Converting Larch Plantations to Larch-Walnut Mixed Stands: Effects of Spatial Distribution Pattern of Larch Plantations on the Rodent-Mediated Seed Dispersal of Juglans mandshurica

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
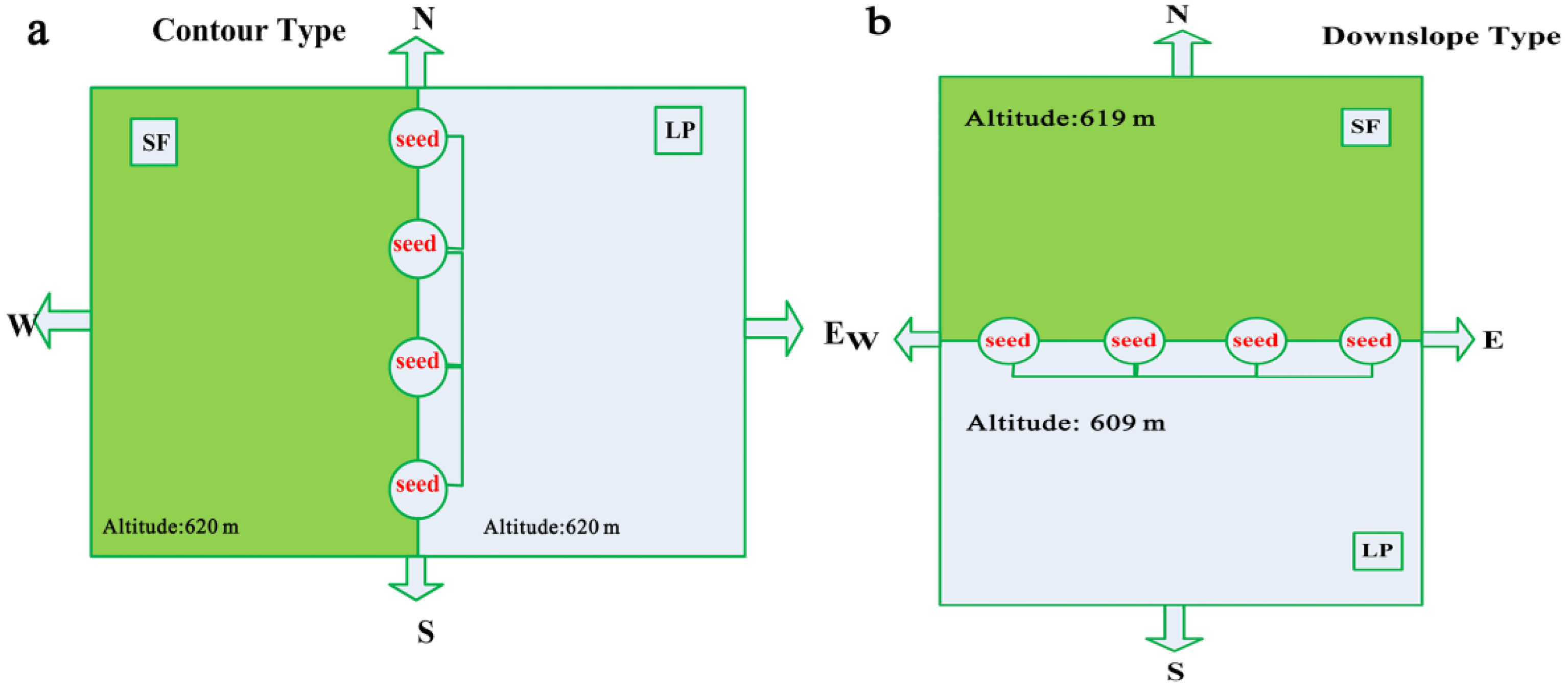
2.2. Seed Collection and Marking
2.3. Seed Release and Seed Removal
2.4. Identification of Seed Disperser
2.5. Statistical Analysis
3. Results
3.1. Identification of Seed Dispersers
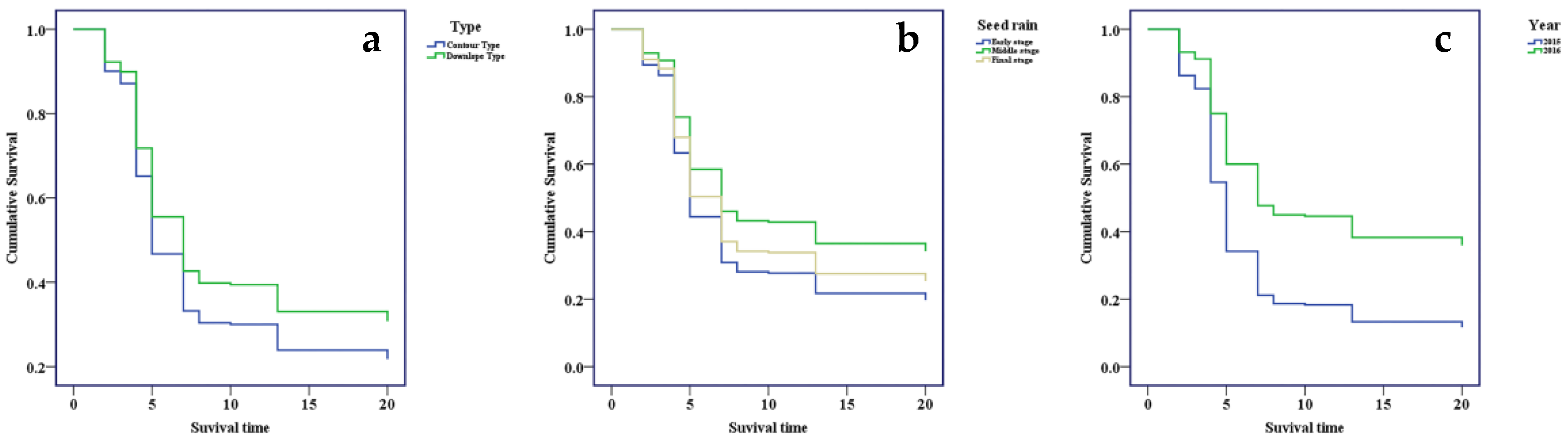
3.2. Seed Dispersal Rate
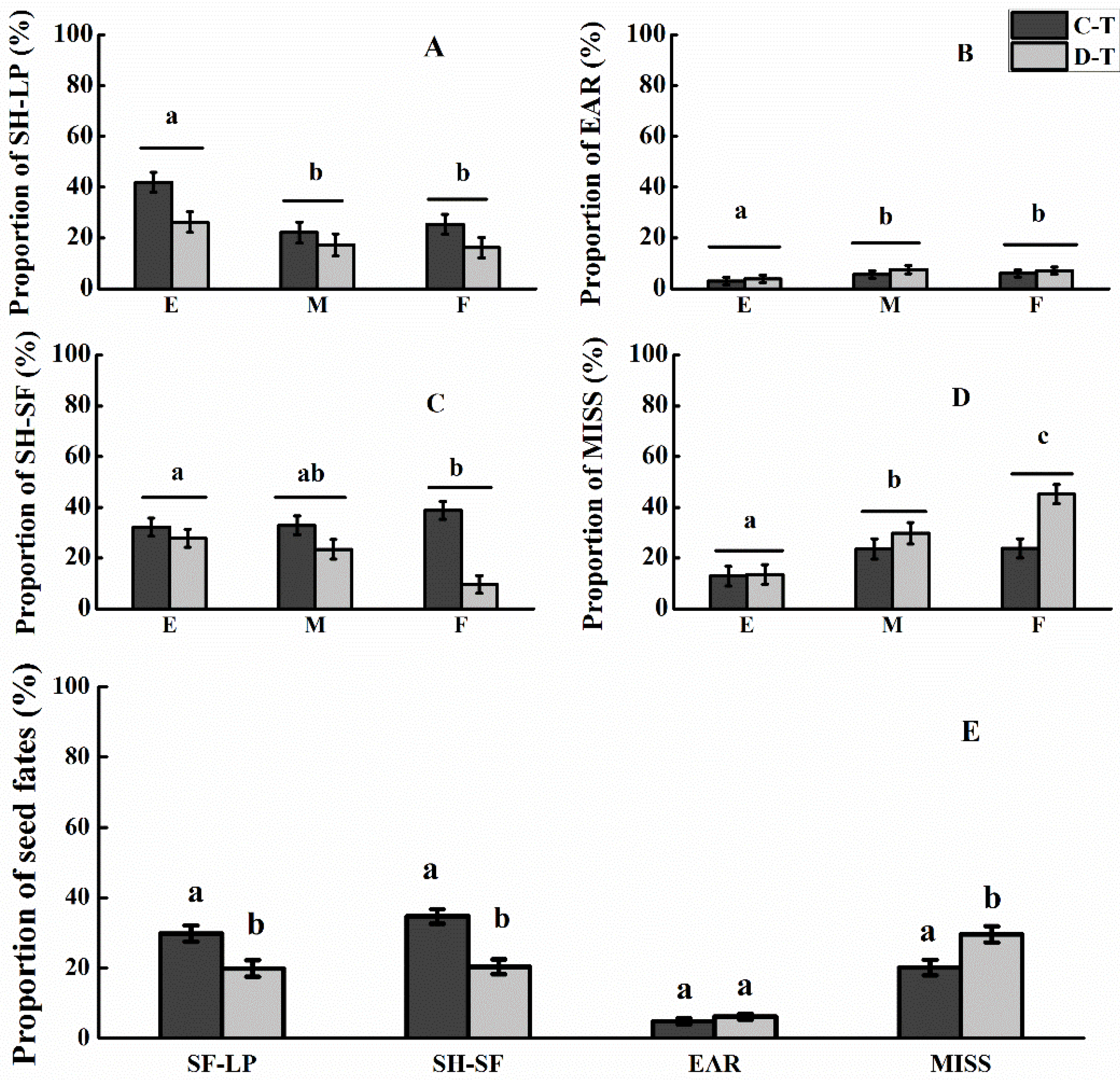
3.3. Seed Fates
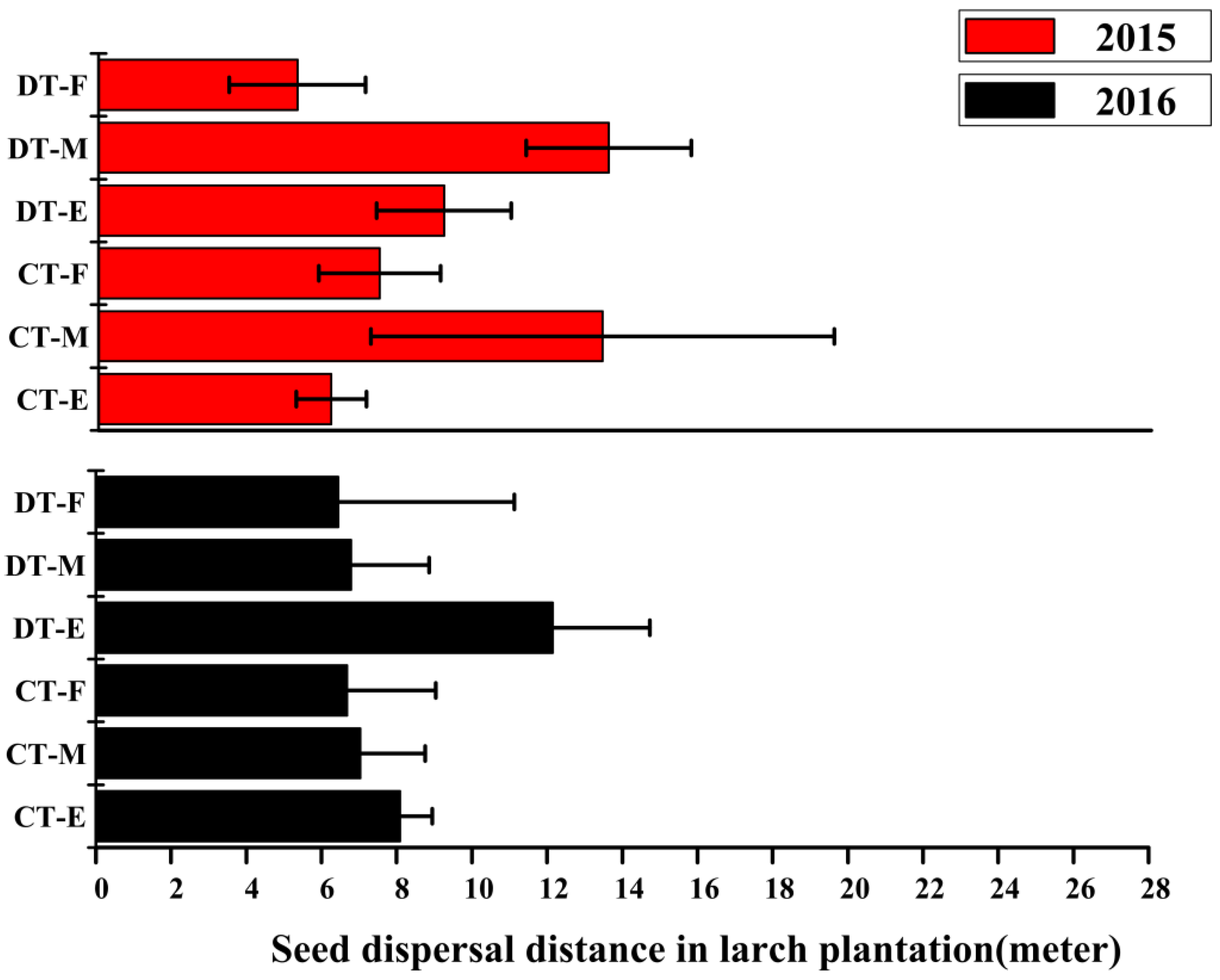
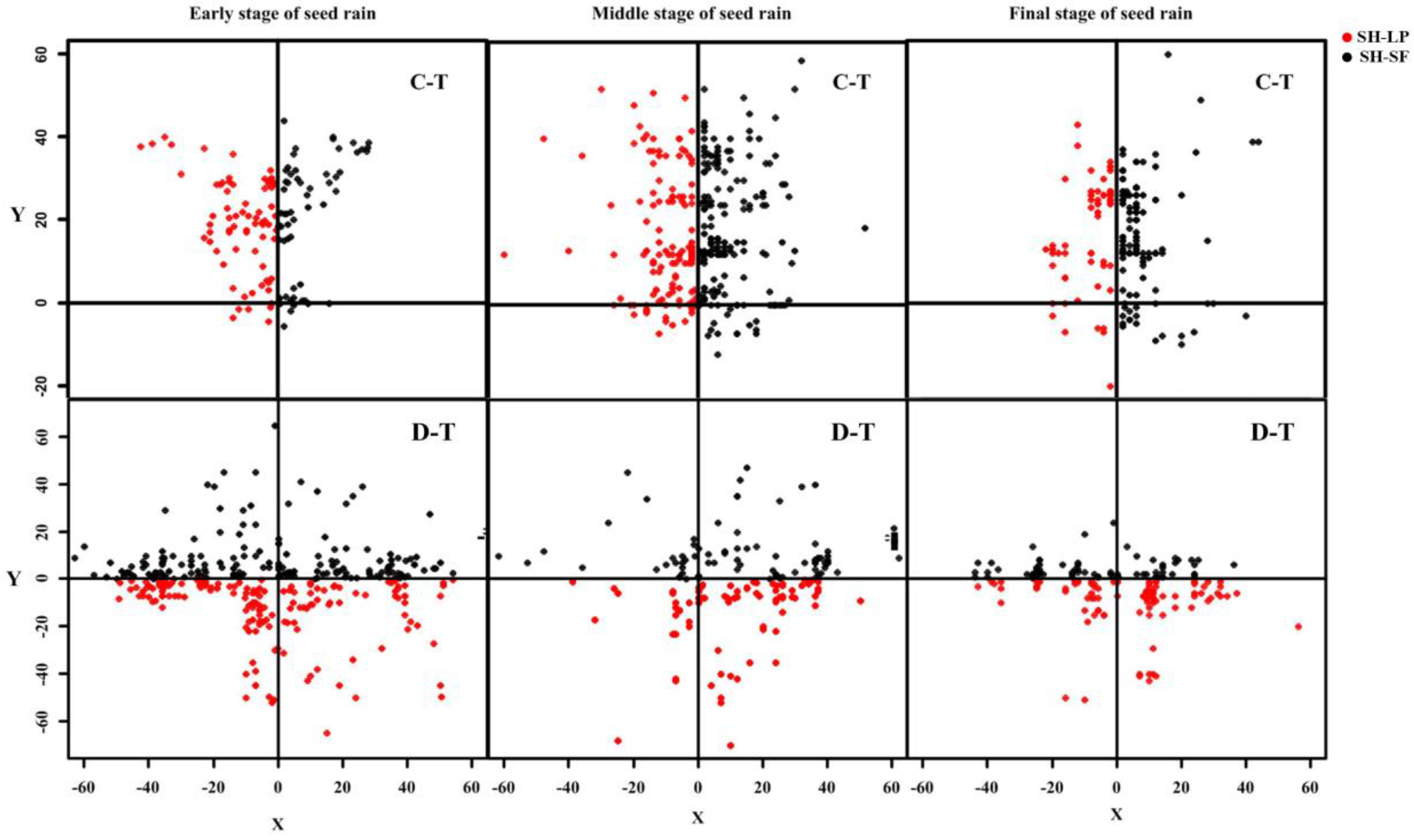
3.4. Seed Dispersal Distance
4. Discussions
4.1. Effects of Spatial Allocation Type of LP and Adjacent SF on Rodent-Mediated Seed Dispersal
4.2. Effects of the Inter-Annual Variation of Seed Production and Different Stages of Seed Rain on Rodent-Mediated Seed Dispersal
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | Explanatory Variables Used in Model | Likehoods | AIC | ∆AIC |
|---|---|---|---|---|
| SH-LP | Null model | −45.7 | −47.70 | 8.55 |
| SH-LP | allocation | −52.7 | −50.70 | 5.55 |
| SH-LP | allocation + year | −48.63 | −46.63 | 9.62 |
| SH-LP | allocation + year + seed rain + nests (station/site) | −58.25 | −54.25 | 6.55 |
| SH-LP | allocation + year + seed rain | −58.25 | −56.25 | 0 |
| SH-SF | Null model | 74.04 | 72.04 | 155.77 |
| SH-SF | allocation + year | −78.37 | −78.37 | 160.1 |
| SH-SF | allocation + year + seed rain | −72.86 | −70.86 | 154.53 |
| SH-SF | allocation + year + seed rain + nests (station/site) | −72.86 | −68.86 | 154.59 |
| SH-SF | allocation | −81.73 | −83.73 | 0 |
| MISS | Null model | 77.13 | 79.13 | 106.62 |
| MISS | allocation | 12.99 | 14.99 | 42.48 |
| MISS | allocation + year | −24.82 | −22.82 | 4.67 |
| MISS | allocation + year + seed rain + nests (station/site) | −31.49 | −29.49 | 2 |
| MISS | allocation + year + seed rain | −34.49 | −27.49 | 0 |
| IS | Null model | 43.11 | 45.11 | 51.3 |
| IS | allocation | 4.12 | 6.12 | 12.31 |
| IS | allocation + year + seed rain | −0.79 | 3.21 | 9.4 |
| IS | allocation + year + seed rain + nests (station/site) | −0.79 | 1.2 | 7.40 |
