Cross-Talk between Physiological and Metabolic Adjustments Adopted by Quercus cerris to Mitigate the Effects of Severe Drought and Realistic Future Ozone Concentrations

 , , ,
, , ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Ecophysiological Analyses
2.3. Biochemical Analyses
2.4. Statistical Analyses
3. Results
3.1. Visible Injury
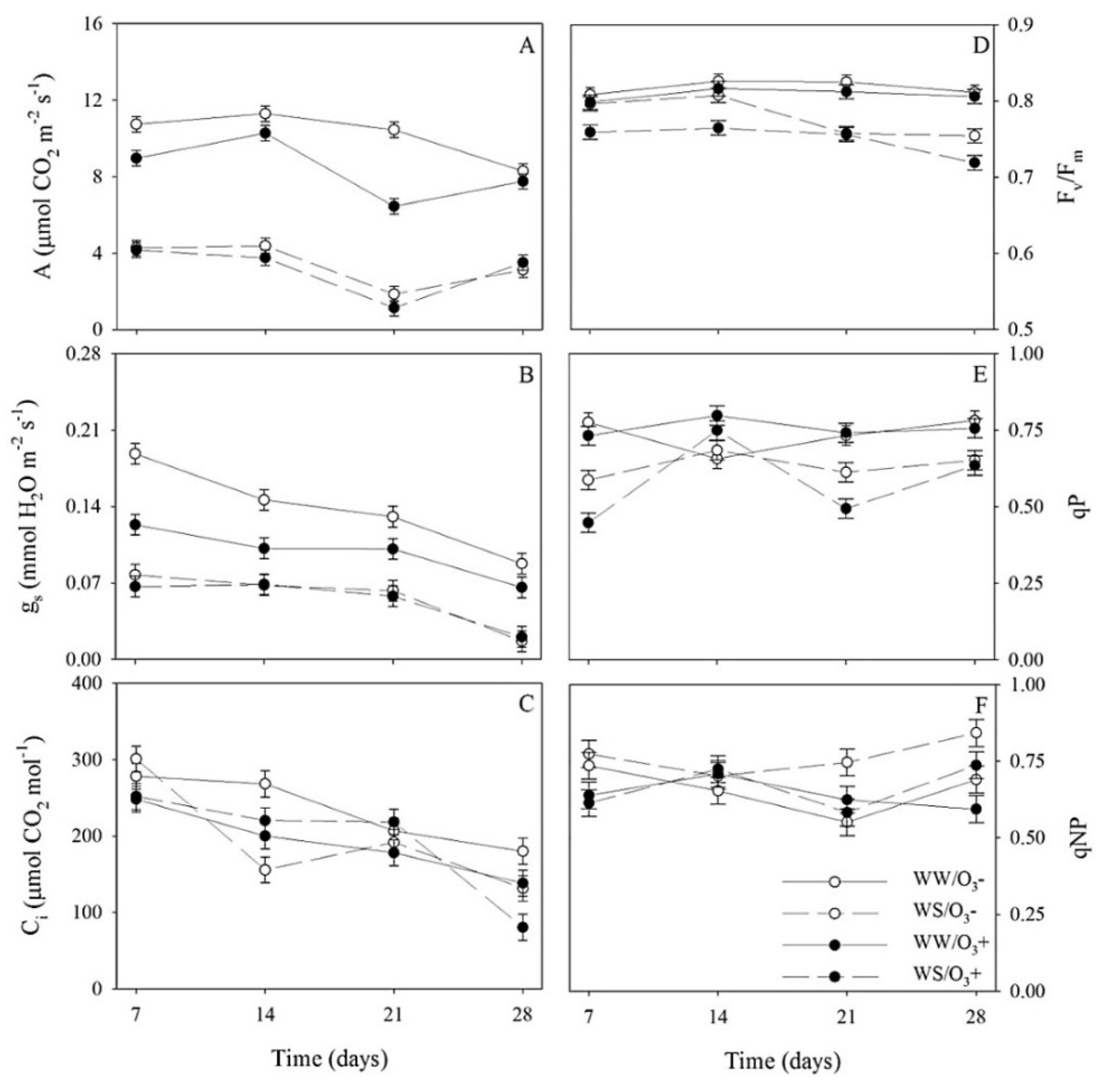
3.2. Weekly Profiles of Gas Exchanges and Chlorophyll a Fluorescence
3.3. Diurnal Profiles of Gas Exchanges and Chlorophyll a Fluorescence
3.4. Leaf Water Status and Osmolyte Contents
3.5. Leaf Pigments
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gray, S.B.; Brady, S.M. Plant developmental responses to climate change. Dev. Biol. 2016, 419, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Combourieu-Nebout, N.; Bertini, A.; Russo-Ermoli, E.; Peyron, O.; Klotz, S.; Montade, V.; Fauquette, S.; Allen, J.R.M.; Fusco, F.; Goring, S.; et al. Climate changes in the central Mediterranean and Italian vegetation dynamics since the Pliocene. Rev. Palaeobot. Palynol. 2015, 218, 127–147. [Google Scholar] [CrossRef]
- IPCC. Managing the risks of extreme events and disasters to advance climate change adaptation. In A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V., Stocker, T.F., Qin, D., Dokken, D.J., Ebi, K.L., Mastrandrea, M.D., Mach, K.J., Plattner, G.-K., Allen, S.K., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2012; p. 582. [Google Scholar]
- Bussotti, F.; Pollastrini, M.; Holland, V.; Brüggerman, W. Functional traits and adaptative capacity of European forests to climate change. Environ. Exp. Bot. 2015, 111, 91–113. [Google Scholar] [CrossRef]
- Bussotti, F.; Ferrini, F.; Pollastrini, M.; Fini, A. The challenge of Mediterranean sclerophyllous vegetation under climate change: From acclimation to adaptation. Environ. Exp. Bot. 2014, 103, 80–98. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Landi, M.; Massai, R.; Nali, C.; Guidi, L.; Lorenzini, G. Variations in physiological and biochemical traits of oak seedlings grown under drought and ozone stress. Physiol. Plant. 2015, 157, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Matyssek, R.; Le Thiec, D.; Löw, M.; Dizengremel, P.; Nunn, A.J.; Häberle, K.-H. Interactions between drought stress and O3 in forest trees. Plant Biol. 2005, 7, 1–7. [Google Scholar]
- Haberer, K.; Herbinger, K.; Alexou, M.; Rennenberg, H.; Tausz, M. Effects of drought and canopy ozone exposure on antioxidants in fine roots of mature European beech (Fagus sylvatica). Tree Physiol. 2008, 28, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Calderòn Guerrero, C.C.; Günthardt-Goerg, M.S.; Vollenweider, P. Correction: Foliar symptoms triggered by ozone stress in irrigated holm oaks from the city of Madrid, Spain. PLoS ONE 2014. [Google Scholar] [CrossRef]
