Soil Degradation and the Decline of Available Nitrogen and Phosphorus in Soils of the Main Forest Types in the Qinling Mountains of China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Sampling Work
2.3. Data Analysis
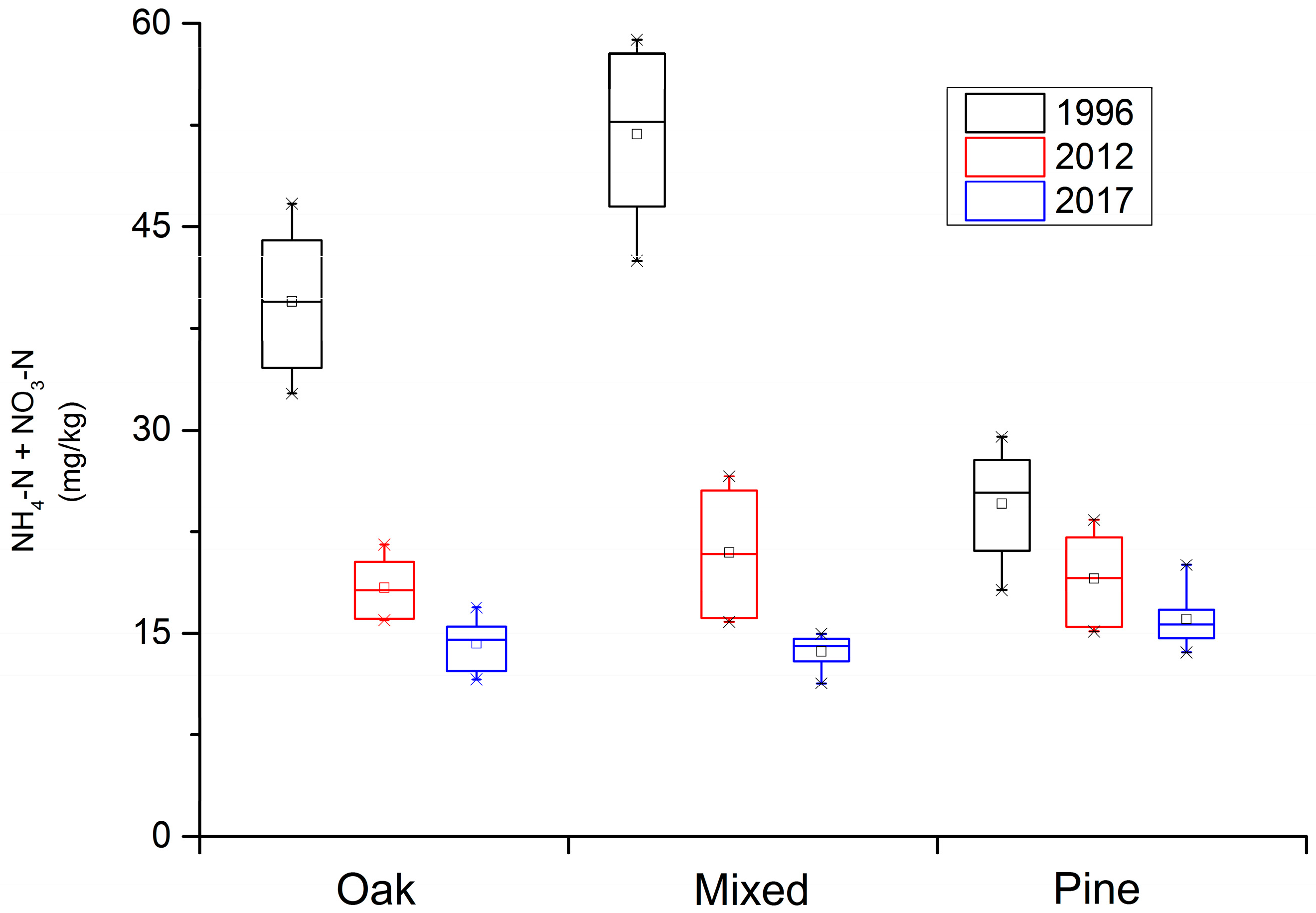
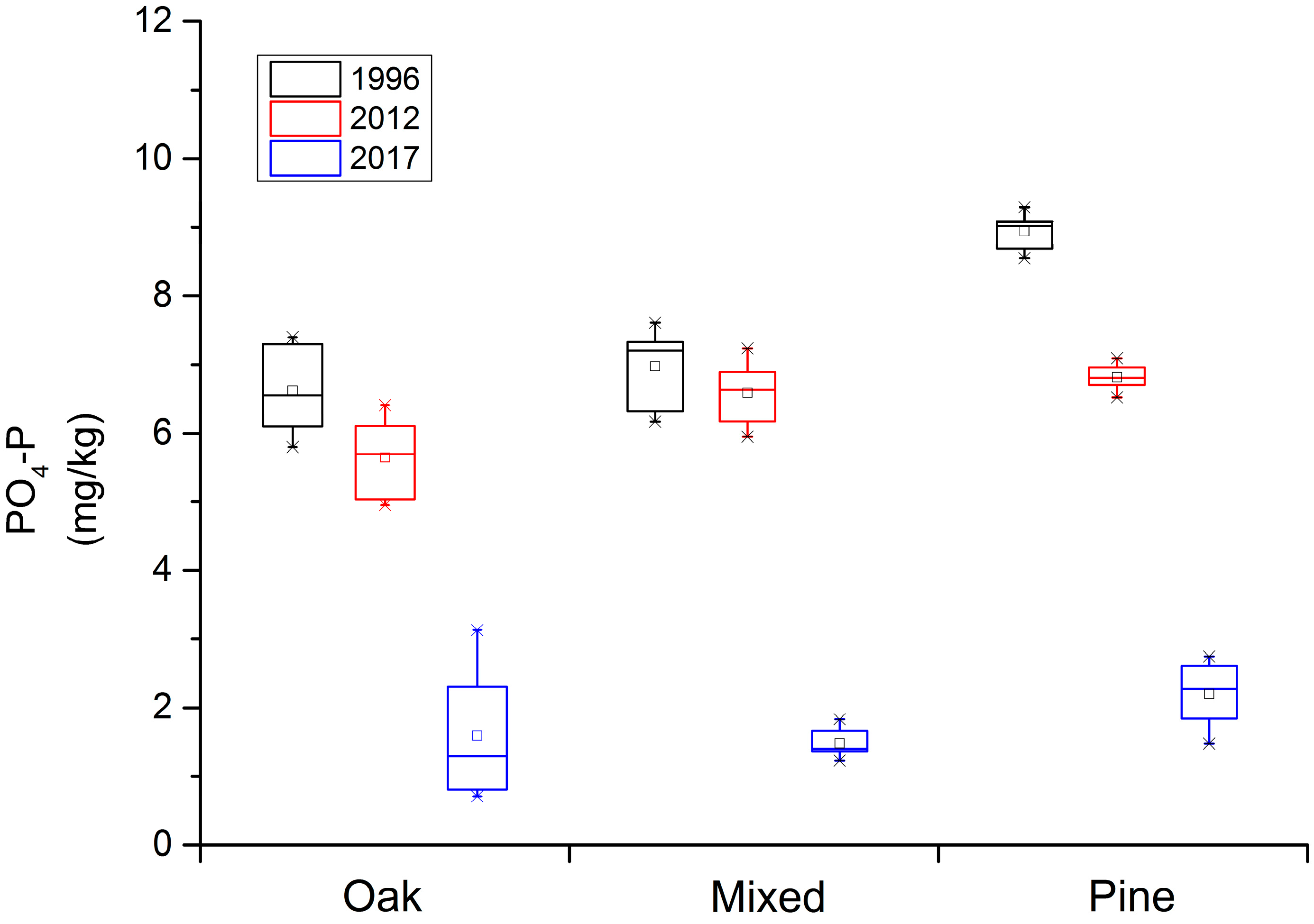
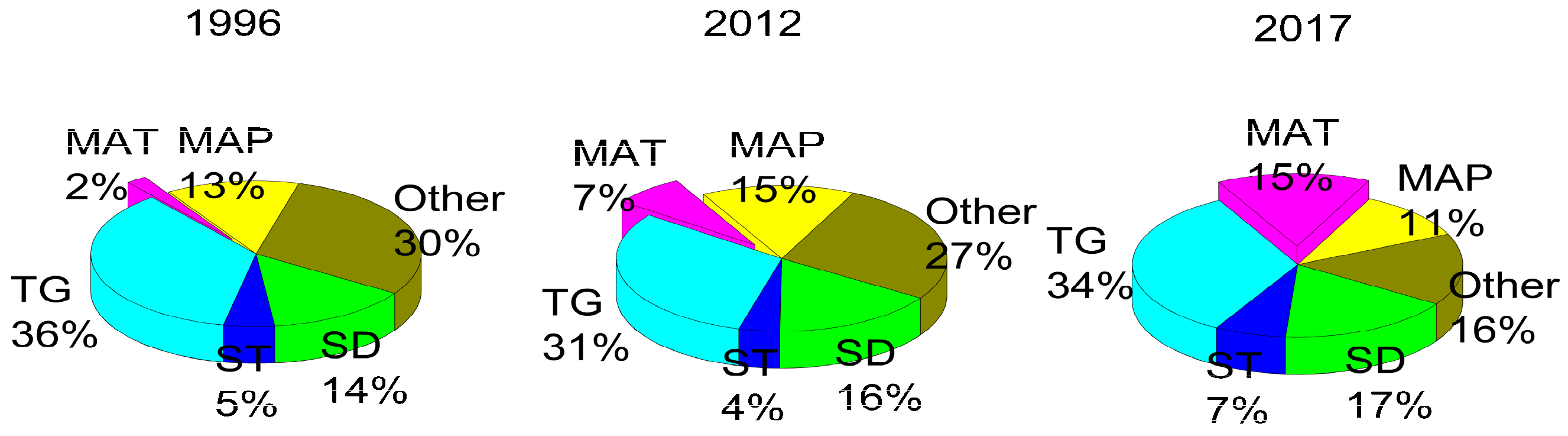
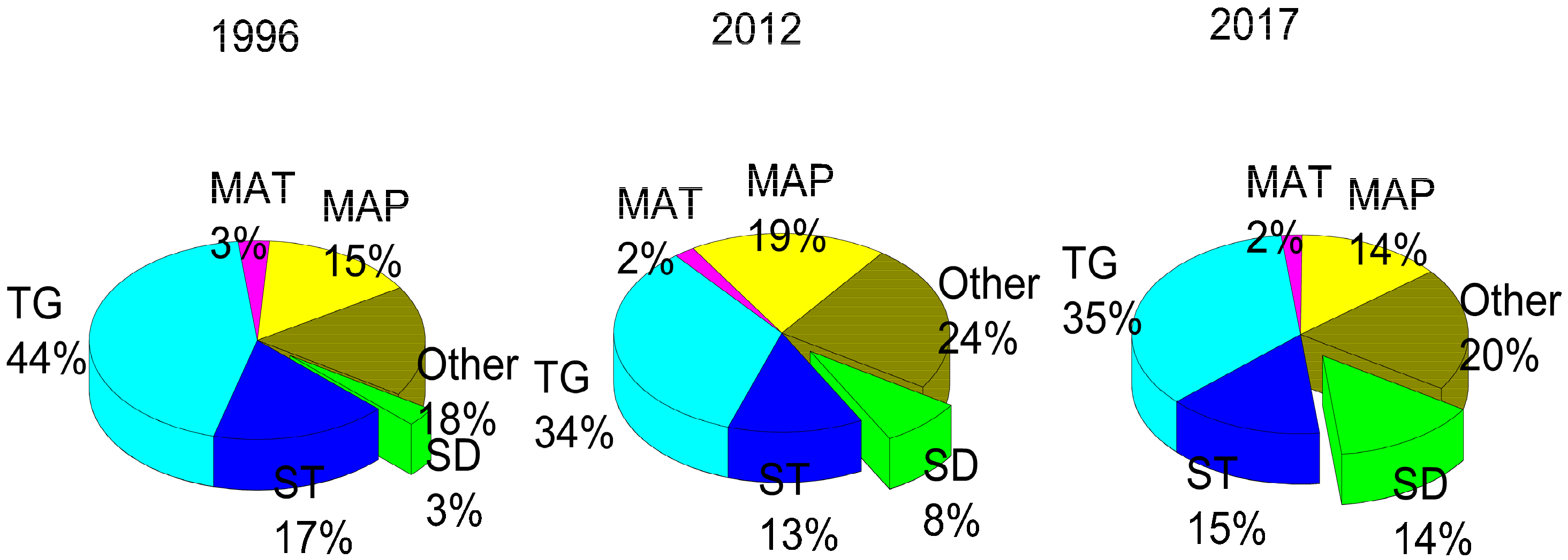
3. Results
3.1. Change in Soil Available N and P over Time
3.2. Factors Influencing the Abundance of Soil Available N and P
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oldeman, L.R.; Hakkeling, R.U.; Sombroek, W.G. World Map of the Status of Human-Induced Soil Degradation: An Explanatory Note; International Soil Reference and Information Centre: Wageningen, The Netherlands, 1991. [Google Scholar]
- Wang, T.; Yan, C.Z.; Song, X.; Li, S. Landsat images reveal trends in the Aeolian desertification in a source area for sand and dust storms in China’s Alashan plateau (1975–2007). Land Degrad. Dev. 2013, 24, 422–429. [Google Scholar] [CrossRef]
- Zhang, S.; Lei, R.; Liu, G.; Dang, K.; Shang, L.; Zhang, Y. Nutrient cycle in main types of forests at Huoditang Forest Region in the Qinling Mountains. J. Northwest For. Coll. 1996, 11, 115–120, (In Chinese with English Abstract). [Google Scholar]
- Hurni, H. Land degradation, famine, and land resource scenarios in Ethiopia. In World Soil Erosion and Conservation; Pimentel, D., Ed.; Cambridge University Press: Cambridge, UK, 1993; pp. 27–62. [Google Scholar]
- Shapiro, J. Environmental Degradation in China under Mao and Today: A Comparative Reflection. Glob. Environ. 2016, 9, 440–457. [Google Scholar] [CrossRef]
- Chen, J.; Chen, J.Z.; Tan, M.Z.; Gong, Z.T. Soil degradation: A global problem endangering sustainable development. J. Geogr. Sci. 2002, 12, 243–252. [Google Scholar]