| IS | allocation + year | −8.19 | −6.19 | 0 |
| EAR | Null model | −265.63 | −263.63 | 47.44 |
| EAR | allocation + year | −302.73 | −298.73 | 12.34 |
| EAR | allocation + year + seed rain | −309.62 | −307.59 | 7.66 |
| EAR | allocation + year + seed rain + nests (station/site) | −302.73 | −300.73 | 10.34 |
| EAR | allocation | −313.07 | −311.07 | 0 |
| Distance | Null model | 311.98 | 311.02 | 215.29 |
| Distance | allocation | 91.09 | 97.23 | 1.5 |
| Distance | allocation + year | 90.79 | 99.15 | 3.42 |
| Distance | allocation + year + seed rain + nests (station/site) | 102.11 | 108.32 | 12.59 |
| Distance | allocation + year + seed rain | 82.98 | 95.73 | 0 |
References
- Zhu, J.J.; Mao, Z.H.; Hu, L.L.; Zhang, J.Z. Plant diversity of secondary forests in response to anthropogenic disturbance levels in montane regions of northeastern China. J. For. Res. 2007, 12, 403–416. [Google Scholar] [CrossRef]
- Chen, X.W.; Li, B.L.; Li, Z.S. The acceleration of succession for the restoration of the mixed-broadleaved Korean pine forests in Northeast China. For. Ecol. Manag. 2003, 117, 503–514. [Google Scholar] [CrossRef]
- Yan, Q.L.; Zhu, J.J.; Yu, L.Z. Seed regeneration potential of canopy gaps at early formation stage in temperate secondary forests, Northeast China. PLoS ONE 2012, 7, e39502. [Google Scholar] [CrossRef] [PubMed]
- Mason, W.L.; Zhu, J.J. Silviculture of planted forests managed for multi-functional objectives: Lessons from Chinese and British experience. In Challenge and Opportunity for the World’s Forests in the 21st Century; Fenning, T., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 37–54. [Google Scholar]
- Yang, K.; Zhu, J.J.; Zhang, M.; Yan, Q.L.; Sun, O.J. Soil microbial biomass carbon and nitrogen in forest ecosystems of Northeast China: A comparison between natural secondary forest and Larch plantation. J. Plant Ecol. 2010, 3, 175–182. [Google Scholar] [CrossRef]
- Yang, K.; Shi, W.; Zhu, J.J. The impact of secondary forests conversion into larch plantations on soil chemical and microbiological properties. Plant Soil 2013, 368, 535–546. [Google Scholar] [CrossRef]