- Pollastrini, M.; Desotgiu, R.; Camin, F.; Ziller, L.; Marzuoli, R.; Gerosa, G.; Bussotti, F. Intra-annual pattern of photosynthesis, growth and stable isotope partitioning in a poplar clone subjected to ozone and water stress. Water Air Soil Pollut. 2013, 224, 1761–1772. [Google Scholar] [CrossRef]
- Alonso, R.; Elvira, S.; González-Fernández, I.; Calvete, H.; García-Gómez, H.; Bermejo, V. Drought stress does not protect Quercus ilex L. from ozone effects: Results from a comparative study of two subspecies differing in ozone sensitivity. Plant Biol. 2014, 16, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Wingler, A.; Roitsch, T. Metabolic regulation of leaf senescence: Interactions of sugar signaling with biotic and abiotic stress responses. Plant Biol. 2008, 10, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 2012. [Google Scholar]
- Tognetti, R.; Raschi, A.; Béres, C.; Fenyvesi, A.; Ridder, H.-W. Comparison of water flow, cavitation and water status of Quercus cerris and Quercus petraea trees with special reference to computer tomography. Plant Cell Environ. 1996, 19, 928–938. [Google Scholar] [CrossRef]
- Nardini, A.; Lo Gullo, M.A.; Salleo, S. Competitive strategies for water availability in two Mediterranean Quercus species. Plant Cell Environ. 1999, 22, 109–116. [Google Scholar] [CrossRef]
- Valentini, R.; Epron, D.; De Angelis, P.; Matteucci, G.; Dreyer, E. In situ estimation of net CO2 assimilation, photosynthetic electron flow and photorespiration in Turkey oak (Q. cerris L.) leaves: Diurnal cycles under different levels of water supply. Plant Cell Environ. 1995, 18, 631–640. [Google Scholar] [CrossRef]
- D’Alessandro, C.M.; Saracino, A.; Borghetti, M. Thinning affects water use efficiency of hardwood saplings naturally recruited in a Pinus radiata D. Don plantation. For. Ecol. Manag. 2006, 222, 116–122. [Google Scholar] [CrossRef]
- Manes, F.; Vitale, M.; Donato, E.; Giannini, M.; Puppi, G. Different ability of three Mediterranean oak species to tolerate progressive dehydration stress. Photosynthetica 2006, 44, 387–393. [Google Scholar] [CrossRef]
- McDowell, N.G.; Sevanto, S. The mechanisms of carbon starvation: How, when, or does it even occur at all? New Phytol. 2010, 186, 264–266. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Piper, F.; Hoch, G. Physiological mechanisms of drought-induced tree mortality are far from being resolved. New Phytol. 2010, 186, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.; Ziegler, W.; Kolle, O.; Trumbore, S. Thirst beats hunger—Declining hydration during drought prevents carbon starvation in Norway spruce saplings. New Phytol. 2013, 200, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Struke, D.K.; Ferrini, F.; Fini, A.; Pennati, L. Relative growth and water use of seedlings from three Italian Quercus species. Arboric. Urban For. 2009, 35, 113–121. [Google Scholar]
- Pellegrini, E. PSII photochemistry is the primary target of oxidative stress imposed by ozone in Tilia americana. Urban For. Urban Green. 2014, 13, 94–102. [Google Scholar] [CrossRef]
