
Effects of Drought on Xylem Anatomy and Water-Use Efficiency of Two Co-Occurring Pine Species

 and
and
Abstract
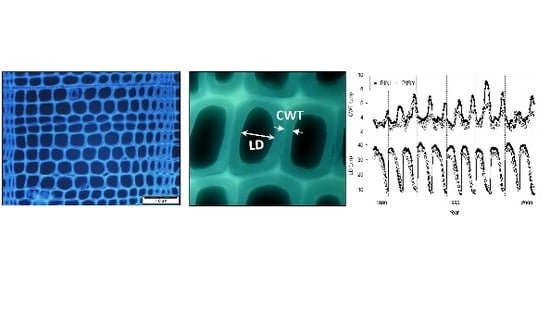
:
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Dendrochronological and Wood Anatomical Methods
2.3. Stable Isotopes
2.4. Statistical Analysis
3. Results
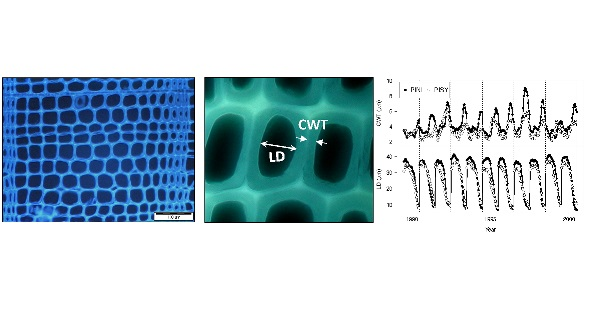
3.1. Changes in Climate
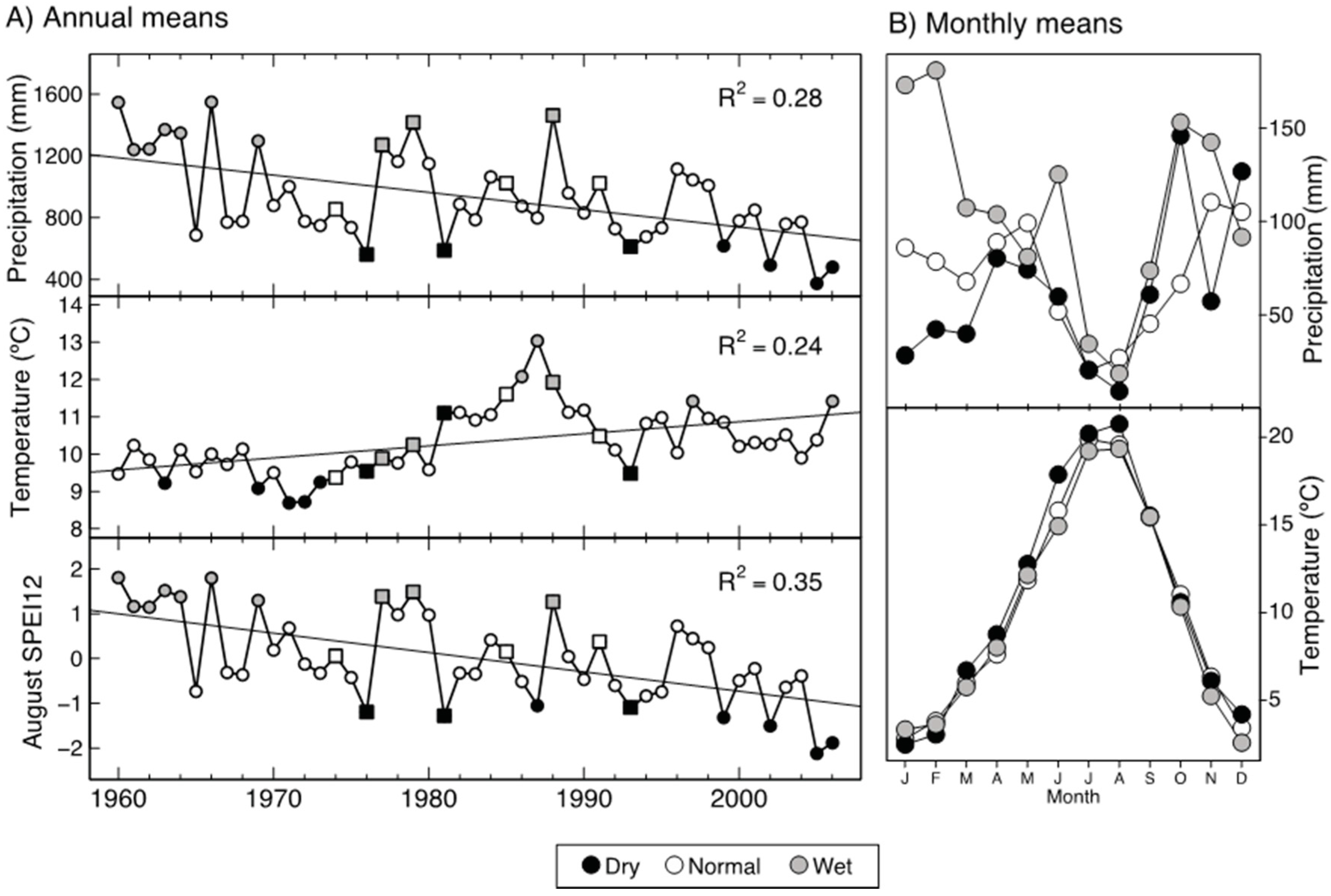
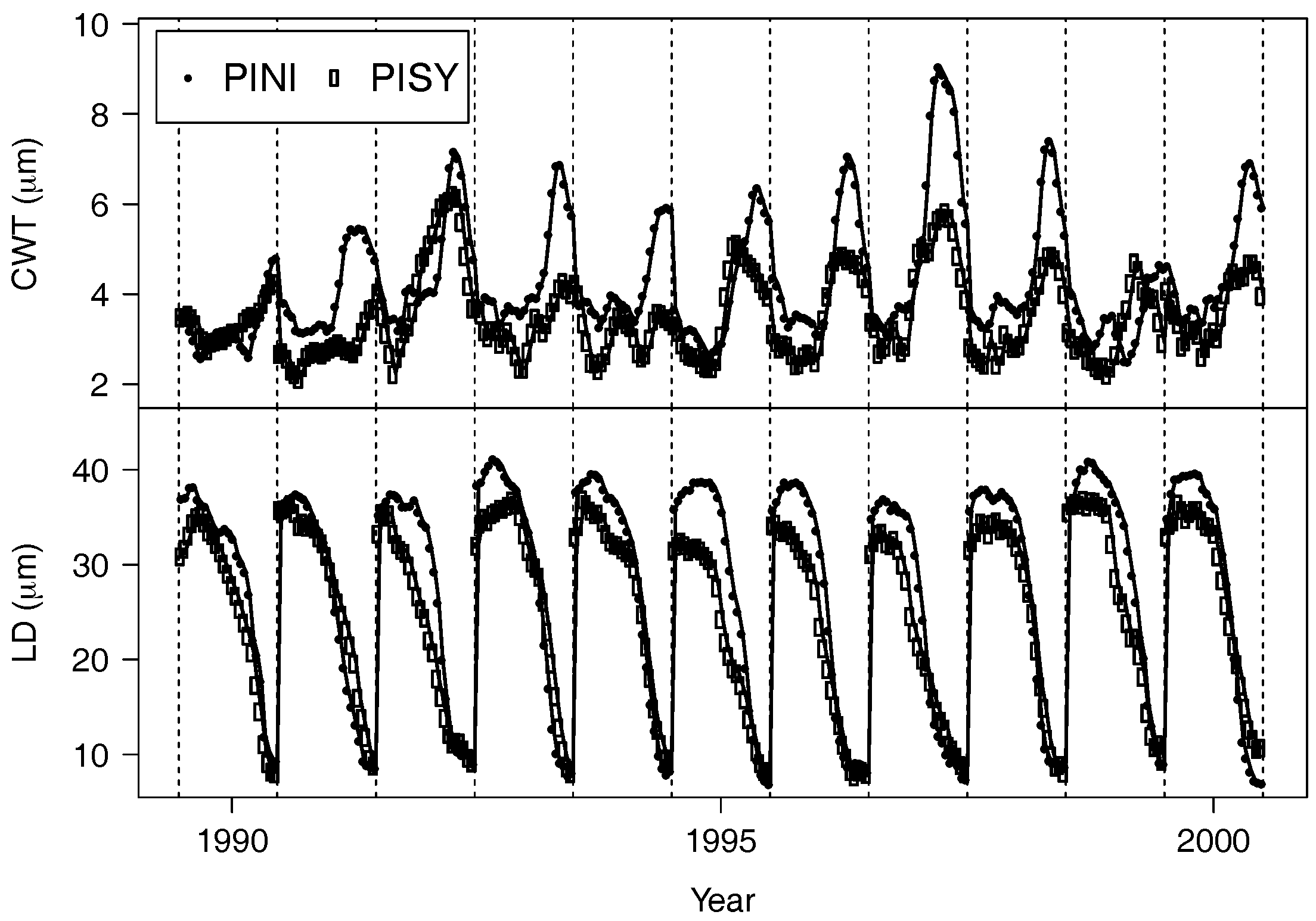
3.2. Interspecific Growth and Anatomical Differences
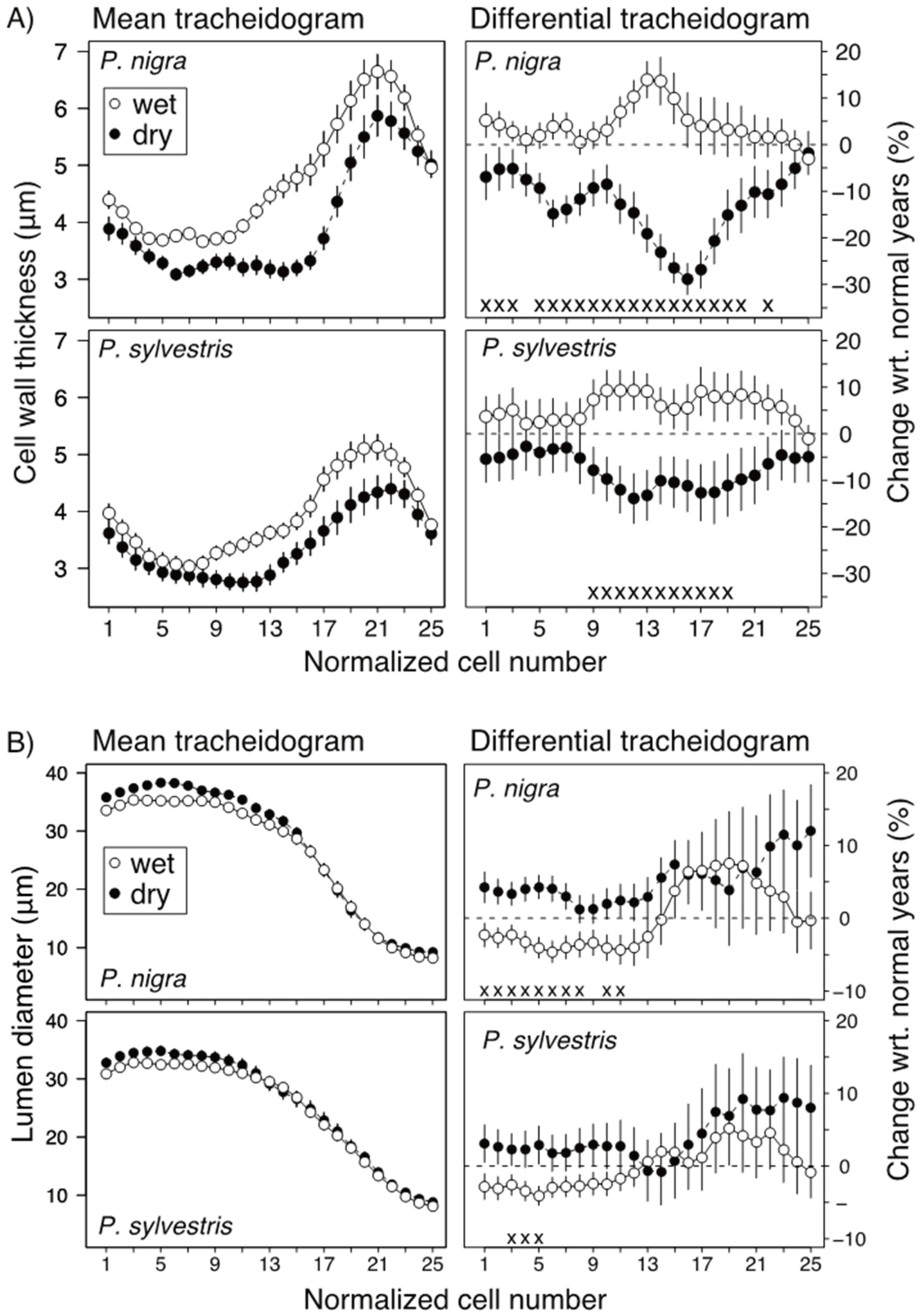
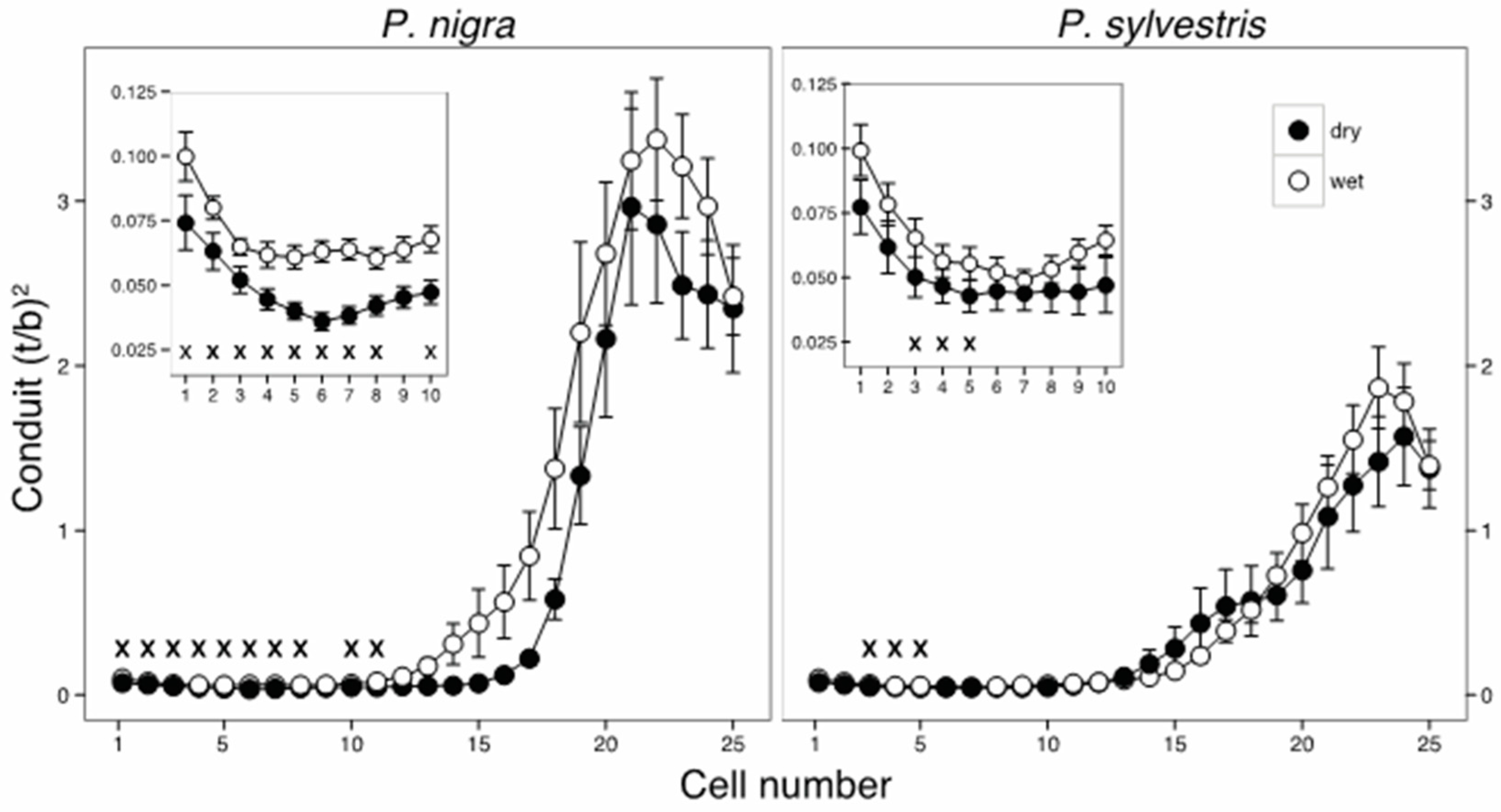
3.3. Effect of Moisture Availability on Tree Growth and Anatomical Traits
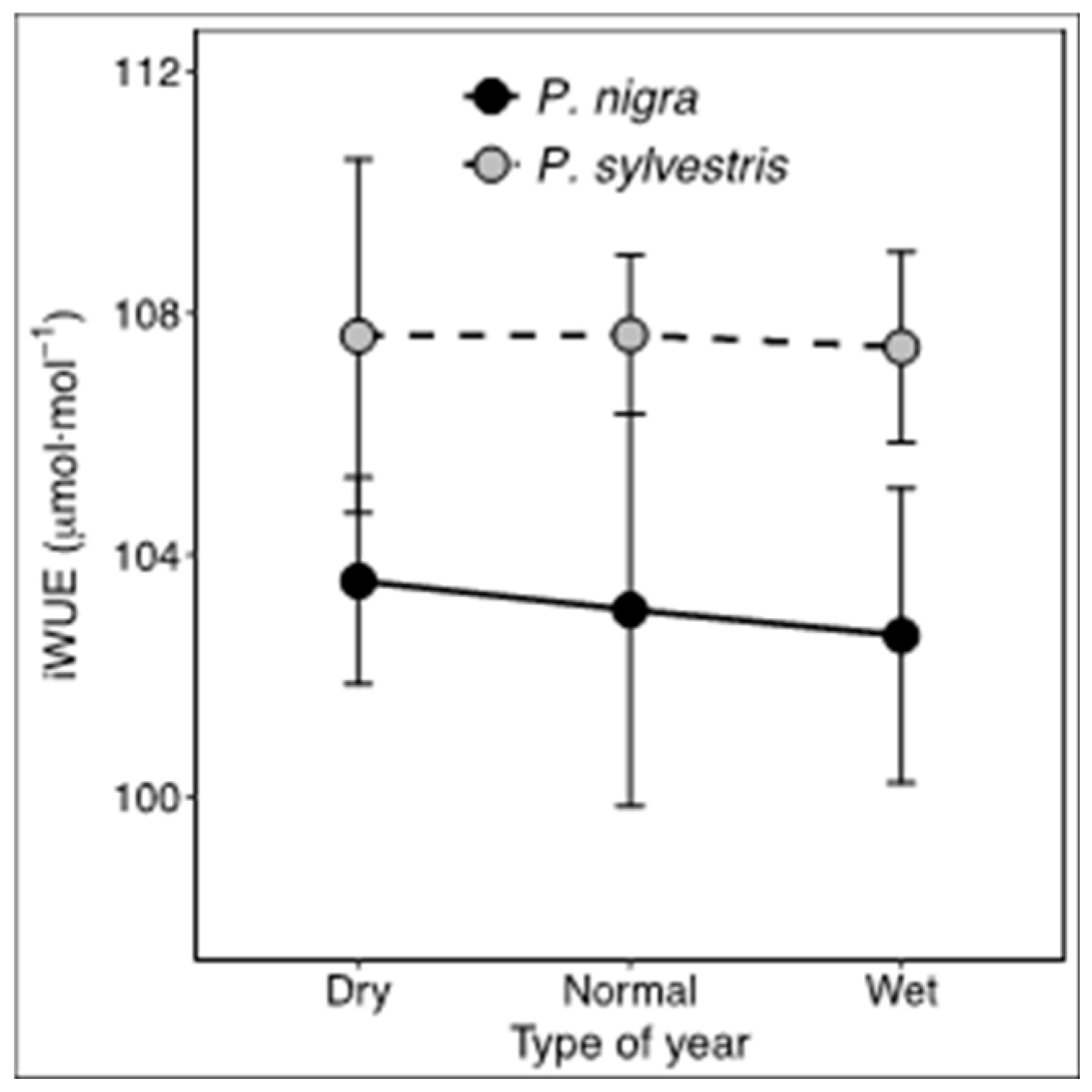
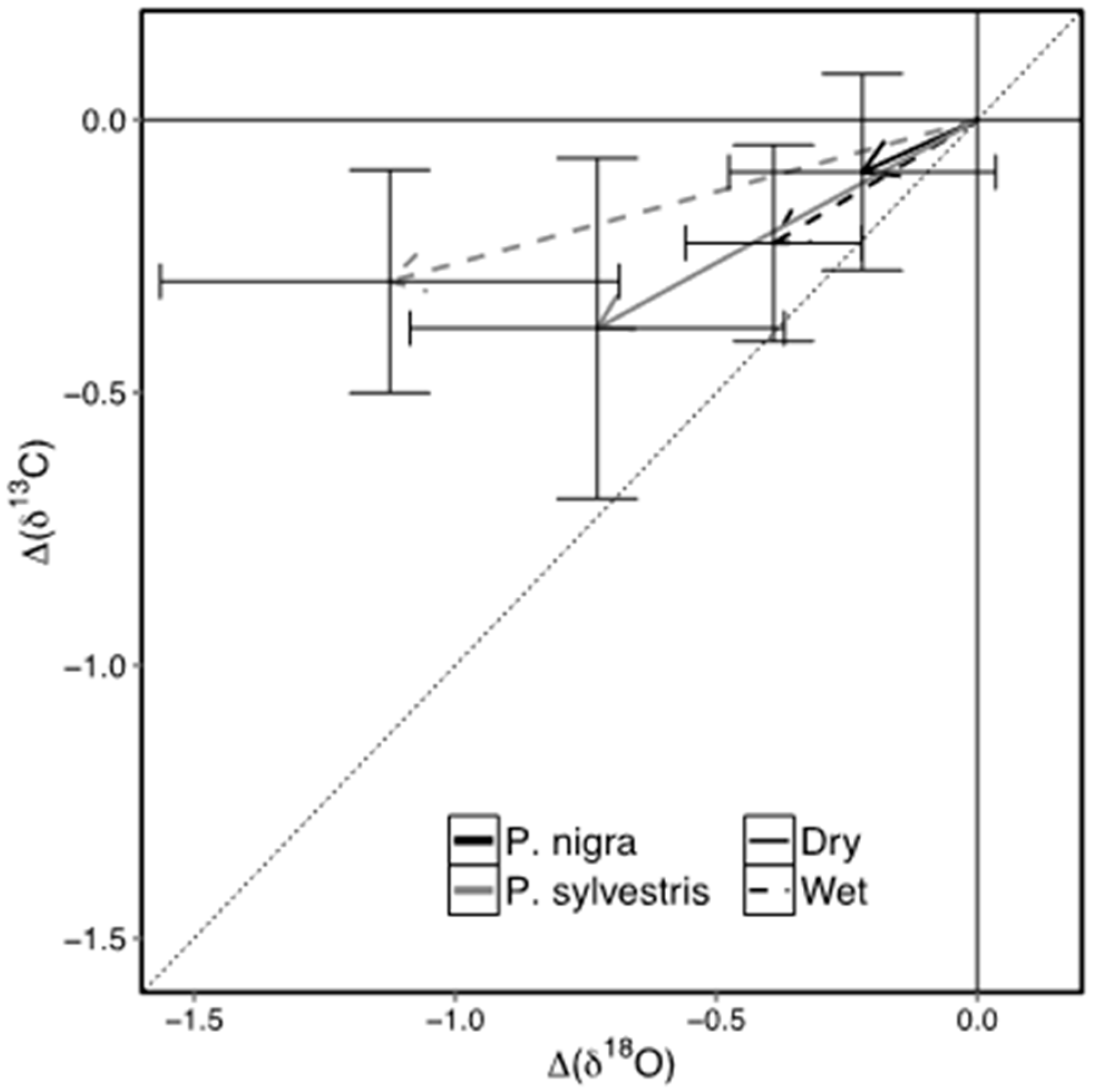
3.4. Stable Isotopes
4. Discussion
4.1. Effects of Drought on Interannual Radial Growth and Xylem Morphology
4.2. Intraannual Responses to Drought
4.3. Interspecific Responses to Drought
4.4. Intrinsic Water-Use Efficiency
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L.D.L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Piñol, J. Drought-induced mortality and hydraulic architecture in pine populations of the NE Iberian Peninsula. For. Ecol. Manag. 2002, 161, 247–256. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; Yepez, E.A. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef]
- Abe, H.; Nakai, T.; Utsumi, Y.; Kagawa, A. Temporal water deficit and wood formation in Cryptomeria japonica. Tree Physiol. 