- Zhu, J.J.; Yang, K.; Yan, Q.L.; Liu, Z.G.; Wang, H.X. The feasibility of implementing thinning in pure even-aged Larix olgensis plantations to establish uneven aged larch-broadleaved mixed forests. J. For. Res. 2010, 15, 70–81. [Google Scholar] [CrossRef]
- Zhu, J.J.; Liu, Z.G.; Wang, H.X.; Yan, Q.L.; Fang, H.Y.; Hu, L.L.; Yu, L.Z. Effects of site preparations on emergence and early establishment of Larix olgensis in montane regions of northeastern China. New For. 2008, 36, 247–260. [Google Scholar] [CrossRef]
- Kolo, H.; Ankerst, D.; Knoke, T. Predicting natural forest regeneration: A statistical model based on inventory data. Eur. J. For. Res. 2017, 136, 923–938. [Google Scholar] [CrossRef]
- Wang, B.C.; Smith, T.B. Closing the seed dispersal loop. Trends Ecol. Evol. 2002, 17, 379–385. [Google Scholar] [CrossRef]
- Nomuraa, N.; Tsuyuzaki, S. Hares promote seed dispersal and seedling establishment after volcanic eruptions. Acta Oecol. 2015, 63, 22–27. [Google Scholar] [CrossRef]
- Manzano, P.; Malo, J.E. Extreme long-distance seed dispersal via sheep. Front Ecol. Environ. 2006, 4, 244–248. [Google Scholar] [CrossRef]
- Yan, Q.L.; Zhu, J.J.; Gang, Q. Comparison of spatial patterns of soil seed banks between larch plantations and adjacent secondary forests in Northeast China: Implication for spatial distribution mode of larch plantations. Trees 2013, 27, 1747–1754. [Google Scholar] [CrossRef]
- Vander Wall, S.B.; Longland, W.S. Diplochory: Are two seed dispersers better than one? Trends Evol. Evol. 2004, 19, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Ortega, M.; Bartimachi, A.; Neves, J.; Bruna, E.M.; Vasconcelos, H.L. Seed removal patterns of pioneer trees in an agricultural Landscape. Plant Ecol. 2017, 218, 737–748. [Google Scholar] [CrossRef]
- Miyaki, M. Seed dispersal of the Korean pine; Pinus koraiensis by the red squirrel; Sciurus vulgaris. Ecol. Res. 1987, 2, 147–157. [Google Scholar] [CrossRef]
- Litch, I.N.; Steele, M.A.; Swihair, R.K. Seed fate and decision-making processes in scatter-hoarding rodents. Biol. Rev. 2017, 92, 474–504. [Google Scholar]