- Lorenzini, G.; Medeghini Bonatti, P.; Nali, C.; Baroni Fornasiero, R. The protective effect of rust infection against ozone, sulphur dioxide and paraquat toxicity symptoms in broad bean. Physiol. Mol. Plant Path. 1994, 45, 263–279. [Google Scholar] [CrossRef]
- Turner, N.C.; Long, M.J. Errors arising from rapid water loss in the measurement of leaf water potential by the pressure chamber technique. Aust. J. Plant Physiol. 1980, 7, 527–537. [Google Scholar] [CrossRef]
- Gucci, R.; Lombardini, L.; Tattini, M. Analysis of leaf water relations in leaves of two olive (Olea europaea) cultivars differing in tolerance to salinity. Tree Physiol. 1997, 17, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Nali, C.; Pucciariello, C.; Mills, G.; Lorenzini, G. On the different sensitivity of white clover clones to ozone: Physiological and biochemical parameters in a multivariate approach. Water Air Soil Pollut. 2005, 164, 137–153. [Google Scholar] [CrossRef]
- Schreiber, U.; Schilwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Döring, A.S.; Pellegrini, E.; Campanella, A.; Trivellini, A.; Gennai, C.; Petersen, M.; Nali, C.; Lorenzini, G. How sensitive is Melissa officinalis to realistic ozone concentration? Plant Physiol. Biochem. 2014, 74, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, J.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Perata, P.; Matsukura, C.; Vernieri, P.; Yamaguchia, J. Repression of gibberellin-dependent signaling pathway in barley embryos. Plant Cell 1997, 9, 2197–2208. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, E.; Campanella, A.; Paolocci, M.; Trivellini, A.; Gennai, C.; Muganu, M.; Nali, C.; Lorenzini, G. Functional leaf traits and diurnal dynamics of photosynthetic parameters predict the behavior of grapevine varieties towards ozone. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Chater, C.; Kamisugi, Y.; Movahedi, M.; Fleming, A.; Cuming, A.C.; Gray, J.E.; Beerling, D.J. Regulatory mechanism controlling stomatal behavior conserved across 400 million years of land plant evolution. Curr. Biol. 2011, 21, 1025–1029. [Google Scholar] [CrossRef] [PubMed]
- Maselli, F.; Cherubini, P.; Chiesi, M.; Gilabert, M.A.; Lombardi, F.; Moreno, A.; Teobaldelli, M.; Tognetti, R. Start of the dry season as a main determinant of inter-annual Mediterranean forest production variations. Agr. For. Meteorol. 2014, 194, 197–206. [Google Scholar] [CrossRef]
- Gratani, L. Plant phenotypic plasticity in response to environmental factors. Advances Bot. 2014. [Google Scholar] [CrossRef]