- Smil, V. The Bad Earth. Environmental Degradation in China; ME Sharpe, Inc.: Armonk, NY, USA, 1984; Volume 9, pp. 332–333. [Google Scholar]
- Le Houérou, H.N. Climate change, drought and desertification. J. Arid Environ. 1996, 34, 133–185. [Google Scholar] [CrossRef]
- Schoenholtz, S.H.; Van Miegroet, H.; Burger, J.A. A review of chemical and physical properties as indicators of forest soil quality: Challenges and opportunities. For. Ecol. Manag. 2000, 138, 335–356. [Google Scholar] [CrossRef]
- Lemanceau, P.; Maron, P.A.; Mazurier, S.; Mougel, C.; Pivato, B.; Plassart, P.; Ranjard, L.; Revellin, C.; Tardy, V.; Wipf, D. Understanding and managing soil biodiversity: A major challenge in agroecology. Agron. Sustain. Dev. 2015, 35, 67–81. [Google Scholar] [CrossRef]
- Elser, J.J.; Andersen, T.; Baron, J.S.; Bergström, A.K.; Jansson, M.; Kyle, M.; Hessen, D.O. Shifts in lake N: P stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition. Science 2009, 326, 835–837. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Hobbie, S.E.; Lee, T.; Ellsworth, D.S.; West, J.B.; Tilman, D.; Trost, J. Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 2006, 440, 922–925. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Chapin, F.S.; Firestone, M.K.; Field, C.B.; Chiariello, N.R. Nitrogen limitation of microbial decomposition in a grassland under elevated CO2. Nature 2001, 409, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Koerselman, W.; Meuleman, A.F. The vegetation N: P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Conant, R.T.; Ryan, M.G.; Ågren, G.I.; Birge, H.E.; Davidson, E.A.; Eliasson, P.E.; Hyvönen, R. Temperature and soil organic matter decomposition rates–synthesis of current knowledge and a way forward. Glob. Chang. Biol. 2011, 17, 3392–3404. [Google Scholar] [CrossRef]