- Pesendorfer, M.B.; Sillett, T.S.; Koenig, W.D.; Morrison, S.A. Scatter-hoarding corvids as seed dispersers for oaks and pines: A review of a widely distributed mutualism and its utility to habitat restoration. Condor 2016, 118, 215–237. [Google Scholar] [CrossRef]
- Bello, C.; Galetti, M.; Pizo, M.A.; Magnago, L.F.S.; Rocha, M.F.; Lima, R.A.F.; Jordano, P. Defaunation affects carbon storage in tropical forests. Science 2015, 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mokany, K.; Prasad, S.; Westcott, D.A. Loss of frugivore seed dispersal services under climate change. Nat. Commun. 2014, 5, 3971. [Google Scholar] [CrossRef] [PubMed]
- Mokany, K.; Prasad, S.; Westcott, D.A. Impacts of climate change and management responses in tropical forests depend on complex frugivore-mediated seed dispersal. Glob. Ecol. Biogeogr. 2015, 24, 685–694. [Google Scholar] [CrossRef]
- Melo, F.P.L.; Dirzo, R.; Tabarelli, M. Biased seed rain in forest edges: Evidence from the Brazilian Atlantic forest. Biol. Conserv. 2006, 132, 50–60. [Google Scholar] [CrossRef]
- Xiao, Z.S.; Zhang, Z.B. Contrasting patterns of short-term indirect seed–seed interactions mediated by scatter-hoarding rodents. J. Anim. Ecol. 2016, 85, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Bogdziewicz, M.; Lichti, N.I.; Zwolak, R. Consumer-mediated indirect interaction with a native plant lowers the fitness of an invasive competitor. J. Ecol. 2018. [Google Scholar] [CrossRef]
- Dirzo, R.; Mendoza, E.; Ortiz, P. Size-related differential seed predation in a heavily defaunated neotropical rain forest. Biotropica 2007, 39, 355–362. [Google Scholar] [CrossRef]
- Wang, J.; Yan, Q.L.; Yan, T.; Song, Y.; Sun, Y.R.; Zhu, J.J. Rodent-mediated seed dispersal of Juglans mandshurica regulated by gap size and within-gap position in larch plantations: Implication for converting pure larch plantations into larch-walnut mixed forests. For. Ecol. Manag. 2017, 404, 205–213. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Zhang, M.M.; Yi, X.F. Small rodents trading off forest gaps for scatter-hoarding differs between seed species. For. Ecol. Manag. 2016, 379, 226–231. [Google Scholar] [CrossRef]