- Mitchell, P.J.; O’Grady, A.P.; Tissue, D.T.; White, D.A.; Ottenschlager, M.L.; Pinkard, E.A. Drought response strategies define the relative contributions of hydraulic dysfunction and carbohydrate depletion during tree mortality. New Phytol. 2013, 197, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Savi, T.; Casolo, V.; Luglio, J.; Bertuzzi, S.; Trifilò, P.; Lo Gullo, M.A.; Nardini, A. Species-specific reversal of stem xylem embolism after a prolonged drought correlates to endpoint concentration of soluble sugars. Plant Physiol. Biochem. 2016, 106, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, M.K.; Zhang, Y.; Kreidler, N.; Sun, S.; Ardy, R.; Cao, K.; Sack, L. Global analysis of plasticity in turgor loss point, a key drought tolerance trait. Ecol. Lett. 2014, 17, 1580–1590. [Google Scholar] [CrossRef] [PubMed]
- Garnier, E.; Berger, A. Testing water potential in peach trees as an indicator of water stress. J. Hortic. Sci. 1985, 60, 47–56. [Google Scholar] [CrossRef]
- Gomes, F.P.; Oliva, M.A.; Mielke, M.S.; Almeida, A.-A.F.; Aquino, L.A. Osmotic adjustment, proline accumulation and cell membrane stability in leaves of Cocos nucifera submitted to drought stress. Sci. Hort. 2010, 126, 379–384. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Guo, K.; Fan, D.; Li, G.; Zheng, Y.; Yu, L.; Yang, R. Effect of drought on pigments, osmotic adjustment and antioxidant enzymes in six woody plant species in karst habitats of southwestern China. Environ. Exp. Bot. 2011, 71, 174–183. [Google Scholar] [CrossRef]
- Ugolini, F.; Bussotti, F.; Raschi, A.; Tognetti, R.; Ennos, A.R. Physiological performance and biomass production of two ornamental shrub species under deficit irrigation. Trees 2015, 29, 407–422. [Google Scholar] [CrossRef]
- Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S.; Boukhris, M.; Ben Abdallah, F. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Wingler, A.; Purdy, S.; MacLean, J.A.; Pourtau, N. The role of sugars in integrating environmental signals during the regulation of leaf senescence. J. Exp. Bot. 2006, 57, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 1–875. [Google Scholar] [CrossRef] [PubMed]
- Versules, P.E.; Bray, E.A. Role of abscisic acid (ABA) and Arabidopsis thaliana ABA-insensitive loci in low water potential-induced ABA and proline accumulation. J. Exp. Bot. 2006, 57, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinherio, C. How plant cope with water stress in the field. Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, E.; Anselmi, N.; Franceschini, A. Pre-exposure to ozone predispose oak leaves to attacks by Diplodia corticola and Biscognauxia mediterranea. Sci. World J. 2007, 7, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S.; Queval, G.; Foyer, C.H. The impact of global change factors on redox signaling underpinning stress tolerance. Plant Physiol. 2013, 161, 5–19. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Effects | d.f. | A | gs | Ci | Fv/Fm | qP | qNP |
|---|---|---|---|---|---|---|---|
| Drought | 1 | 1971.33 *** | 92.60 *** | 5.98 * | 90.27 *** | 40.70 *** | 8.67 * |
| Ozone | 1 | 60.89 *** | 10.65 * | 8.88 * | 14.36 ** | 0.55 ns | 6.95 * |
| Time | 3 | 32.24 *** | 35.00 *** | 44.16 *** | 7.26 ** | 7.53 ** | 3.13 * |