- Durán, J.; Morse, J.L.; Groffman, P.M.; Campbell, J.L.; Christenson, L.M.; Driscoll, C.T.; Mitchell, M.J. Climate change decreases nitrogen pools and mineralization rates in northern hardwood forests. Ecosphere 2016, 7, e01251. [Google Scholar] [CrossRef]
- Yuan, J.; Jose, S.; Zheng, X.F.; Cheng, F.; Hou, L.; Li, J.X.; Zhang, S.X. Dynamics of Coarse Woody Debris Characteristics in the Qinling Mountain Forests in China. Forests 2017, 8, 403. [Google Scholar] [CrossRef]
- Liu, G.S.; Jiang, N.H.; Zhang, L.D.; Liu, Z.L. Soil Physical and Chemical Analysis and Description of Soil Profiles; China Standard Methods Press: Beijing, China, 1996; pp. 24–266, (In Chinese with English Abstract). [Google Scholar]
- Jeppesen, E.; Kronvang, B.; Olesen, J.E.; Audet, J.; Søndergaard, M.; Hoffmann, C.C.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Larsen, S.E.; et al. Climate change effects on nitrogen loading from cultivated catchments in Europe: Implications for nitrogen retention, ecological state of lakes and adaptation. Hydrobiologia 2011, 663, 1–21. [Google Scholar] [CrossRef]
- Veraart, A.J.; De Klein, J.J.; Scheffer, M. Warming can boost denitrification disproportionately due to altered oxygen dynamics. PLoS ONE 2011, 6, e18508. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wei, X.; Zhang, S. Tree species diversity and identity effects on soil properties in the Huoditang area of the Qinling Mountains, China. Ecosphere 2017, 8, e01732. [Google Scholar] [CrossRef]
- Binkley, D.; Sollins, P.; Bell, R.; Sachs, D.; Myrold, D. Biogeochemistry of adjacent conifer and alder-conifer stands. Ecology 1992, 73, 2022–2033. [Google Scholar] [CrossRef]
