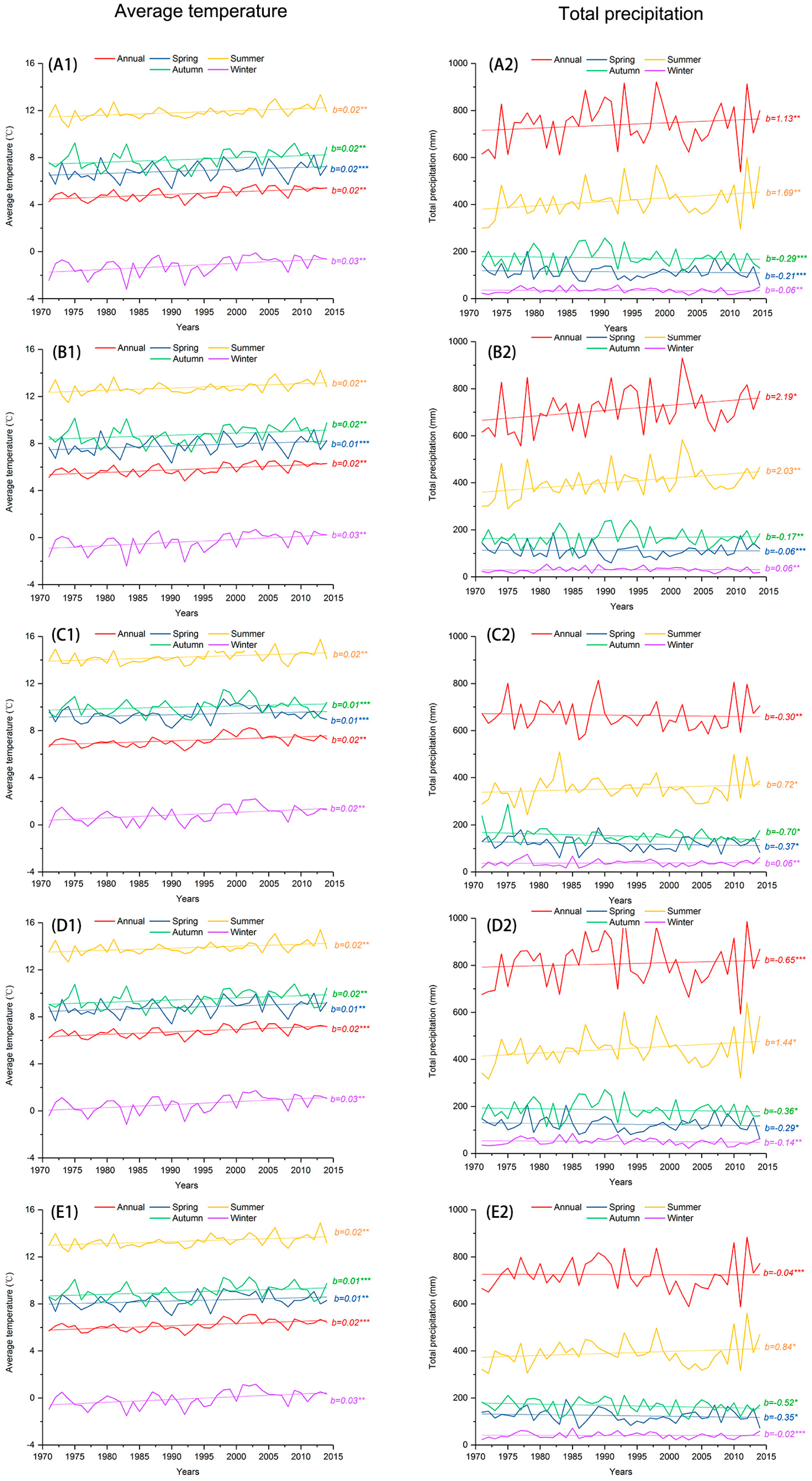
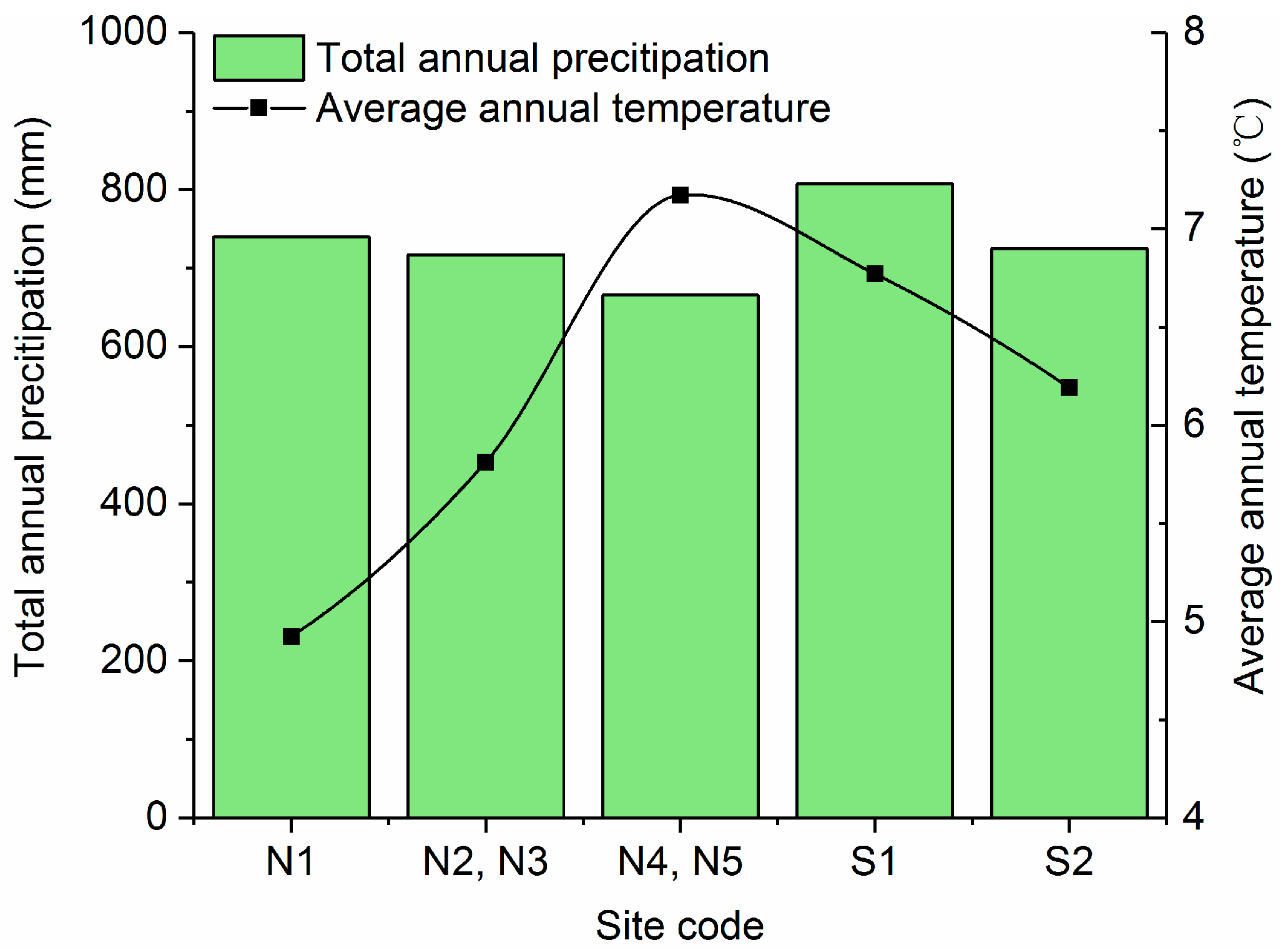
2.2. Climate Data
2.3. Field Investigation and Tree-Age Estimation
2.4. Treeline Dynamics
2.5. Recruitment and Density Dynamics
2.6. Spatial Distribution Pattern
3. Results
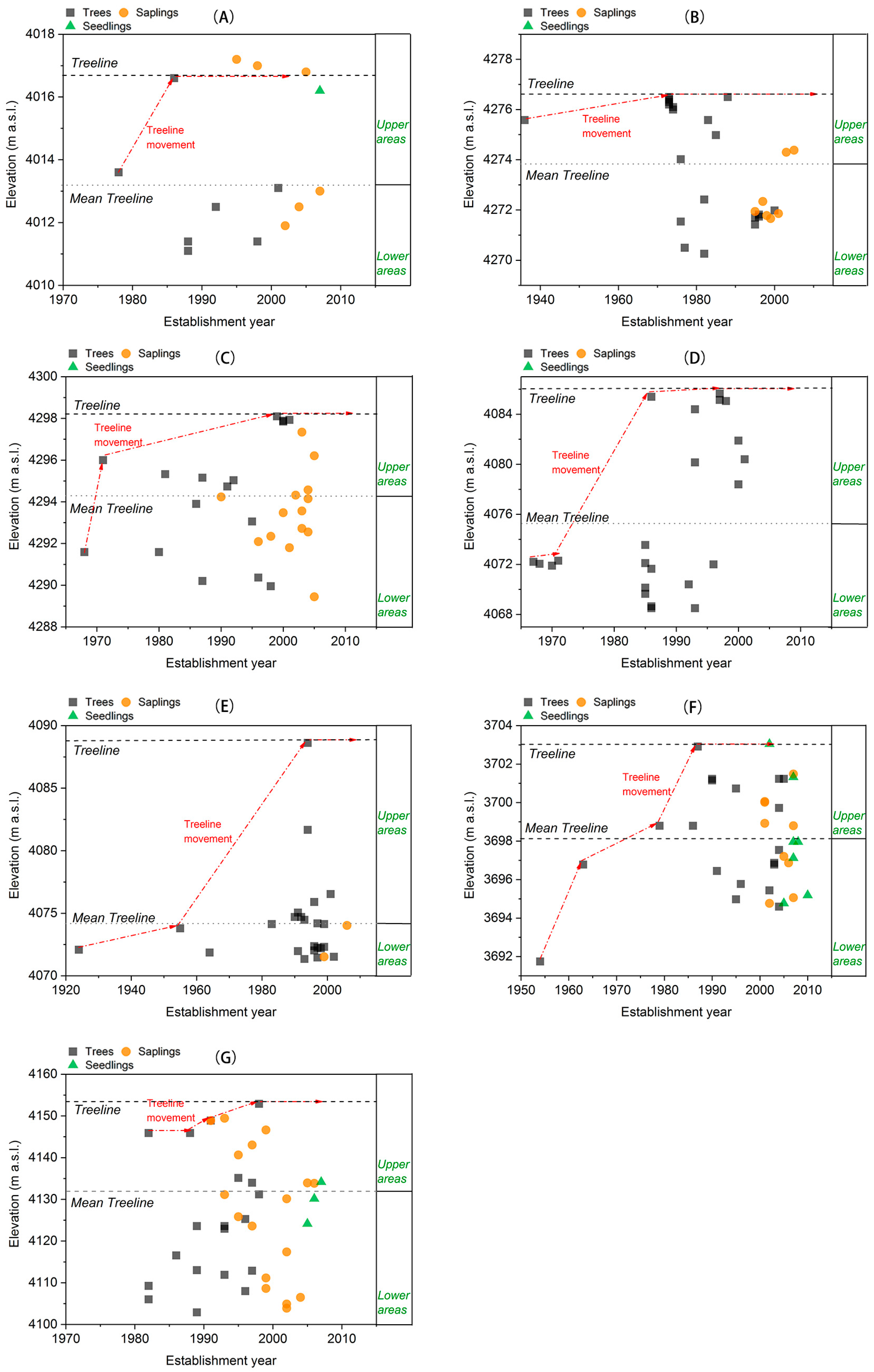
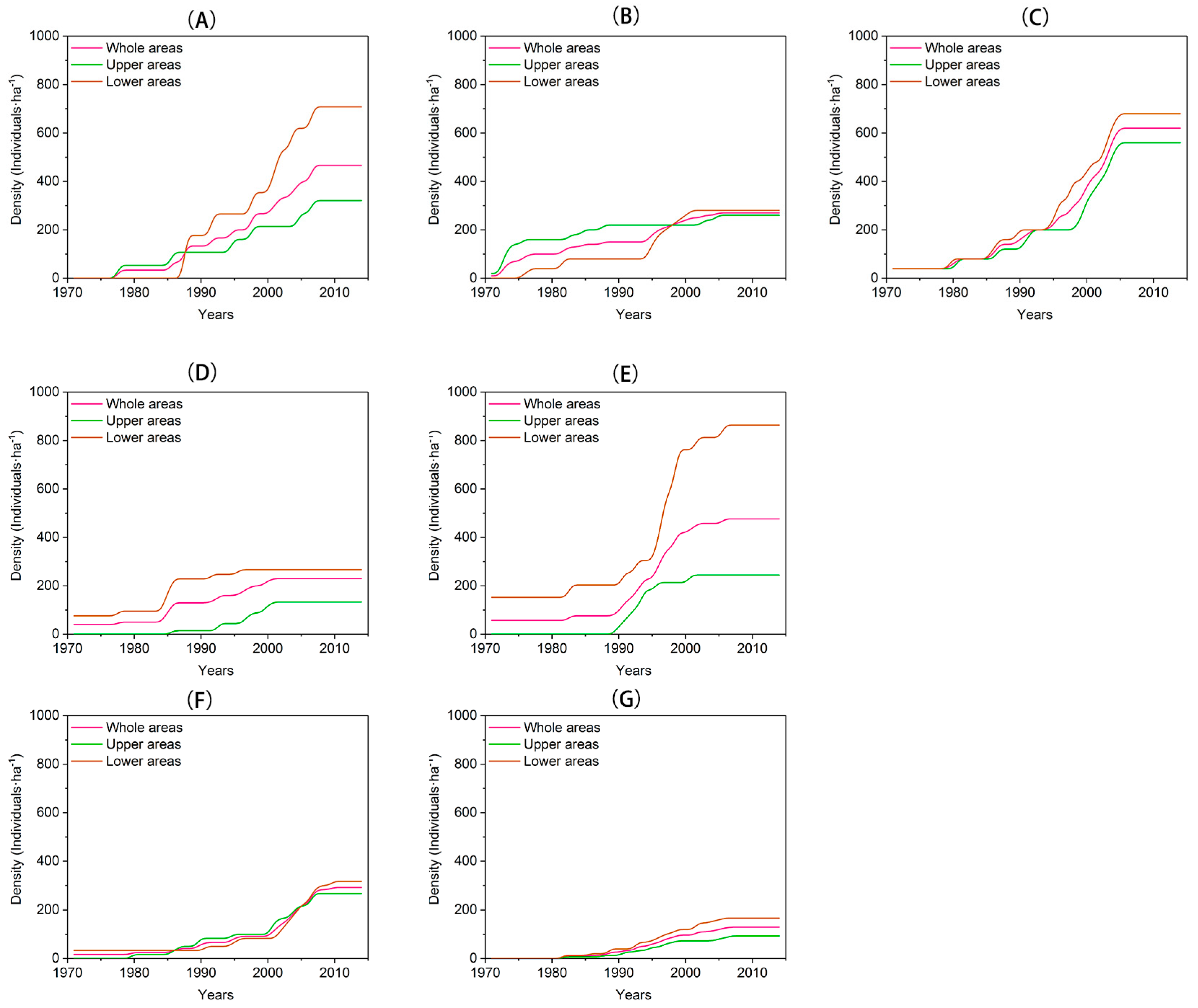
3.1. Treeline Dynamics
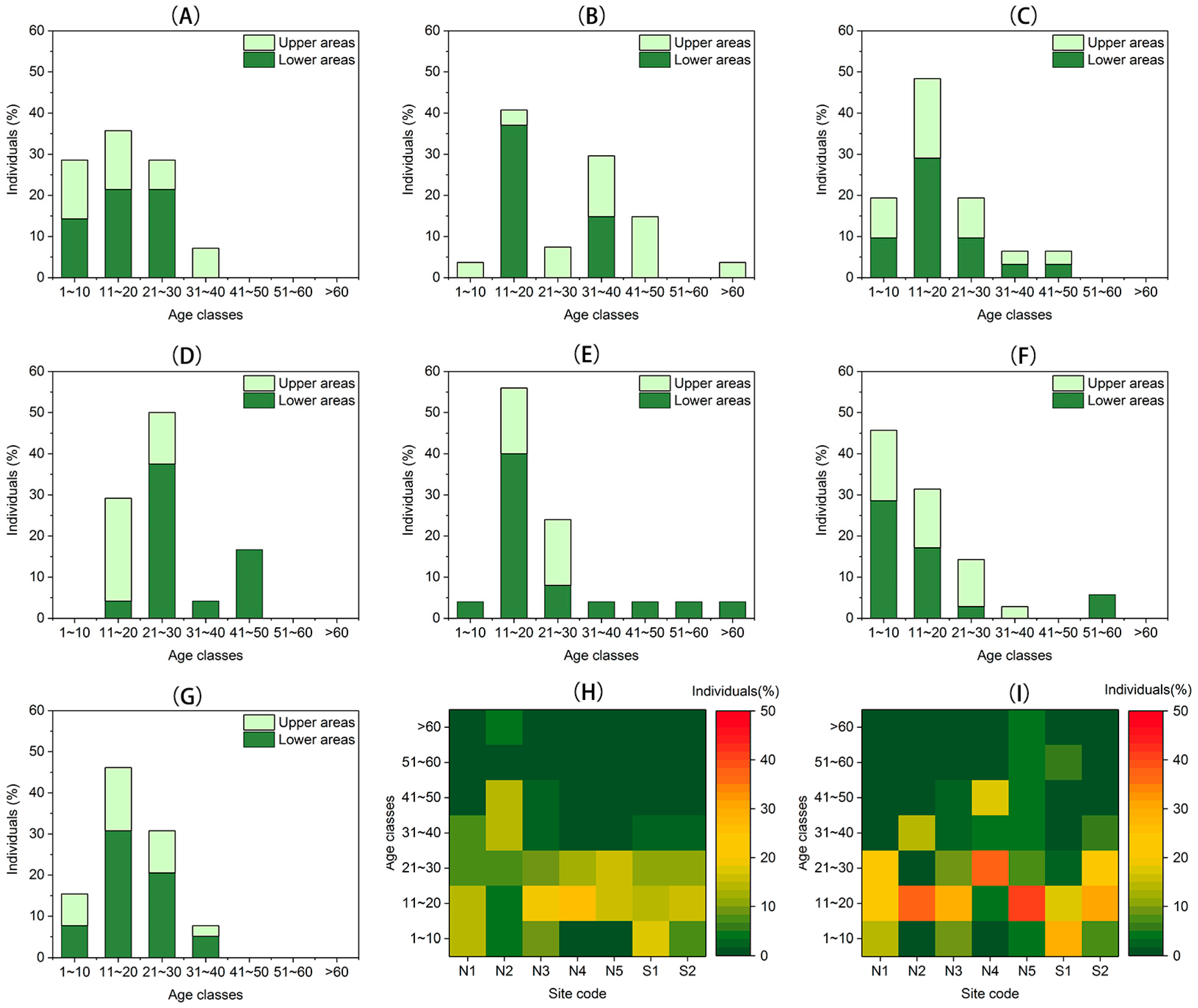
3.2. Recruitment and Density Dynamics
3.3. Spatial Distribution Patterns
4. Discussion
4.1. Treeline Dynamics and Recruitment
4.2. Slope Exposure Effects and Climate Change
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station Code | Longitude (°E) | Latitude (°N) | Elevation (m a.s.l.) | Duration Period | Nearby Sampling Sites |
|---|---|---|---|---|---|
| Yajiang | 101.0 | 30.0 | 2600.9 | 1971–2014 | N1 |
| Litang | 100.2 | 30.0 | 3948.9 | 1971–2014 | N2, N3 |
| Maerkang | 102.1 | 31.5 | 2664.4 | 1971–014 | N4, N5 |
| Kangding | 101.6 | 30.0 | 2615.7 | 1971–1982 | S1 |
| Xiaojin | 102.2 | 31.0 | 2369.0 | 1971–2014 | S2 |
| Station Code | Nearby Sampling Sites | Temperature | Precipitation |
|---|---|---|---|
| Yajiang | N1 | 0.997 ** | 0.992 ** |
| Litang | N2, N3 | 0.999 ** | 0.978 ** |
| Maerkang | N4, N5 | 0.996 ** | 0.976 ** |
| Kangding | S1 | 0.996 ** | 0.959 ** |
| Xiaojin | S2 | 0.933 ** | 0.958 ** |
References
- Stocker, T. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014; ISBN 110705799X. [Google Scholar]
- Chhetri, P.K.; Cairns, D.M. Contemporary and historic population structure of Abies spectabilis at treeline in Barun valley, eastern Nepal Himalaya. J. Mt. Sci. 2015, 12, 558–570. [Google Scholar] [CrossRef]