| Drought × Ozone | 1 | 31.88 *** | 8.08 * | 7.06 * | 3.82 ns | 2.81 ns | 3.69 ns |
| Drought × Time | 3 | 6.36 ** | 2.13 ns | 4.58 * | 5.34 ** | 9.45 *** | 1.97 ns |
| Ozone × Time | 3 | 5.71 ** | 1.65 ns | 2.06 ns | 0.82 ns | 7.24 ** | 2.83 ns |
| Drought × Ozone × Time | 3 | 2.44 ns | 0.61 ns | 4.28 * | 1.24 ns | 0.85 ns | 1.36 ns |
| Treatment | PDΨw (–MPa) | Ψπ (–MPa) | MDA (nmol g−1 DW) | RWC (%) | EL (%) |
|---|---|---|---|---|---|
| WW/O3− | 0.5 a | 3.6 a | 119 a | 79 | 26.2 a |
| WS/O3− | 2.7 c | 4.8 c | 139 c | 73 | 51.4 c |
| WW/O3+ | 0.9 a | 4.0 ab | 118 a | 75 | 26.6 a |
| WS/O3+ | 1.5 b | 4.2 b | 125 b | 73 | 36.1 b |
| SEM | 0.18 | 0.14 | 1.5 | 1.4 | 2.02 |
| Drought | *** | ** | *** | * | *** |
| Ozone | * | ns | ** | ns | ** |
| Drought × Ozone | ** | ** | ** | ns | ** |
| Treatment | Proline | ABA | Hexoses |
|---|---|---|---|
| WW/O3− | 0.78 | 0.14 a | 584 a |
| WS/O3− | 1.65 | 0.37 c | 561 a |
| WW/O3+ | 0.36 | 0.13 a | 561 a |
| WS/O3+ | 1.13 | 0.20 b | 712 b |
| SEM | 0.047 | 0.015 | 12.3 |
| Drought | *** | *** | *** |
| Ozone | *** | ** | *** |
| Drought × Ozone | ns | * | *** |
| Treatment | Car/Chl (µmol µmol−1) | VAZ/Chl (µmol µmol−1) | β-Carotene (µmol g−1 DW) | α-Tocopherol (µmol g−1 DW) |
|---|---|---|---|---|
| WW/O3− | 0.97 b | 0.27 | 19.2 a | 3.5 b |
| WS/O3− | 1.04 c | 0.29 | 22.8 b | 5.3 d |
| WW/O3+ | 0.91 a | 0.24 | 24.4 b | 4.4 c |
| WS/O3+ | 1.06 c | 0.28 | 19.9 a | 2.8 a |
| SEM | 0.034 | 0.008 | 1.22 | 0.08 |
| Drought | *** | ** | ns | ns |
| Ozone | ns | * | ns | *** |
| Drought × Ozone | * | ns | *** | *** |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Guidi, L.; Lorenzini, G.; Massai, R.; Nali, C.; Landi, M. Cross-Talk between Physiological and Metabolic Adjustments Adopted by Quercus cerris to Mitigate the Effects of Severe Drought and Realistic Future Ozone Concentrations. Forests 2017, 8, 148. https://doi.org/10.3390/f8050148
Cotrozzi L, Remorini D, Pellegrini E, Guidi L, Lorenzini G, Massai R, Nali C, Landi M. Cross-Talk between Physiological and Metabolic Adjustments Adopted by Quercus cerris to Mitigate the Effects of Severe Drought and Realistic Future Ozone Concentrations. Forests. 2017; 8(5):148. https://doi.org/10.3390/f8050148
Chicago/Turabian StyleCotrozzi, Lorenzo, Damiano Remorini, Elisa Pellegrini, Lucia Guidi, Giacomo Lorenzini, Rossano Massai, Cristina Nali, and Marco Landi. 2017. "Cross-Talk between Physiological and Metabolic Adjustments Adopted by Quercus cerris to Mitigate the Effects of Severe Drought and Realistic Future Ozone Concentrations" Forests 8, no. 5: 148. https://doi.org/10.3390/f8050148
APA StyleCotrozzi, L., Remorini, D., Pellegrini, E., Guidi, L., Lorenzini, G., Massai, R., Nali, C., & Landi, M. (2017). Cross-Talk between Physiological and Metabolic Adjustments Adopted by Quercus cerris to Mitigate the Effects of Severe Drought and Realistic Future Ozone Concentrations. Forests, 8(5), 148. https://doi.org/10.3390/f8050148