2003, 23, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Paradigm shift in plant growth control. Curr. Opin. Plant Biol. 2015, 25, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Piper, F.; Hoch, G. Physiological mechanisms of drought-induced tree mortality are far from being resolved. New Phytol. 2010, 186, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; Pantin, F.; Génard, M.; Turc, O.; Freixes, S.; Piques, M.; Gibon, Y. Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs. J. Exp. Bot. 2011, 62, 1715–1729. [Google Scholar] [CrossRef] [PubMed]
- Wiley, E.; Helliker, B. A re-evaluation of carbon storage in trees lends greater support for carbon limitation to growth. New Phytol. 2012, 195, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Little, C.H. A.; Savidge, R.A. The role of plant growth regulators in forest tree cambial growth. Plant Growth Regul. 1987, 6, 137–169. [Google Scholar] [CrossRef]
- Dewar, R.C.; Ludlow, A.R.; Dougherty, P.M. Environmental Influences on Carbon Allocation in Pines. Ecol. Bull. 1994, 92–101. [Google Scholar]
- Gruber, A.; Pirkebner, D.; Florian, C.; Oberhuber, W. No evidence for depletion of carbohydrate pools in Scots pine (Pinus sylvestris L.) under drought stress. Plant Biol. 2012, 14, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Sterck, F.J.; Zweifel, R.; Sass-Klaassen, U.; Chowdhury, Q. Persisting soil drought reduces leaf specific conductivity in Scots pine (Pinus sylvestris) and pubescent oak (Quercus pubescens). Tree Physiol. 2008, 28, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Irvine, J.; Perks, M.P.; Magnani, F.; Grace, J. The response of Pinus sylvestris to drought: Stomatal control of transpiration and hydraulic conductance. Tree Physiol. 