- Hofgaard, A.; Dalen, L.; Hytteborn, H. Tree recruitment above the treeline and potential for climate-driven treeline change. J. Veg. Sci. 2009, 20, 1133–1144. [Google Scholar] [CrossRef]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Kullman, L. Recent tree-limit history of Picea abies in the southern Swedish Scandes. Can. J. For. Res. 1986, 16, 761–771. [Google Scholar] [CrossRef]
- Major, J.; Kullman, L. Change and stability in the altitude of the birch tree-limit in the southern Swedish Scandes 1915–75. Arct. Alp. Res. 2012, 11, 477. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gutiérrez, E. Pace and pattern of recent treeline dynamics: Response of ecotones to climatic variability in the Spanish Pyrenees. Clim. Chang. 2004, 63, 181–200. [Google Scholar] [CrossRef]
- Paulsen, J.; Körner, C. GIS-analysis of tree-line elevation in the Swiss Alps suggests no exposure effect. J. Veg. Sci. 2001, 12, 817–824. [Google Scholar] [CrossRef]
- Lloyd, A.H.; Rupp, T.S.; Fastie, C.L.; Starfield, A.M. Patterns and dynamics of treeline advance on the Seward Peninsula, Alaska. J. Geophys. Res. Atmos. 2002, 107. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Qi, Z.; Liu, H.; Wu, X.; Hao, Q. Climate-driven speedup of alpine treeline forest growth in the Tianshan Mountains, Northwestern China. Glob. Chang. Biol. 2015, 21, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.B.; Moseley, R.K. Advancing treeline and retreating glaciers: Implications for conservation in Yunnan, P.R. China. Arct. Antarct. Alp. Res. 2007, 39, 200–209. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Liang, E.; Wang, Y.; Eckstein, D.; Luo, T. Little change in the fir tree-line position on the southeastern Tibetan Plateau after 200 years of warming. New Phytol. 2011, 190, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Holtmeier, F.-K.; Broll, G. Treeline advance–driving processes and adverse factors. Lands. Online 2007, 1, 1–33. [Google Scholar] [CrossRef]
- Kullman, L.; Öberg, L. Post-Little Ice Age tree line rise and climate warming in the Swedish Scandes: A landscape ecological perspective. J. Ecol. 2009, 97, 415–429. [Google Scholar] [CrossRef]
- Panigrahy, S.; Anitha, D.; Kimothi, M.; Singh, S. Timberline change detection using topographic map and satellite imagery. Tropical Ecol. 2010, 51, 87–91. [Google Scholar]
- Shrestha, K.B.; Hofgaard, A.; Vandvik, V. Recent treeline dynamics are similar between dry and mesic areas of Nepal, central Himalaya. J. Plant Ecol. 2015, 8, 347–358. [Google Scholar] [CrossRef]
- Holtmeier, F.-K. Mountain Timberlines: Ecology, Patchiness, and Dynamics; Springer Science & Business Media: Berlin, Germany, 2009; Volume 36. [Google Scholar]
- Hagedorn, F.; Shiyatov, S.G.; Mazepa, V.S.; Devi, N.M.; Grigor’ev, A.A.; Bartysh, A.A.; Fomin, V.V.; Kapralov, D.S.; Terent’ev, M.; Bugman, H. Treeline advances along the Urals mountain range–driven by improved winter conditions? Glob. Chang. Biol. 2014, 20, 3530–3543. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, K.B.; Hofgaard, A.; Vandvik, V. Tree-growth response to climatic variability in two climatically contrasting treeline ecotone areas, central Himalaya, Nepal. Can. J. For. Res 2015, 45, 1643–1653. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Glob. Ecol. Biogeogr. 2005, 14, 395–410. [Google Scholar] [CrossRef]
