Effectiveness of Diesel as a Mammal Repellent for Direct Seeding of Acorns


Abstract
1. Introduction
2. Materials and Methods
2.1. Acorn Predation Experiment
2.2. Nursery Experiment
2.3. Statistical Analyses
3. Results and Discussion
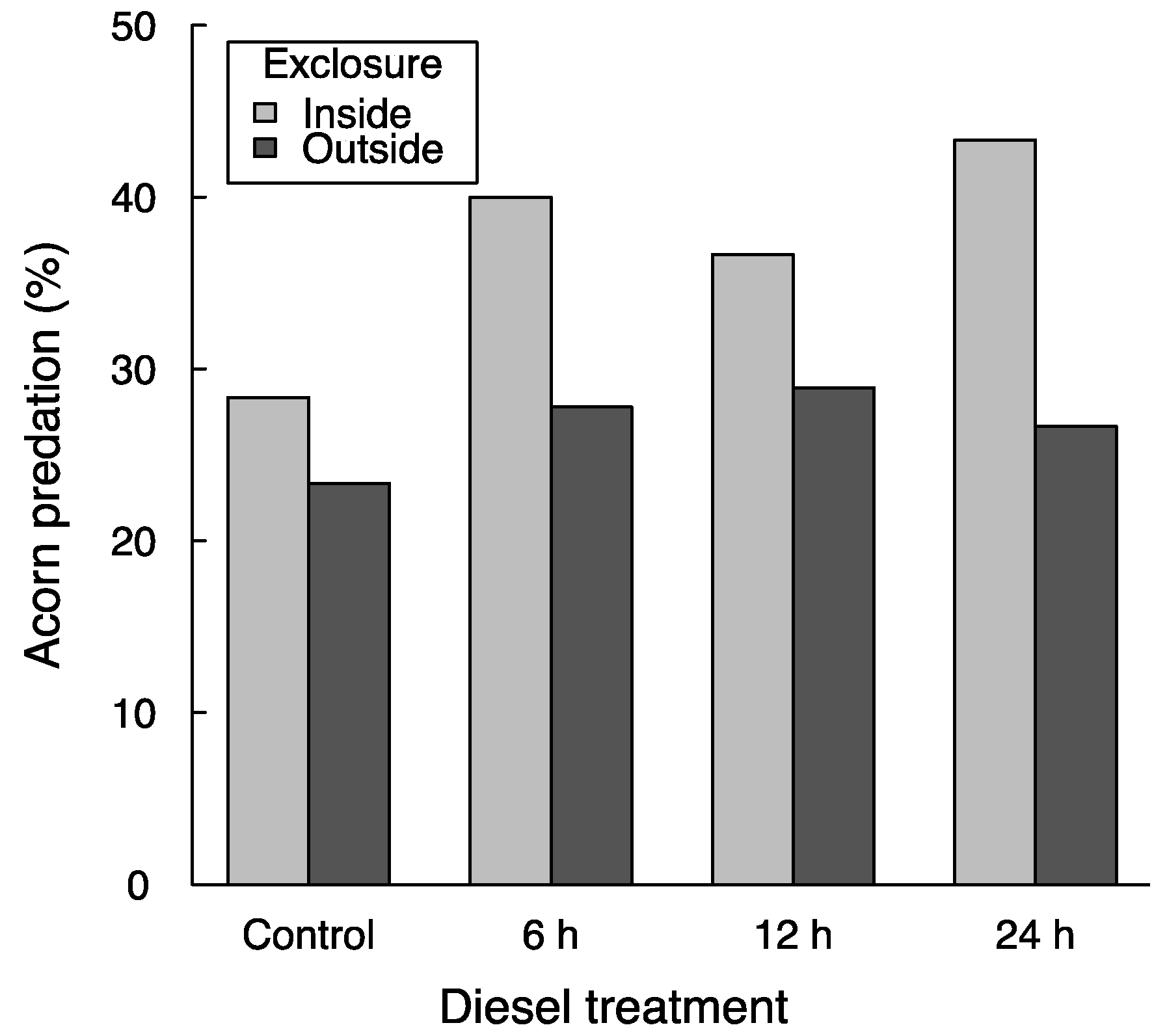
3.1. Acorn Predation
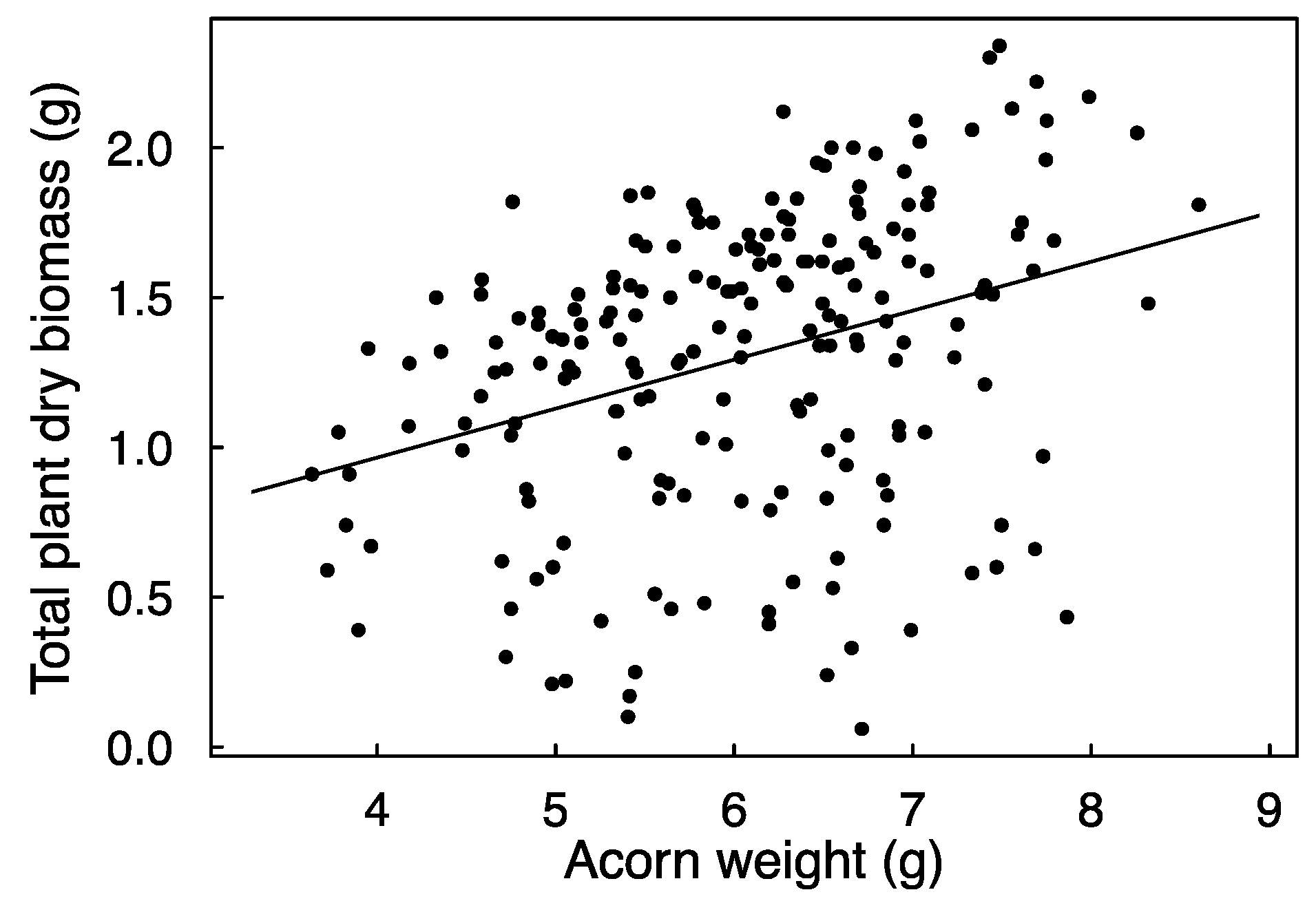
3.2. Seedling Emergence and Growth
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Twedt, D.J.; Wilson, R.R. Development of oak plantations established for wildlife. For. Ecol. Manag. 2002, 162, 287–298. [Google Scholar] [CrossRef]
- Dey, D.C.; Jacobs, D.; Mcnabb, K.; Miller, G.; Baldwin, V.; Foster, G. Artificial Regeneration of Major Oak (Quercus) Species in the Eastern United States—A Review of the Literature. For. Sci. 2008, 54, 77–106. [Google Scholar]
- Benayas, J.M.R.; Navarro, J.; Espigares, T.; Nicolau, J.M.; Zavala, M.A. Effects of artificial shading and weed mowing in reforestation of Mediterranean abandoned cropland with contrasting Quercus species. For. Ecol. Manag. 2005, 212, 302–314. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Castro, J.; Delgado-Capel, M.J.; Molinas-González, C.; Pulgar, M.; Marañón-Jiménez, S.; Delgado-Huertas, A.; Querejeta, J.I. Restoring for the present or restoring for the future: Enhanced performance of two sympatric oaks (Quercus ilex and Quercus pyrenaica) above the current forest limit. Restor. Ecol. 2015, 23, 936–946. [Google Scholar] [CrossRef]
- Valdecantos, A.; Cortina, J.; Vallejo, V.R. Nutrient status and field performance of tree seedlings planted in Mediterranean degraded areas. Ann. For. Sci. 2006, 63, 249–256. [Google Scholar] [CrossRef]
- Zadworny, M.; Jagodziński, A.M.; Łakomy, P.; Ufnalski, K.; Oleksyn, J. The silent shareholder in deterioration of oak growth: Common planting practices affect the long-term response of oaks to periodic drought. For. Ecol. Manag. 2014, 318, 133–141. [Google Scholar] [CrossRef]
- Tsakaldimi, M.; Tsitsoni, T.; Ganatsas, P.; Zagas, T. A comparison of root architecture and shoot morphology between naturally regenerated and container-grown seedlings of Quercus ilex. Plant Soil 2009, 324, 103–113. [Google Scholar] [CrossRef]