1998, 18, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Jones, H. Stomatal control of photosynthesis and transpiration. J. Exp. Bot. 1998, 49, 387–398. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Poyatos, R.; Aguadé, D.; Retana, J.; Mencuccini, M. A new look at water transport regulation in plants. New Phytol. 2014, 204, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Pittermann, J.; Sperry, J.S.; Hacke, U.G.; Wheeler, J.K.; Sikkema, E.H. Inter-tracheid pitting and the hydraulic efficiency of conifer wood: The role of tracheid allometry and cavitation protection. Am. J. Bot. 2006, 93, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Cobb, A.R.; Jansen, S. Structure and function of bordered pits: New discoveries and impacts on whole-plant hydraulic function. New Phytol. 2008, 177, 608–626. [Google Scholar] [CrossRef] [PubMed]
- Pittermann, J.; Sperry, J.S.; Wheeler, J.K.; Hacke, U.G.; Sikkema, E.H. Mechanical reinforcement of tracheids compromises the hydraulic efficiency of conifer xylem. Plant Cell Environ. 2006, 29, 1618–1628. [Google Scholar] [CrossRef] [PubMed]
- Olano, J.M.; Linares, J.C.; Garcia-Cervigon, A.I.; Arzac, A.; Delgado, A.; Rozas, V. Drought-induced increase in water-use efficiency reduces secondary tree growth and tracheid wall thickness in a Mediterranean conifer. Oecologia 2014, 176, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.; Hill, R.S. The importance of xylem constraints in the distribution of conifer species. New Phytol. 1999, 143, 365–372. [Google Scholar] [CrossRef]