- Juntunen, V.N.S. Natural regeneration of Scots pine and Norway spruce close to the timberline in northern Finland. Silv. Fenn. 2006, 40, 443–458. [Google Scholar] [CrossRef]
- Wiegand, T.; Gunatilleke, S.; Gunatilleke, N. Species associations in a heterogeneous Sri lankan dipterocarp forest. Am. Nat. 2007, 170, E77. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Camarero, J.J.; Luo, T.; Liang, E. Spatial patterns of Smith fir alpine treelines on the south-eastern Tibetan Plateau support that contingent local conditions drive recent treeline patterns. Plant Ecol. Divers. 2012, 5, 311–321. [Google Scholar] [CrossRef]
- Klasner, F.L.; Fagre, D.B. A half century of change in alpine treeline patterns at Glacier National Park, Montana, USA. Arct. Antarct. Alp. Res. 2002, 49–56. [Google Scholar] [CrossRef]
- MacDonald, G.M.; Szeicz, J.M.; Claricoates, J.; Dale, K.A. Response of the central Canadian treeline to recent climatic changes. Ann. Assoc. Am. Geogr. 1998, 88, 183–208. [Google Scholar] [CrossRef]
- Burga, C.A.; Perret, R. Monitoring of Eastern and Southern Swiss Alpine Timberline Ecotones; Springer Netherlands: Dordrecht, The Netherlands, 2001; pp. 179–194. [Google Scholar]
- Shi, P.; Körner, C.; Hoch, G. A test of the growth-limitation theory for alpine tree line formation in evergreen and deciduous taxa of the eastern Himalayas. Funct. Ecol. 2008, 22, 213–220. [Google Scholar] [CrossRef]