- Yan, Q.L.; Zhu, J.J.; Gang, Q.; Huang, L.Y.; Wang, G.G. Comparison of spatial distribution patterns of seed rain between larch plantations and adjacent secondary forests in Northeast China. For. Sci. 2016, 62, 652–662. [Google Scholar] [CrossRef]
- Dalling, J.W.; Muller-landau, H.C.; Wright, S.J.; Hubbell, S.P. Role of dispersal in the recruitment limitation of neotropical pioneer species. J. Ecol. 2002, 90, 714–727. [Google Scholar] [CrossRef]
- Slavík, O.; Horky, P.; Maciak, M.; Horká, P.; Langrová, I. Diel movement of brown trout, Salmotrutta, is reduced in dense populations with high site fidelity. Ecol. Evol. 2018, 8, 4495–4507. [Google Scholar] [CrossRef] [PubMed]
- Gomes, V.S.D.M.; Tamashiro, J.Y.; Silva, W.R. Seed inflow to a forest patch promoted by understory frugivorous birds. Biota Neotrop. 2011, 11, 95–102. [Google Scholar] [CrossRef]
- Xiao, Z.S.; Zhang, Z.B.; Krebs, C.J. Long-term seed survival and dispersal dynamics in a rodent-dispersed tree: Testing the predator satiation hypothesis and the predator dispersal hypothesis. J. Ecol. 2013, 101, 1256–1264. [Google Scholar] [CrossRef]
- Bogdziewicz, M.; Zwolak, R.; Crone, E.E. How do vertebrates respond to mast seeding? Oikos 2016, 125, 300–307. [Google Scholar] [CrossRef]
- Aliyu, B.; Thia, J.A.; Moltchanova, E.; Forget, M.P.; Chapman, H.M. Forest disturbance and seasonal food availability influence a conditional seed dispersal mutualism. Biotropica 2018, 50, 750–757. [Google Scholar] [CrossRef]
- Yi, X.F.; Zhang, Z.B. Influence of insect-infested cotyledons on early seedling growth of Mongolian oak, Quercus mongolica. Photosynthetica 2008, 46, 139–142. [Google Scholar] [CrossRef]
- Yi, X.F.; Wang, Z.Y.; Liu, C.; Liu, G.; Zhang, M.M. Acorn cotyledons are larger than their seedlings’ need: Evidence from artificial cutting experiments. Sci. Rep. 2015, 5, 8112. [Google Scholar] [CrossRef] [PubMed]
- Piao, Z.; Tang, L.; Swihart, R.K.; Wang, S. Human–wildlife competition for Korean pine seeds: Vertebrate responses and implications for mixed forests on Changbai Mountain, China. Ann. For. Sci. 2011, 68, 911–919. [Google Scholar] [CrossRef]
- Tamura, N.; Hayashi, F. Geographic variation in walnut seed size correlates with hoarding behaviours of two rodent species. Ecol. Res. 2008, 23, 607–614. [Google Scholar] [CrossRef]