- Herrero, A.; Castro, J.; Zamora, R.; Delgado-Huertas, A.; Querejeta, J.I. Growth and stable isotope signals associated with drought-related mortality in saplings of two coexisting pine species. Oecologia 2013, 173, 1613–1624. [Google Scholar] [CrossRef] [PubMed]
- Quero, J.L.; Sterck, F.J.; Martínez-Vilalta, J.; Villar, R. Water-use strategies of six co-existing Mediterranean woody species during a summer drought. Oecologia 2011, 166, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Petrucco, L.; Nardini, A.; von Arx, G.; Saurer, M.; Cherubini, P. Isotope signals and anatomical features in tree rings suggest a role for hydraulic strategies in diffuse drought-induced die-back of Pinus nigra. Tree Physiol. 2017, 37, 523–535. [Google Scholar] [PubMed]
- Martinez-Vilalta, J.; Sala, A.; Pinol, J. The hydraulic architecture of Pinaceae—A review. Plant Ecol. 2004, 171, 3–13. [Google Scholar] [CrossRef]
- Barbéro, M.; Losiel, R.; Queézel, P.; Richardson, D.M.; Romane, F. Pines of the Mediterranean Basin. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Nikolov, N.; Helmisaari, H. Silvics of the circumpolar boreal forest tree species. In A Systems Analysis of the Global Boreal Forest; Shugart, H.H., Leemans, R., Bonan, G.B., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 13–84. [Google Scholar]
- Martin-Benito, D.; Beeckman, H.; Cañellas, I. Influence of drought on tree rings and tracheid features of Pinus nigra and Pinus sylvestris in a mesic Mediterranean forest. Eur. J. For. Res. 2013, 132, 33–45. [Google Scholar] [CrossRef]
- Froux, F.; Huc, R.; Ducrey, M.; Dreyer, E. Xylem hydraulic efficiency versus vulnerability in seedlings of four contrasting Mediterranean tree species (Cedrus atlantica, Cupressus sempervirens, Pinus halepensis and Pinus nigra). Ann. For. Sci. 2002, 59, 409–418. [Google Scholar] [CrossRef]