- Fang, K.; Gou, X.; Chen, F.; Peng, J.; D’Arrigo, R.; Wright, W.; Li, M.-H. Response of regional tree-line forests to climate change: Evidence from the northeastern Tibetan Plateau. Trees 2009, 23, 1321. [Google Scholar] [CrossRef]
- Liu, W.; Wang, L.; Chen, D.; Tu, K.; Ruan, C.; Hu, Z. Large-scale circulation classification and its links to observed precipitation in the eastern and central Tibetan Plateau. Clim. Dyn. 2015, 46, 3481–3497. [Google Scholar] [CrossRef]
- Elliott, G.P.; Cowell, C.M. Slope aspect mediates fine-scale tree establishment patterns at upper treeline during wet and dry periods of the 20th century. Arct. Antarct. Alp. Res. 2015, 47, 681–692. [Google Scholar] [CrossRef]
- Elliott, G.P.; Kipfmueller, K.F. Multiscale influences of climate on upper treeline dynamics in the southern rocky mountains, USA: Evidence of intraregional variability and bioclimatic thresholds in response to twentieth-century warming. Ann. Assoc. Am. Geogr. 2011, 101, 1181–1203. [Google Scholar] [CrossRef]
- Danby, R.K.; Hik, D.S. Responses of white spruce (Picea glauca) to experimental warming at a subarctic alpine treeline. Glob. Chang. Biol. 2007, 13, 437–451. [Google Scholar] [CrossRef]
- Qiu, D.-Y.; Yang, J.; Feng, L.; Ni, D.-W.; Huang, C.-X.; Li, Z.-H. Complete plastid genome of Larix potaninii var. macrocarpa, an endangered conifer endemic to China. Conserv. Genet. Resour. 2017. [Google Scholar] [CrossRef]
- University of East Anglia. Climatic Research Unit. Available online: http://www.cru.uea.ac.uk/data/ (accessed on 20 July 2017).