- Hao, H.; Jiang, C.; Wu, J.P.; Zong, C.; Zeng, D.; Alimubike, D. The effects of scatter-hoarding rodents on Mongolian oak (Quercus mongolica) acorn dispersal and seedling distribution in urban woodland. Urban Ecosyst. 2017, 20, 1277–1283. [Google Scholar] [CrossRef]
- Shi, F.Y.; Chen, X.Q.; Chen, N.Q. Study on the artificial mixed forest of Juglans mandshurica and Larixolgensis. J. Northeast For. Univ. 1991, 51, 32–44. (In Chinese) [Google Scholar]
- Yang, L.X.; Wang, P.; Kong, C. Effect of larch (Larixg melini Rupr.) root exudates on Manchurian walnut (Juglans mandshurica Maxim.) growth and soil juglone in a mixed-species plantation. Plant Soil 2010, 329, 249–258. [Google Scholar] [CrossRef]
- Wang, F.; Zhu, W.X.; Zoo, B.; Deborah, A.; Neher, D.A.; Fu, S.L.; Xia, H.P.; Li, Z.A. Seedling growth and soil nutrient availability inexotic and native tree species: Implications for afforestation in southern China. Plant Soil 2013, 364, 207–218. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.J. Impact of tree litter decomposition on soil biochemical properties obtained from a temperate secondary forest in Northeast China. J. Soil Sediment. 2015, 15, 13–23. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, J.; Sichilima, A.M.; Wang, W.; Lu, J. Effects of thinning on scatter-hoarding by rodents in temperate forest. Integr. Zool. 2016, 11, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chu, W.; Zhang, Z. Cultivated walnut trees showed earlier but not final advantage over its wild relatives in competing for seed dispersers. Integr. Zool. 2017, 12, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.B.; Wang, F.S. Effect of rodents on seed dispersal and survival of wild apricot (Prunus armeniaca). Acta Ecol. Sin. 2001, 21, 839–845. (In Chinese) [Google Scholar]
- Li, H.J.; Zhang, Z.B. Effect of rodents on acorn dispersal and survival of the Liaodong oak (Quercus liaotungensis Koidz.). For. Ecol. Manag. 2003, 176, 387–396. [Google Scholar] [CrossRef]
- Caballero, I.; Olano, J.M.; Loidi, J.; Escudero, A. Seed bank structure along a semi-arid gypsum gradient in Central Spain. J. Arid Environ. 2003, 55, 287–299. [Google Scholar] [CrossRef]