- Meehl, G.A.; Tebaldi, C. More Intense, more frequent, and longer lasting heat waves in the 21st century. Science 2004, 305, 994–997. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon Isotope Discrimination and Photosynthesis. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- Beeckman, H. Wood anatomy and trait-based ecology. IAWA J. 2016, 37, 127–151. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Rinn, F. TSAP−Win Professional, Time Series Analysis and Presentation for Dendrochronology and Related Applications; Version 0.3; Quick Reference; Frank Rinn: Heidelberg, Germany, 2003; p. 20. [Google Scholar]
- Holmes, R.L. Computer−assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Sangüesa-Barreda, G.; Camarero, J.J.; García-Martín, A.; Hernández, R.; de la Riva, J. Remote-sensing and tree-ring based characterization of forest defoliation and growth loss due to the Mediterranean pine processionary moth. For. Ecol. Manag. 2014, 320, 171–181. [Google Scholar] [CrossRef]
- Schmitz, N.; Verheyden, A.; Kairo, J.G.; Beeckman, H.; Koedam, N. Successive cambia development in Avicennia marina (Forssk.) Vierh. is not climatically driven in the seasonal climate at Gazi Bay, Kenya. Dendrochronologia 2007, 25, 87–96. [Google Scholar] [CrossRef]
- Donaldson, L.A. Abnormal lignin distribution in wood from severely drought stressed Pinus radiata trees. IAWA J. 2002, 23, 161–178. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ (1.38x); U.S. National Institutes of Health: Bethesda, MD, USA, 1997.