- Allen, J.A.; Keeland, B.D.; Stanturf, J.A.; Clewell, A.F.; Kennedy, H.E., Jr. A Guide to Bottomland Hardwood Restoration; General Technical Report SRS-40; U.S. Department of Agriculture, U.S. Department of the Interior and Forest Service: Washington, DC, USA; U.S. Geological Survey: Reston, VA, USA, 2001; p. vii-132.
- Bullard, S.H.; Hodges, J.D.; Johnson, R.L.; Straka, T.J. Economics of direct seeding and planting for establishing oak stands on old-field sites in the South. South. J. Appl. For. 1992, 16, 35–40. [Google Scholar]
- Crawley, M.J.; Long, C.R. Alternate bearing, predator satiation and seedling recruitment in Quercus robur L. J. Ecol. 1995, 83, 683–696. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Castro, J.; Puerta-Piñero, C.; Rey Benayas, J.M. Suitability of the management of habitat complexity, acorn burial depth, and a chemical repellent for post-fire reforestation of oaks. Ecol. Eng. 2013, 53, 15–22. [Google Scholar] [CrossRef]
- Pérez-Ramos, I.M.; Marañón, T. Factors affecting post-dispersal seed predation in two coexisting oak species: Microhabitat, burial and exclusion of large herbivores. For. Ecol. Manag. 2008, 255, 3506–3514. [Google Scholar] [CrossRef]
- Puerta-Piñero, C.; Sánchez-Miranda, A.; Leverkus, A.; Castro, J. Management of burnt wood after fire affects post-dispersal acorn predation. For. Ecol. Manag. 2010, 260, 345–352. [Google Scholar] [CrossRef]
- Birkedal, M.; Löf, M.; Olsson, G.E.; Bergsten, U. Effects of granivorous rodents on direct seeding of oak and beech in relation to site preparation and sowing date. For. Ecol. Manag. 2010, 259, 2382–2389. [Google Scholar] [CrossRef]
- Farlee, L.D. Direct seeding of fine hardwood tree species. In Managing Fine Hardwoods after a Half Century of Research, Proceedings of the Seventh Walnut Council Research Symposium, Madison, WI, USA, 1–3 August 2011; General Technical Report NRS-P-115; Van Sambeek, J.W., Jackson, E.A., Coggeshall, M.V., Thomas, A.L., Michler, C.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA; Northern Research Station: Newtown Square, PA, USA, 2013; pp. 31–47. [Google Scholar]
- Madsen, P.; Löf, M. Reforestation in southern Scandinavia using direct seeding of oak (Quercus robur L.). Forestry 2005, 78, 55–64. [Google Scholar] [CrossRef]