- Zhao, T.; Fu, C. Comparison of Products from ERA-40,NCEP-2,and CRU with Station Data for Summer Precipitation over China. Adv. Atmos. Sci. 2006, 23, 593–604. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, Q.B.; Ma, K. Treeline dynamics in relation to climatic variability in the central Tianshan Mountains, northwestern China. Glob. Ecol. Biogeogr. 2006, 15, 406–415. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree rings and climate. Ecology 1977, 58, 95–99. [Google Scholar] [CrossRef]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences; Springer Science & Business Media: Berlin, Germany, 2013; ISBN 9401578796. [Google Scholar]
- Stokes, M.A. An Introduction to Tree-Ring Dating; University of Arizona Press: Tuscon, AZ, USA, 1996; ISBN 0816516804. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983. [Google Scholar] [CrossRef]
- Buckley, H.L.; Case, B.S.; Vallejos, R.; Camarero, J.J.; Gutiérrez, E.; Liang, E.; Wang, Y.; Ellison, A.M. Detecting ecological patterns along environmental gradients: Alpine treeline ecotones. Chance 2016, 29, 10–15. [Google Scholar] [CrossRef]
- Pielou, E. A single mechanism to account for regular, random and aggregated populations. J. Ecol. 1960, 575–584. [Google Scholar] [CrossRef]
- Lashkari, M.; Shahbazvar, N. Spatial distribution pattern of Diaphorina citri Kuwayama (Hem.: Liviidae) on Lime (Citrus aurantifolia) and Orange (Citrus sinensis) in citrus orchards of Iran. Entomol. Appl. Sci. Lett. 2016, 3, 81–87. [Google Scholar] [CrossRef]
- Shiyatov, S.; Terent’ev, M.; Fomin, V.; Zimmermann, N. Altitudinal and horizontal shifts of the upper boundaries of open and closed forests in the Polar Urals in the 20th century. Russ. J. Ecol. 2007, 38, 223. [Google Scholar] [CrossRef]
- Elliott, G.P.; Kipfmueller, K.F. Multi-scale influences of slope aspect and spatial pattern on ecotonal dynamics at upper treeline in the Southern Rocky Mountains, U.S.A. Ann. Assoc. Am. Geogr. 2010, 42, 45–56. [Google Scholar] [CrossRef]
- Danby, R.K.; Hik, D.S. Variability, contingency and rapid change in recent subarctic alpine tree line dynamics. J. Ecol. 2007, 95, 352–363. [Google Scholar] [CrossRef]
- You, Q.; Min, J.; Lin, H.; Pepin, N.; Sillanpää, M.; Kang, S. Observed climatology and trend in relative humidity in the central and eastern Tibetan Plateau. J. Geophys. Res. Atmos. 2015, 120, 3610–3621. [Google Scholar] [CrossRef] [Green Version]

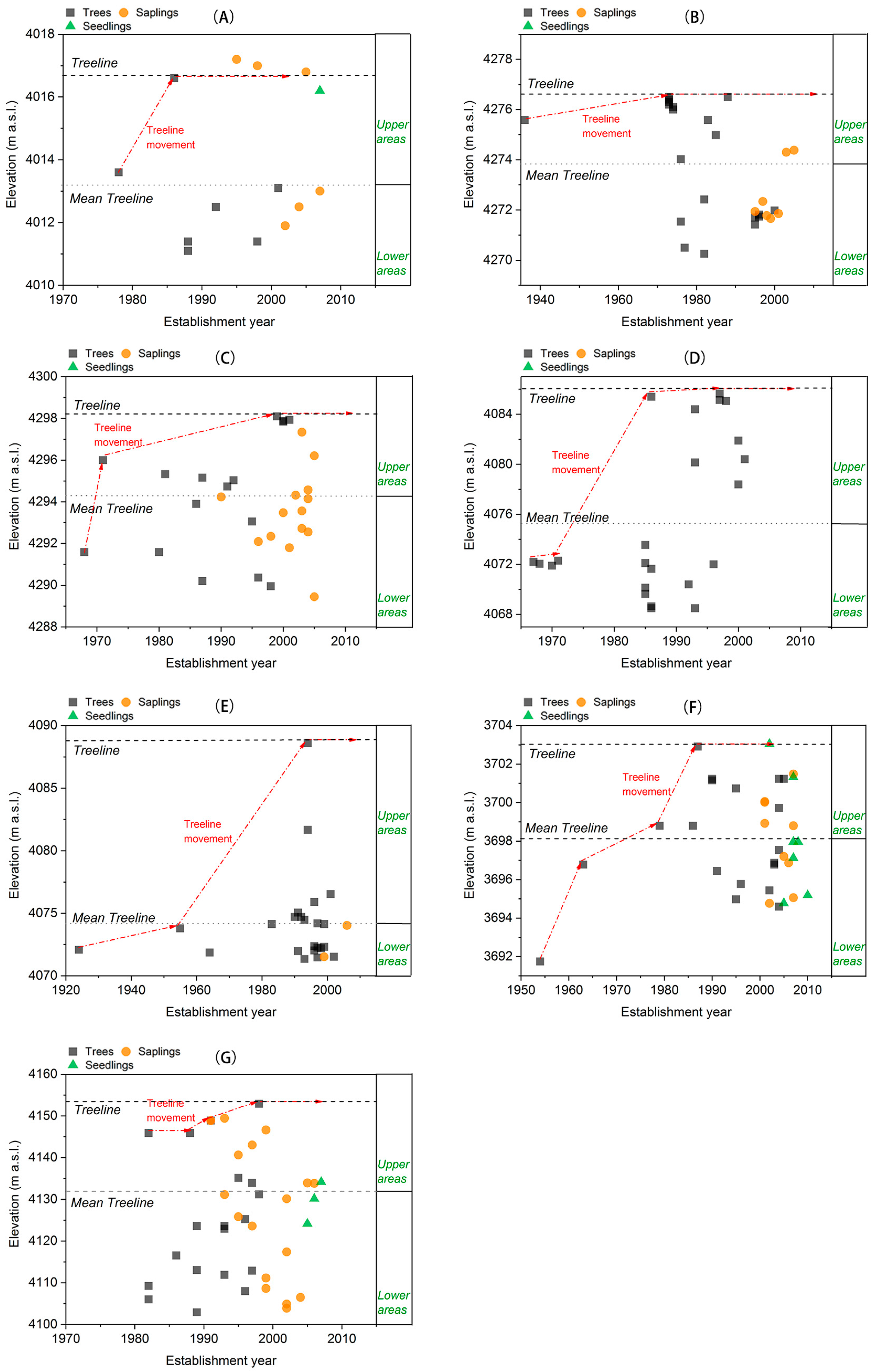

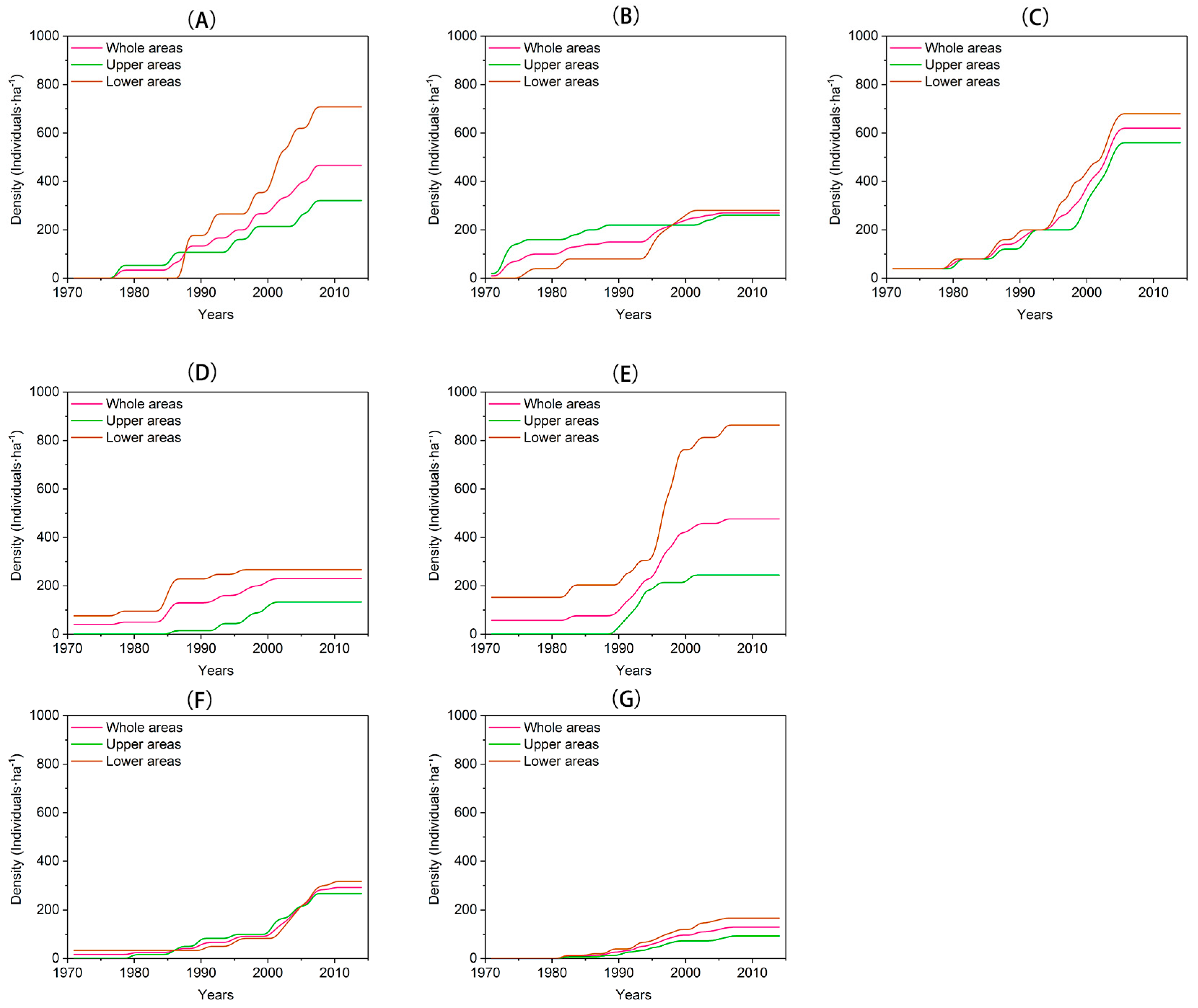
| Site Code | Elevation (m a.s.l.) | Slope Inclination | Slope Aspect | Horizontal Length (m) | Vertical Length (m) | Numbers. of Larix Individuals |