- Vysotskaya, L.G.; Vaganov, E.A. Components of the variability of radial cell size in tree rings of conifers. Iawa Bull. 1989, 10, 417–426. [Google Scholar] [CrossRef]
- Filion, L.; Cournoyer, L. Variation in wood structure of eastern larch defoliated by the larch sawfly in subarctic Quebec, Can. Can. J. For. Res. 1995, 25, 1263–1268. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Pockman, W.T.; Davis, S.D.; McCulloch, K.A. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 2001, 126, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Vaganov, E.A. The tracheidogram method in tree-ring analysis and its application. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L., Eds.; Kluwer: Dordrecht, The Netherlands, 1989; pp. 63–76. [Google Scholar]
- Loader, N.J.; Robertson, I.; Barker, A.C.; Switsur, V.R.; Waterhouse, J.S. An improved technique for the batch processing of small wholewood samples to α-cellulose. Chem. Geol. 1997, 136, 313–317. [Google Scholar] [CrossRef]
- Wieloch, T.; Helle, G.; Heinrich, I.; Voigt, M.; Schyma, P. A novel device for batch-wise isolation of α-cellulose from small−amount wholewood samples. Dendrochronologia 2011, 29, 115–117. [Google Scholar] [CrossRef]
- Evans, M.N.; Selmer, K.J.; Breeden, B.T.; Lopatka, A.S.; Plummer, R.E. Correction algorithm for on-line continuous flow stable isotope analyses. G3 2016, 17, 3580–3588. [Google Scholar]
- Keeling, C.D. The Suess effect: 13Carbon−14Carbon interrelations. Environ. Int. 1979, 2, 229–300. [Google Scholar] [CrossRef]
- McCarroll, D.; Gagen, M.H.; Loader, N.J.; Robertson, I.; Anchukaitis, K.J.; Los, S.; Young, G.H.F.; Jalkanen, R.; Kirchhefer, A.; Waterhouse, J.S. Correction of tree ring stable carbon isotope chronologies for changes in the carbon dioxide content of the atmosphere. Geochim. Cosmochim. Acta 2009, 73, 1539–1547. [Google Scholar] [CrossRef]
- Scheidegger, Y.; Saurer, M.; Bahn, M.; Siegwolf, R. Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: A conceptual model. Oecologia 2000, 125, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Roden, J.; Siegwolf, R. Is the dual-isotope conceptual model fully operational? Tree Physiol. 2012, 32, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: http://www.r-project.org/ (accessed on 18 January 2016).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-128. Available online: http://CRAN.R-project.org/package=nlme (accessed on 1 December 2016).
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Information theory and log-likelihood models: A basis for model selection and Inference. In Model Selection and Multimodel Inference: A Practical Information−Theoretic Approach; Springer: New York, NY, USA, 2002; pp. 33–74. [Google Scholar]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Fonti, P.; Rigling, A. Drought-induced adaptation of the xylem in Scots pine and pubescent oak. Tree Physiol. 2009, 29, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Graf Pannatier, E.; Rigling, A. Drought alters timing, quantity, and quality of wood formation in Scots pine. J. Exp. Bot. 2011, 62, 2763–2771. [Google Scholar] [CrossRef] [PubMed]
- Fernández-de-Uña, L.; Rossi, S.; Aranda, I.; Fonti, P.; González-González, B.D.; Cañellas, I.; Gea-Izquierdo, G. Xylem and Leaf Functional Adjustments to Drought in Pinus sylvestris and Quercus pyrenaica at Their Elevational Boundary. Front. Plant. Sci. 2017, 8, 1200. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Simard, S.; Rathgeber, C.; Deslauriers, A.; De Zan, C. Effects of a 20-day-long dry period on cambial and apical meristem growth in Abies balsamea seedlings. Trees 2009, 23, 85–93. [Google Scholar] [CrossRef]
- Sperry, J.S.; Hacke, U.G.; Pittermann, J. Size and function in conifer tracheids and angiosperm vessels. Am. J. Bot. 2006, 93, 1490–1500. [Google Scholar] [CrossRef] [PubMed]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap; Springer: New York, NY, USA, 2002. [Google Scholar]
- Esteban, L.; Martín, J.; de Palacios, P.; Fernández, F. Influence of region of provenance and climate factors on wood anatomical traits of Pinus nigra Arn. subsp. salzmannii. Eur. J. For. Res. 2012, 131, 633–645. [Google Scholar] [CrossRef]
- Martín, J.; Esteban, L.; de Palacios, P.; Fernández, F. Variation in wood anatomical traits of Pinus sylvestris L. between Spanish regions of provenance. Trees 2010, 24, 1017–1028. [Google Scholar] [CrossRef]