- Smith, H.G.; Hopmans, P.; Sheridan, G.J.; Lane, P.N.J.; Noske, P.J.; Bren, L.J. Impacts of wildfire and salvage harvesting on water quality and nutrient exports from radiata pine and eucalypt forest catchments in South-Eastern Australia. For. Ecol. Manag. 2012, 263, 160–169. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Rojo, M.; Castro, J. Habitat complexity and individual acorn protectors enhance the post-fire restoration of oak forests via seed sowing. Ecol. Eng. 2015, 83, 276–280. [Google Scholar] [CrossRef]
- Reque, J.A.; Martin, E. Designing acorn protection for direct seeding of quercus species in high predation areas. For. Syst. 2015, 24, e018. [Google Scholar] [CrossRef]
- Bäumler, W.; Mielke, H.; Zahner, V. Behandlung von Eicheln mit Repellentien gegen Nagetierschäden. Curr. J. Pest Sci. 1990, 63, 15–19. [Google Scholar] [CrossRef]
- Puerta-Piñero, C. Intermediate spatial variations on acorn predation shapes Holm oak establishment within a Mediterranean landscape context. Plant Ecol. 2010, 210, 213–224. [Google Scholar] [CrossRef]
- Gómez, J.M.; Puerta-Piñero, C.; Schupp, E.W. Effectiveness of rodents as local seed dispersers of Holm oaks. Oecologia 2008, 155, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Leverkus, A.B.; Fuster, F. A new device to foster oak forest restoration via seed sowing. New For. 2015, 46, 919–929. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M.; Gómez-Aparicio, L. Benefits of using shrubs as nurse plants for reforestation in Mediterranean mountains: A 4-year study. Restor. Ecol. 2004, 12, 352–358. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Use of shrubs as nurse plants: A new technique for reforestation in Mediterranean mountains. Restor. Ecol. 2002, 10, 297–305. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 10 July 2017).
- Bates, D.; Mächler, M.; Bolker, B. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models; R Package Version 3.1–128; 2016; Available online: http://CRAN.R-project.org/package=nlme (accessed on 10 July 2017).
- Therneau, T.M. coxme: Mixed Effects Cox Models; R Package Version 2.2–5; 2015; Available online: http://CRAN.R-project.org/package=coxme (accessed on 10 July 2017).