|---|---|---|---|---|---|---|
| N1 | 4015 | 30° | NW 30° | 20 | 15 | 14 |
| N2 | 4274 | 20° | NE 50° | 40 | 25 | 27 |
| N3 | 4293 | 25° | NW 50° | 25 | 20 | 31 |
| N4 | 4076 | 30° | NW 53° | 30 | 40 | 24 |
| N5 | 4076 | 35° | N | 15 | 35 | 25 |
| S1 | 3698 | 25° | SW 75° | 40 | 30 | 35 |
| S2 | 4214 | 25° | S | 30 | 100 | 39 |
| N1 | N2 | N3 | N4 | N5 | S1 | S2 | ||
|---|---|---|---|---|---|---|---|---|
| Monthly mean temperature | October * | ○ | ○ | ○ | ○ | ○ | ○○ | |
| November * | ● | ● | ● | ● | ● | ● | ● | |
| December * | ● | ● | ● | ● | ● | ● | ● | |
| January | ● | ● | ● | ● | ● | ● | ● | |
| February | ● a | ● b | ● b | ● b | ● a | ● b | ● a | |
| March | ● | ● | ● | ● | ● | ● | ● | |
| April | ○ | ○ | ○ | ○ | ○ | ○ | ○ | |
| May | ● | ● | ● | ● | ● | |||
| June | ● | ● | ● | ● | ● | ● | ● | |
| July | ● | ○ | ● | ○ | ● | ● | ● | |
| August | ● | ○ | ● | ○ | ○ | ○ | ○ | |
| September | ● | ● | ● | ● | ● | ● | ● | |
| Total Monthly precipitation | October * | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| November * | ● | ○ | ○ | ○ | ● | ○ | ● | |
| December * | ● | ○ | ○ | ○ | ○ | ○ | ○ | |
| January | ● | ○ | ● | ● | ● | ● | ● | |
| February | ● | ○ | ● | ● | ● | ○ | ● | |
| March | ● | ● | ● | ● | ● | ● | ○ | |
| April | ● b | ● | ● | ● | ● | ● | ● | |
| May | ○ | ○ | ○ | ○ | ○ | ○ | ○ | |
| June | ● | ● | ● | ● | ● | ● b | ● | |
| July | ● | ● | ○ | ○ | ○ | ○ | ○ | |
| August | ● | ○ | ○ | ○ | ○ | ○ | ○ | |
| September | ● | ○ | ○ | ○ | ○ | ○ | ○ | |
| Site | Item | N | DI | m* | PI | |||
|---|---|---|---|---|---|---|---|---|
| N1 | T | 300 | 0.05 | 0.07 | 1.53 | 0.58 | 0.04 | 0.09 |
| U | 187 | 0.03 | 0.11 | 4.14 | 3.16 | 0.78 | 0.01 | |
| L | 113 | 0.07 | 0.09 | 1.30 | 0.37 | 0.04 | 0.24 | |
| N2 | T | 1000 | 0.03 | 0.10 | 3.77 | 2.80 | 0.11 | 0.01 |
| U | 500 | 0.03 | 0.04 | 1.44 | 0.46 | 0.04 | 0.06 | |
| L | 500 | 0.03 | 0.07 | 2.55 | 1.58 | 0.12 | 0.02 | |
| N3 | T | 500 | 0.06 | 0.06 | 1.00 | 0.07 | 0.00 | 13.70 |
| U | 250 | 0.06 | 0.09 | 1.52 | 0.58 | 0.04 | 0.11 | |
| L | 250 | 0.07 | 0.07 | 1.05 | 0.12 | 0.00 | 1.26 | |
| N4 | T | 1200 | 0.02 | 0.02 | 1.23 | 0.25 | 0.01 | 0.09 |
| U | 675 | 0.01 | 0.02 | 1.66 | 0.67 | 0.08 | 0.02 | |
| L | 525 | 0.03 | 0.04 | 1.37 | 0.40 | 0.03 | 0.08 | |
| N5 | T | 525 | 0.05 | 0.06 | 1.35 | 0.40 | 0.01 | 0.13 |
| U | 328 | 0.02 | 0.04 | 1.49 | 0.52 | 0.07 | 0.05 | |
| L | 197 | 0.09 | 0.11 | 1.29 | 0.38 | 0.02 | 0.30 | |
| S1 | T | 1200 | 0.03 | 0.03 | 1.09 | 0.12 | 0.00 | 0.34 |
| U | 600 | 0.03 | 0.03 | 1.19 | 0.21 | 0.01 | 0.14 | |