- Wilkinson, S.; Ogée, J.; Domec, J.-C.; Rayment, M.; Wingate, L. Biophysical modelling of intra-ring variations in tracheid features and wood density of Pinus pinaster trees exposed to seasonal droughts. Tree Physiol. 2017, 35, 305–318. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Saurer, M.; Aronne, G.; Tognetti, R.; Cherubini, P. Variations of wood anatomy and δ13C within-tree rings of coastal Pinus pinaster showing intra-annual density fluctuations. IAWA J. 2007, 28, 61–74. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Lebourgeois, F.; Levy, G.; Aussenac, G.; Clerc, B.; Willm, F. Influence of soil drying on leaf water potential, photoshyntesis, stomatal conductance and growth in two black pine varieties. Ann. For. Sci. 1998, 55, 287–299. [Google Scholar] [CrossRef]
- Voltas, J.; Camarero, J.J.; Carulla, D.; Aguilera, M.; Ortiz, A.; Ferrio, J.P. A retrospective, dual-isotope approach reveals individual predispositions to winter-drought induced tree dieback in the southernmost distribution limit of Scots pine. Plant Cell Environ. 2013, 36, 1435–1448. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, R.; Rosner, S.; Kayler, Z.E.; Andreassen, K.; Børja, I.; Solberg, S.; Tveito, O.E.; Priesack, E.; Gessler, A. Norway spruce physiological and anatomical predisposition to dieback. For. Ecol. Manag. 2014, 322, 27–36. [Google Scholar] [CrossRef]
- Robert, E.M. R.; Koedam, N.; Beeckman, H.; Schmitz, N. A safe hydraulic architecture as wood anatomical explanation for the difference in distribution of the mangroves Avicennia and Rhizophora. Funct. Ecol. 2009, 23, 649–657. [Google Scholar] [CrossRef]
- Zhang, J.-L.; Cao, K.-F. Stem hydraulics mediates leaf water status, carbon gain, nutrient use efficiencies and plant growth rates across dipterocarp species. Funct. Ecol. 2009, 23, 658–667. [Google Scholar] [CrossRef]
- Delucia, E.H.; Maherali, H.; Carey, E.V. Climate-driven changes in biomass allocation in pines. Glob. Chang. Biol. 2000, 6, 587–593. [Google Scholar] [CrossRef]
- McDowell, N.G. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, M.; Siegwolf, R.; Saurer, M.; Eilmann, B.; Rigling, A. Increased water-use efficiency does not lead to enhanced tree growth under xeric and mesic conditions. New Phytol. 2014, 203, 94–109. [Google Scholar] [CrossRef] [PubMed]
- Aguadé, D.; Poyatos, R.; Rosas, T.; Martínez-Vilalta, J. Comparative drought responses of Quercus ilex L. and Pinus sylvestris L. in a montane forest undergoing a vegetation shift. Forests 2015, 6, 2505–2529. [Google Scholar] [CrossRef]
- Galiano, L.; Martínez-Vilalta, J.; Lloret, F. Drought-induced multifactor decline of Scots pine in the Pyrenees and potential vegetation change by the expansion of co-occurring oak species. Ecosystems 2010, 13, 978–991. [Google Scholar] [CrossRef]
- Poyatos, R.; Aguade, D.; Galiano, L.; Mencuccini, M.; Martinez-Vilalta, J. Drought-induced defoliation and long periods of near-zero gas exchange play a key role in accentuating metabolic decline of Scots pine. New Phytol. 2013, 200, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Mencuccini, M.; Bonosi, L. Leaf/sapwood area ratios in Scots pine show acclimation across Europe. Can. J. For. Res. 2001, 31, 442–456. [Google Scholar] [CrossRef]
- Gruber, A.; Strobl, S.; Veit, B.; Oberhuber, W. Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree. Physiol. 2010, 30, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Belmecheri, S.; Maxwell, R.S.; Taylor, A.H.; Davis, K.J.; Freeman, K.H.; Munger, W.J. Tree-ring δ13C tracks flux tower ecosystem productivity estimates in a NE temperate forest. Environ. Res. Lett. 2014, 9, 074011. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (df = 8) | SPEI12 (df = 423) | Year (df = 423) | Species × SPEI12 (df = 423) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | Section | Estimate | SE | p | Estimate | SE | p | Estimate | SE | p | Estimate | SE | p |
| Width | Ring | 47.67 | 87.78 | 0.6019 | 81.13 | 23.26 | 0.0005 | −6.71 | 1.35 | <0.0001 | −35.85 | 29.31 | 0.2220 |
| EW | 51.23 | 66.05 | 0.4602 | 49.27 | 18.75 | 0.0089 | −6.41 | 1.10 | <0.0001 | −21.82 | 23.60 | 0.3558 | |
| LW | −3.63 | 39.49 | 0.9290 | 31.84 | 7.41 | <0.0001 | −0.29 | 0.43 | 0.4924 | −13.98 | 9.33 | 0.1347 | |
| CWT | Ring | −0.35 | 0.26 | 0.2046 | 0.03 | 0.07 | 0.9581 | −0.02 | <0.01 | <0.0001 | −0.06 | 0.10 | 0.4894 |