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000. [Google Scholar]
- Crawley, M.J. The R Book, 2nd ed.; Wiley & Sons: New Delhi, India, 2013; ISBN 9780262232586. [Google Scholar]
- Gómez, J.M.; Hódar, J.A. Wild boars (Sus scrofa) affect the recruitment rate and spatial distribution of holm oak (Quercus ilex). For. Ecol. Manag. 2008, 256, 1384–1389. [Google Scholar] [CrossRef]
- Gómez, J.M. Importance of microhabitat and acorn burial on Quercus ilex early recruitment: Non-additive effects on multiple demographic processes. Plant Ecol. 2004, 172, 287–297. [Google Scholar] [CrossRef]
- Wróbel, A.; Bogdziewicz, M. It is raining mice and voles: Which weather conditions influence the activity of Apodemus flavicollis and Myodes glareolus? Eur. J. Wildl. Res. 2015, 61, 475–478. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Term | df | Emergence Probability | Emergence Time | Stem Height | Seedling Dry Biomass | ||||
|---|---|---|---|---|---|---|---|---|---|
| χ2 | p | χ2 | p | χ2 | p | χ2 | p | ||
| Diesel (D) | 3 | 17.71 | <0.001 | 6.26 | 0.10 | 3.21 | 0.36 | 5.83 | 0.12 |
| Acorn weight (W) | 1 | 2.03 | 0.15 | 0.43 | 0.51 | 2.60 | 0.11 | 29.39 | <0.001 |
| D × W | 3 | --- 1 | --- 1 | 3.66 | 0.30 | 1.15 | 0.77 | 1.51 | 0.68 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leverkus, A.B.; Carrión, M.; Molina-Morales, M.; Castro, J. Effectiveness of Diesel as a Mammal Repellent for Direct Seeding of Acorns. Forests 2017, 8, 276. https://doi.org/10.3390/f8080276
Leverkus AB, Carrión M, Molina-Morales M, Castro J. Effectiveness of Diesel as a Mammal Repellent for Direct Seeding of Acorns. Forests. 2017; 8(8):276. https://doi.org/10.3390/f8080276
Chicago/Turabian StyleLeverkus, Alexandro B., Manuel Carrión, Mercedes Molina-Morales, and Jorge Castro. 2017. "Effectiveness of Diesel as a Mammal Repellent for Direct Seeding of Acorns" Forests 8, no. 8: 276. https://doi.org/10.3390/f8080276
APA StyleLeverkus, A. B., Carrión, M., Molina-Morales, M., & Castro, J. (2017). Effectiveness of Diesel as a Mammal Repellent for Direct Seeding of Acorns. Forests, 8(8), 276. https://doi.org/10.3390/f8080276