| L | 600 | 0.03 | 0.03 | 1.08 | 0.11 | 0.00 | 0.42 | |
| S2 | T | 3000 | 0.01 | 0.03 | 2.42 | 1.43 | 0.04 | 0.01 |
| U | 1500 | 0.01 | 0.01 | 1.42 | 0.43 | 0.03 | 0.02 | |
| L | 1500 | 0.02 | 0.02 | 1.22 | 0.24 | 0.01 | 0.08 |
| Site | Age Classes | N | DI | m* | PI | |||
|---|---|---|---|---|---|---|---|---|
| N1 | 1~20 | 300 | 0.03 | 0.06 | 1.87 | 0.90 | 0.11 | 0.03 |
| 21~40 | 300 | 0.02 | 0.41 | 24.67 | 23.68 | 5.92 | 0.00 | |
| N2 | 1~20 | 1000 | 0.01 | 0.02 | 1.66 | 0.67 | 0.06 | 0.02 |
| 21~40 | 1000 | 0.01 | 0.01 | 1.39 | 0.40 | 0.04 | 0.03 | |
| 41~60 | 1000 | 0.00 | 0.01 | 2.75 | 1.75 | 0.58 | 0.00 | |
| >60 | 1000 | 0.00 | 0.00 | 1.00 | 0.00 | / | / | |
| N3 | 1~20 | 500 | 0.04 | 0.04 | 1.06 | 0.10 | 0.00 | 0.76 |
| 21~40 | 500 | 0.02 | 0.01 | 0.86 | −0.12 | −0.02 | −0.12 | |
| 41~60 | 500 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | −2.00 | |
| N4 | 1~20 | 1200 | 0.01 | 0.01 | 0.99 | 0.00 | 0.00 | −1.17 |
| 21~40 | 1200 | 0.01 | 0.01 | 1.14 | 0.15 | 0.01 | 0.08 | |
| 41~60 | 1200 | 0.00 | 0.01 | 4.24 | 3.25 | 1.08 | 0.00 | |
| N5 | 1~20 | 525 | 0.03 | 0.99 | 34.69 | 33.72 | 2.41 | 0.00 |
| 21~40 | 525 | 0.01 | 0.01 | 0.99 | 0.00 | 0.00 | −1.16 | |
| 41~60 | 525 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | −2.00 | |
| >60 | 525 | 0.00 | 0.00 | 1.00 | 0.00 | / | / | |
| S1 | 1~20 | 1200 | 0.02 | 0.02 | 1.05 | 0.07 | 0.00 | 0.43 |
| 21~40 | 1200 | 0.01 | 0.01 | 1.33 | 0.33 | 0.07 | 0.02 | |
| 41~60 | 1200 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | −2.00 | |
| S2 | 1~20 | 3000 | 0.01 | 0.01 | 0.99 | 0.00 | 0.00 | −1.04 |
| 21~40 | 3000 | 0.01 | 0.00 | 1.00 | 0.00 | 0.00 | −1.07 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, J.; Qin, J.; Sun, H. Population Spatial Dynamics of Larix potaninii in Alpine Treeline Ecotone in the Eastern Margin of the Tibetan Plateau, China. Forests 2017, 8, 356. https://doi.org/10.3390/f8100356
Cui J, Qin J, Sun H. Population Spatial Dynamics of Larix potaninii in Alpine Treeline Ecotone in the Eastern Margin of the Tibetan Plateau, China. Forests. 2017; 8(10):356. https://doi.org/10.3390/f8100356
Chicago/Turabian StyleCui, Jia’nan, Jihong Qin, and Hui Sun. 2017. "Population Spatial Dynamics of Larix potaninii in Alpine Treeline Ecotone in the Eastern Margin of the Tibetan Plateau, China" Forests 8, no. 10: 356. https://doi.org/10.3390/f8100356
APA StyleCui, J., Qin, J., & Sun, H. (2017). Population Spatial Dynamics of Larix potaninii in Alpine Treeline Ecotone in the Eastern Margin of the Tibetan Plateau, China. Forests, 8(10), 356. https://doi.org/10.3390/f8100356