| EW | −0.56 | 0.26 | 0.0642 | 0.10 | 0.04 | 0.0099 | −0.01 | <0.01 | <0.0001 | −0.07 | 0.05 | 0.1608 | |
| LW | −1.54 | 0.53 | 0.0195 | 0.30 | 0.09 | 0.0009 | <0.01 | <0.01 | 0.4976 | <−0.01 | 0.11 | 0.9985 | |
| LD | Ring | −2.48 | 1.42 | 0.1190 | −0.31 | 0.24 | 0.1984 | 0.06 | 0.01 | <0.0001 | 0.01 | 0.30 | 0.9390 |
| EW | −4.01 | 1.39 | 0.0206 | 0.30 | 0.23 | 0.1964 | 0.01 | 0.01 | 0.3781 | 0.02 | 0.30 | 0.9430 | |
| LW | 0.04 | 0.38 | 0.9122 | 0.34 | 0.11 | 0.0028 | 0.01 | <0.01 | 0.0385 | 0.13 | 0.14 | 0.3672 | |
| Number of Cells | Ring | 4.08 | 2.67 | 0.1653 | 2.23 | 0.64 | 0.0006 | −0.18 | 0.04 | <0.0001 | −0.81 | 0.82 | 0.3193 |
| EW | 3.31 | 1.39 | 0.0185 | 1.02 | 0.43 | 0.0185 | −0.15 | 0.03 | <0.0001 | −0.33 | 0.54 | 0.5411 | |
| LW | 0.77 | 1.74 | 0.6695 | 1.21 | 0.31 | 0.0002 | −0.03 | 0.02 | 0.1561 | −0.48 | 0.40 | 0.2280 | |
| P. nigra | P. sylvestris | ||
|---|---|---|---|
| Cell-wall thickness 1 | EW | 0.0136 | 0.0612 |
| LW | 0.0004 | 0.0123 | |
| Total | 0.0140 | 0.0735 | |
| Lumen diameter 1 | EW | 2.0721 | 3.5085 |
| LW | 5.5612 | 6.0029 | |
| Total | 7.6333 | 9.5113 | |
| Number of cells 1 | EW | 89.4257 | 86.7695 |
| LW | 2.1780 | 2.5628 | |
| Total | 91.6037 | 89.3323 | |
| Tree 2 | 0.0654 | 0.0702 | |
| Residuals | 0.6837 | 1.0127 | |
| Dry (n = 8) | Average (n = 29) | Wet (n = 10) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pinus nigra | ||||||||||
| Width (10−2 mm) | Ring | 555.89 | (263.33) | a | 727.56 | (280.45) | b | 948.92 | (409.93) | c |
| EW | 453.82 | (201.88) | a | 588.27 | (219.40 | b | 749.64 | (321.74) | c | |
| LW | 102.07 | (80.43) | a * | 139.28 | (91.54) | b | 199.28 | (128.00) | c | |
| CWT (μm) | Ring | 3.86 | (1.15) | a | 4.16 | (1.12) | ab | 4.46 | (1.02) | a |
| EW | 3.30 | (0.67) | a | 3.79 | (0.66) | b | 3.97 | (0.52) | c | |
| LW | 5.50 | (1.81) | a | 6.05 | (1.46) | a | 6.25 | (1.58) | a | |
| LD (μm) | Ring | 35.93 | (4.06) | a | 34.19 | (3.50) | b | 33.40 | (3.76) | b |
| EW | 34.26 | (4.43) | a | 34.84 | (3.24) | a | 34.46 | (2.73) | a | |
| LW | 7.20 | (1.95) | ab | 7.14 | (1.50) | a | 7.80 | (1.60) | b | |
| Number of Cells | Ring | 16.36 | (6.79) | a | 20.91 | (7.42) | b | 26.94 | (10.73) | c |
| EW | 11.09 | (4.33) | a | 13.93 | (4.82) | b | 17.64 | (6.82) | c | |
| LW | 5.27 | (3.19) | a | 6.98 | (3.64) | b | 9.29 | (5.17) | c | |
| Pinus sylvestris | ||||||||||
| Width (mm) | Ring | 733.38 | (337.88) | ab | 759.36 | (334.55) | a | 985.78 | (441.26) | b |
| EW | 605.38 | (267.78) | a | 621.49 | (265.14) | a | 807.56 | (383.84) | b | |
| LW | 127.99 | (126.21) | ab | 137.87 | (122.07) | a | 178.22 | (126.44) | b* | |
| CWT (μm) | Ring | 3.58 | (1.07) | a | 3.88 | (0.99) | a | 3.97 | (1.05) | a |
| EW | 3.03 | (0.73) | a | 3.17 | (0.71) | ab | 3.40 | (0.77) | b* | |
| LW | 4.15 | (1.40) | a | 4.54 | (1.35) | a | 4.74 | (1.16) | a | |
| LD (μm) | Ring | 32.91 | (4.63) | a * | 31.72 | (3.71) | ab | 30.77 | (3.71) | b* |
| EW | 30.28 | (5.01) | a | 30.73 | (3.68) | a | 30.60 | (3.70) | a | |
| LW | 7.29 | (1.94) | a | 7.14 | (1.48) | a | 8.15 | (1.64) | b* | |
| Number of Cells | Ring | 23.75 | (9.97) | a | 24.54 | (10.19) | a | 31.07 | (12.36) | b |
| EW | 16.63 | (6.56) | a | 16.74 | (6.18) | a | 21.50 | (9.01) | b | |
| LW | 7.12 | (5.59) | ab | 7.81 | (5.59) | a | 9.56 | (5.67) | b | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Benito, D.; Anchukaitis, K.J.; Evans, M.N.; Del Río, M.; Beeckman, H.; Cañellas, I. Effects of Drought on Xylem Anatomy and Water-Use Efficiency of Two Co-Occurring Pine Species. Forests 2017, 8, 332. https://doi.org/10.3390/f8090332
Martin-Benito D, Anchukaitis KJ, Evans MN, Del Río M, Beeckman H, Cañellas I. Effects of Drought on Xylem Anatomy and Water-Use Efficiency of Two Co-Occurring Pine Species. Forests. 2017; 8(9):332. https://doi.org/10.3390/f8090332
Chicago/Turabian StyleMartin-Benito, Dario, Kevin J. Anchukaitis, Michael N. Evans, Miren Del Río, Hans Beeckman, and Isabel Cañellas. 2017. "Effects of Drought on Xylem Anatomy and Water-Use Efficiency of Two Co-Occurring Pine Species" Forests 8, no. 9: 332. https://doi.org/10.3390/f8090332
APA StyleMartin-Benito, D., Anchukaitis, K. J., Evans, M. N., Del Río, M., Beeckman, H., & Cañellas, I. (2017). Effects of Drought on Xylem Anatomy and Water-Use Efficiency of Two Co-Occurring Pine Species. Forests, 8(9), 332. https://doi.org/10.3390/f8090332