- Lee, M.R.; Flory, S.L.; Phillips, R.P. Positive feedbacks to growth of an invasive grass through alteration of nitrogen cycling. Oecologia 2012, 170, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.E.; Richardson, J.P.; Canuel, E.A. Grazer Diversity Effects on Ecosystem Functioning in Seagrass beds. Ecol. Lett. 2003, 6, 637–645. [Google Scholar] [CrossRef]
- Hector, A.; Bagchi, R. Biodiversity and ecosystem multifunctionality. Nature 2007, 448, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Mikusiński, G. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansson, K.; Fröberg, M.; Helmisaari, H.S.; Kleja, D.B.; Olsson, B.A.; Olsson, M.; Persson, T. Carbon and nitrogen pools and fluxes above and below ground in spruce, pine and birch stands in southern Sweden. For. Ecol. Manag. 2013, 309, 28–35. [Google Scholar] [CrossRef]
- Bouwman, L.; Goldewijk, K.K.; Van Der Hoek, K.W.; Beusen, A.H.; Van Vuuren, D.P.; Willems, J.; Rufino, M.C.; Stehfest, E. Exploring global changes in nitrogen and phosphorus cycles in agriculture induced by livestock production over the 1900–2050 period. Proc. Natl. Acad. Sci. USA 2013, 110, 20882–20887. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, B.; Karonen, M.; Adamczyk, S.; Engström, M.T.; Laakso, T.; Saranpää, P. Tannins can slow-down but also speed-up soil enzymatic activity in boreal forest. Soil Biol. Biochem. 2017, 107, 60–67. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, B.; Zhang, J.; Mueller, C.; Cai, Z. Mechanisms of soil N dynamics following long-term application of organic fertilizers to subtropical rain-fed purple soil in China. Soil Biol. Biochem. 2015, 91, 222–231. [Google Scholar] [CrossRef]
- Bai, Z.; Li, H.; Yang, X.; Zhou, B.; Shi, X.; Wang, B.; Oenema, O. The critical soil P levels for crop yield, soil fertility and environmental safety in different soil types. Plant Soil 2013, 372, 27–37. [Google Scholar] [CrossRef]