- Yang, C.S. Research on Species Coexiestence of Five Kinds of Rodents in Forestry Area in Northeast. Ph.D. Thesis, Northest Forestry University, Harbin, China, 2007. (In Chinese). [Google Scholar]
- Levey, D.J.; Byrne, M.M. Complex ant-plant interactions: Rainforest ants as secondary dispersers and post-dispersal seed predators. Ecology 1993, 74, 1802–1812. [Google Scholar] [CrossRef]
- Wang, W.; Ma, K.P. Predation and dispersal of Quercus liaotungensis Koidz. acorns by animals in dangling mountain, northern China I: Effect of rodents removal on loss of acorns. Acta Ecol. Sin. 2001, 21, 204–210. (In Chinese) [Google Scholar]
- Zwolak, R.; Bogdziewicz, M.; Wróbel, A.; Crone, E.E. Advantages of masting in European beech: Timing of granivore satiation and benefits of seed caching support the predator dispersal hypothesis. Oecologia 2016, 180, 749–758. [Google Scholar] [CrossRef] [PubMed]




| Spatial Allocation Pattern of Larch Plantations | Center Coordinates | Altitude (m) | Area (ha) | Slope (°) | Gradien (°) | Mean DBH (cm) | Mean Height (m) | Stand Density (tree/ha) | Distance to Closest Site (km) |
|---|---|---|---|---|---|---|---|---|---|
| Downslope type | N 41°51′15″, E 124°56′39″ | 625 | 0.25 | 186 | <15 | 38.1 | 20 | 766 ± 115 | Closest site |
| N 41°50′43″, E 124°56′48″ | 620 | 0.25 | 248 | <15 | 37.2 | 25 | 1233 ± 450 | 1.8 | |
| N 41°50′57″, E 124°56′45″ | 613 | 0.25 | 213 | <15 | 39.2 | 23 | 1160 ± 289 | 2.7 | |
| Contour type | N 41°50′81″, E 124°56′57″ | 644 | 0.25 | 330 | <15 | 38.1 | 19 | 1466 ± 231 | 1.6 |
| N 41°51′56″, E 124°56′70″ | 603 | 0.25 | 340 | <15 | 43.4 | 24 | 1033 ± 208 | 1.7 | |
| N 41°50′57″, E 124°55′55″ | 615 | 0.25 | 335 | <15 | 40.5 | 24 | 1566 ± 289 | 2.3 |
| Spatial Allocation Pattern of Larch Plantations | Rodent | Residence Time at Three Stages of Seed Rain (minutes) | Frequency at Three Stages of Seed Rain (time/10 days) | ||||
|---|---|---|---|---|---|---|---|
| Early Stage | Middle Stage | Final Stage | Early Stage | Middle Stage | Final Stage | ||
| Contour type | Sciurus vulgaris | 10 | 1 | 2 | 3 | 2 | 2 |