- Niu, S.; Classen, A.T.; Dukes, J.S.; Kardol, P.; Liu, L.; Luo, Y.; Rustad, L.; Sun, J.; Tang, J.; Templer, P.H.; et al. Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle. Ecol. Lett. 2016, 19, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Smyth, C.E.; Titus, B.; Trofymow, J.A.; Moore, T.R.; Preston, C.M.; Prescott, C.E.; CIDET Working Group. Patterns of carbon, nitrogen and phosphorus dynamics in decomposing wood blocks in Canadian forests. Plant Soil 2016, 409, 459–477. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Plot | Latitude and Longitude | Altitude (m) | Aspect | Slope | Stand Density (Trees/ha) |
|---|---|---|---|---|---|
| Q. aliena var. acuteserrata 1# | 33°20′55″ N 108°23′54″ E | 1597 | 318° | 32° | 1183 |
| Q. aliena var. acuteserrata 2# | 33°19′20″ N 108°25′48″ E | 1641 | 14° | 28° | 1482 |
| Q. aliena var. acuteserrata 3# | 33°20′49″ N 108°25′58″ E | 1620 | 277° | 26° | 1232 |
| Q. aliena var. acuteserrata 4# | 33°19′10″ N 108°28′48″ E | 1640 | 260° | 23° | 1024 |
| Q. aliena var. acuteserrata 5# | 33°20′08″ N 108°28′12″ E | 1671 | 240° | 30° | 1086 |
| Q. aliena var. acuteserrata 6# | 33°20′42″ N 108°29′21″ E | 1534 | 218° | 18° | 1584 |
| P. armandii 1# | 33°19′26″ N 108°27′10″ E | 1410 | 288° | 34° | 1628 |
| P. armandii 2# | 33°19′30″ N 108°27′54″ E | 1460 | 198° | 32° | 1486 |
| P. armandii 3# | 33°22′54″ N 108°28′02″ E | 1483 | 245° | 27° | 1712 |
| P. armandii 4# | 33°23′10″ N 108°28′10″ E | 1532 | 194° | 22° | 1426 |
| P. armandii 5# | 33°21′10″ N 108°32′39″ E | 1540 | 243° | 29° | 1267 |
| P. armandii 6# | 33°22′01″ N 108°32′19″ E | 1983 | 245° | 15° | 1834 |
| Mixed forest 1# | 33°22′24″ N 108°27′10″ E | 1580 | 257° | 22° | 1354 |
| Mixed forest 2# | 33°19′01″ N 108°30′24″ E | 1481 | 204° | 24° | 1288 |
| Mixed forest 3# | 33°20′10″ N 108°30′42″ E | 1424 | 255° | 32° | 1078 |
| Mixed forest 4# | 33°21′05″ N 108°24′48″ E | 1582 | 73° | 26° | 1041 |