| Downslope type | Sciurus vulgaris | 1 | 2 | 2 | 1 | 1 | 1 |
| Response Variable | Spatial Allocation Model | Different Stage of Seed Rain | Inter-Annual Variation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F-Ratio | p-Value | df | F-Ratio | p-Value | df | F-Ratio | p-Value | |
| Seed removal rate | 1 | 15.72 | <0.001 | 2 | 27.9 | 0.034 | 1 | 120.4 | <0.001 |
| IS proportion | 1 | 0.41 | 0.53 | 2 | 0.95 | 0.39 | 1 | 22.21 | 0.001 |
| EAR proportion | 1 | 0.53 | 0.47 | 2 | 3.53 | 0.03 | 1 | 0.19 | 0.66 |
| SH-SF proportion | 1 | 8.37 | 0.62 | 2 | 0.47 | 0.004 | 1 | 0.69 | 0.69 |
| SH-LP proportion | 1 | 0.07 | 0.008 | 2 | 7.29 | 0.001 | 1 | 6.98 | 0.79 |
| MISS proportion | 1 | 0.11 | 0.75 | 2 | 8.71 | <0.001 | 1 | 51.03 | <0.001 |
| Seed dispersal distance | 1 | 0.41 | 0.52 | 2 | 0.54 | 0.59 | 1 | 0.37 | 0.99 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Yan, Q.; Zhang, T.; Lu, D.; Xie, J.; Sun, Y.; Zhang, J.; Zhu, J. Converting Larch Plantations to Larch-Walnut Mixed Stands: Effects of Spatial Distribution Pattern of Larch Plantations on the Rodent-Mediated Seed Dispersal of Juglans mandshurica. Forests 2018, 9, 716. https://doi.org/10.3390/f9110716
Wang J, Yan Q, Zhang T, Lu D, Xie J, Sun Y, Zhang J, Zhu J. Converting Larch Plantations to Larch-Walnut Mixed Stands: Effects of Spatial Distribution Pattern of Larch Plantations on the Rodent-Mediated Seed Dispersal of Juglans mandshurica. Forests. 2018; 9(11):716. https://doi.org/10.3390/f9110716
Chicago/Turabian StyleWang, Jing, Qiaoling Yan, Ting Zhang, Deliang Lu, Jin Xie, Yirong Sun, Jinxin Zhang, and Jiaojun Zhu. 2018. "Converting Larch Plantations to Larch-Walnut Mixed Stands: Effects of Spatial Distribution Pattern of Larch Plantations on the Rodent-Mediated Seed Dispersal of Juglans mandshurica" Forests 9, no. 11: 716. https://doi.org/10.3390/f9110716
APA StyleWang, J., Yan, Q., Zhang, T., Lu, D., Xie, J., Sun, Y., Zhang, J., & Zhu, J. (2018). Converting Larch Plantations to Larch-Walnut Mixed Stands: Effects of Spatial Distribution Pattern of Larch Plantations on the Rodent-Mediated Seed Dispersal of Juglans mandshurica. Forests, 9(11), 716. https://doi.org/10.3390/f9110716