| Mixed forest 5# | 33°21′20″ N 108°26′50″ E | 1798 | 182° | 31° | 1121 |
| Mixed forest 6# | 33°23′18″ N 108°25′40″ E | 1627 | 202° | 21° | 1311 |
| Nutrient Elements | Forest Types | Year | ||
|---|---|---|---|---|
| 1996 | 2012 | 2017 | ||
| N | Q. aliena var. acuteserrata | 39.50 (5.68) A | 18.36 (1.54) B | 14.21 (1.02) C |
| Mixed forest | 51.75 (6.82) A | 20.87 (5.34) B | 13.54 (1.34) C | |
| P. armandii | 24.67 (4.44) A | 19.14 (1.78) B | 16.15 (1.23) C | |
| P | Q. aliena var. acuteserrata | 6.62 (0.64) A | 5.65 (0.58) B | 1.59 (0.95) C |
| Mixed forest | 6.95 (0.59) A | 6.57 (0.49) A | 1.46 (0.22) B | |
| P. armandii | 8.94 (0.27) A | 6.81 (0.20) B | 2.20 (0.49) C | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Yuan, J.; Zhang, T.; Hao, F.; Jose, S.; Zhang, S. Soil Degradation and the Decline of Available Nitrogen and Phosphorus in Soils of the Main Forest Types in the Qinling Mountains of China. Forests 2017, 8, 460. https://doi.org/10.3390/f8110460
Zheng X, Yuan J, Zhang T, Hao F, Jose S, Zhang S. Soil Degradation and the Decline of Available Nitrogen and Phosphorus in Soils of the Main Forest Types in the Qinling Mountains of China. Forests. 2017; 8(11):460. https://doi.org/10.3390/f8110460
Chicago/Turabian StyleZheng, Xiaofeng, Jie Yuan, Tong Zhang, Fan Hao, Shibu Jose, and Shuoxin Zhang. 2017. "Soil Degradation and the Decline of Available Nitrogen and Phosphorus in Soils of the Main Forest Types in the Qinling Mountains of China" Forests 8, no. 11: 460. https://doi.org/10.3390/f8110460
APA StyleZheng, X., Yuan, J., Zhang, T., Hao, F., Jose, S., & Zhang, S. (2017). Soil Degradation and the Decline of Available Nitrogen and Phosphorus in Soils of the Main Forest Types in the Qinling Mountains of China. Forests, 8(11), 460. https://doi.org/10.3390/f8110460