Abstract
Douglas fir (Pseudotsuga menziesii var. glauca (Mayr) Franco), and western larch (Larix occidentalis Nutt.) are species of ecological and commercial importance that occur throughout the Western United States. Effective reforestation of these species relies on successful seedling establishment, which is affected by planting stock quality, stocktype size, and site preparation techniques. This study examined the effects of container volume (80, 130, 200, and 250 cm3) and vegetative competition on seedling survival and physiological and morphological responses for two years, post-outplanting. Glyphosate application (GS) and grass planting (HC) were used to achieve low and high levels of competition. For all measured attributes, the container volume × vegetative competition was not significant. Mortality was strongly influenced by competition, with higher mortality observed for Douglas fir and western larch planted in HC plots one (28% and 98%) and two (61% and 99%) years following outplanting. When competition was controlled, seedlings of both species exhibited greater net photosynthesis (>9 μmol m−2 s−1), greater predawn water potential (>−0.35 MPa), and lower mortality (2–3%) following one year in the field, indicating establishment success. The 80 cm3 stocktype remained significantly smaller and exhibited lower growth rates for the duration of the study, while all other stocktypes were statistically similar. Our results demonstrate the importance of controlling vegetative competition regardless of stocktype, especially for western larch, and suggest that benefits to post-planting seedling physiology and growth in relation to container size plateau beyond 130 cm3 among the investigated stocktypes.
1. Introduction
In order to ensure success, reforestation efforts must meet diverse objectives, which can involve myriad species, fit within a variety of economic models, and account for different site characteristics. Thus, tree seedlings used in these efforts should be produced with specific parameters in mind (genetics, morphology, and physiology, for example) [1,2,3]. Successful seedling stocking on a site within the required planting window may have economic, ecological, and legal implications—calling for high-quality, specifically-cultured plant material. Managers continually need updated, science-based information to justify on-the-ground decision-making; however, determining which traits lead to optimal seedling performance for a specific site and reforestation objectives continues to be a challenge.
Seedling phenotypic traits are a result of genetics and the environment. Nurseries have the capacity to alter these traits through modifications in nursery culture, including container size. Container-grown seedlings provide a wealth of stocktype choices in terms of dimensions (i.e., depth, diameter, volume) and container composition—factors which have been found to affect seedling phenotype [4,5,6]. Container selection in seedling production also has economic implications. Smaller-volume containers require fewer inputs per plant, maximizing nursery growing space while minimizing media and fertilizer; conversely, larger volume containers require more inputs, but produce larger seedlings [7,8,9,10,11,12]. The choice and difference between the two, however, can offer competitive advantages for survival and growth in the field. For larger containers, i.e., larger seedlings, such advantages stem from the ability to outcompete existing vegetation on the site as a result of higher nutrient reserves, increased photosynthetic capacity, and enhanced water use efficiency [3,13,14,15]. Villar-Salvador et al. [16] also argue that larger seedlings do better in environments that experience seasonal drought. Many studies have shown that larger stocktype seedlings initially remain larger and grow at a more rapid rate following outplanting [8,17,18,19]; however, results are often species- and site-specific and are subject to change following the first growing season [17,20,21].
After outplanting, seedling establishment during the first growing season depends upon initiation and maintenance of a positive feedback loop with its new environment. This loop requires uptake of soil water to support increased stomatal conductance and photosynthesis. The newly-assimilated carbon allows for enhanced root growth, thus continually increasing seedling access to soil water [22]. Competing vegetation is often an obstacle in the effective establishment of this loop; additionally, regional climate regimes can also be of influence. In the Inland Northwest region of the United States, where pronounced summer moisture limitations are common [23], plants must establish adequate root systems before the onset of drought. This is especially imperative for seedlings outplanted in the spring, which have a limited window for root growth to occur. In such conditions, stocktypes with large or deep root systems can be advantageous for overcoming drought stress [24]. Several studies have examined the effects of stocktype size and site treatments aimed at controlling competing vegetation on seedling performance [14,15,25,26]. Much of this work points to stocktype differences in the ability of seedlings to establish root contact with soil water, either through enhancing functional capacity to promote root growth in order to reach deeper soil moisture reserves or through reducing vegetative competition to increase available soil moisture. This is manifested by improved performance (higher survival, growth, and carbon assimilation capacity) among larger seedlings relative to smaller stocktypes under high competition [14,15,26]. The benefits of larger stocktypes, however, are less clear in the absence of vegetative competition or ecosystems without water limitations [14,25].
Despite the abundance of stocktype studies to date, only a small group of them examine the morpho-physiological performance of container stocktypes under different levels of vegetative cover during summer drought [14,25,26]. Unfortunately, these studies, among others, contain some level of confounding, thereby significantly limiting the transferability of findings. Much of this methodology shows that, during nursery cultivation or genetic selection, biases are introduced, thus optimizing growth of a single container size (or stocktype) and creating sub-optimal conditions for all other sizes. Addressing this issue, Pinto et al. [27] outline key considerations for conducting stocktype studies that minimize confounding. With these considerations, our study aims to examine the role of container size on seedling performance, while reducing confounding commonly associated with nursery culture, and create distinct drought differences between vegetative competition treatments at the site level. In order to expand the utility of the study, we use only stocktypes considered operationally feasible and focus on species of interest specific to the Western United States. We hypothesized that (1) larger stocktypes would have an advantage in accessing soil water due to larger root systems and would outperform smaller stocktypes during the summer drought, and (2) that this difference would be greater under high vegetative competition and low soil moisture. Evaluations of seedling performance included survival, growth, net photosynthesis, and plant water potential.
2. Materials and Methods
2.1. Nursery Culture
Open-pollinated, orchard-grown Douglas fir (Pseudotsuga menziesii var. glauca (Mayr) Franco) (Potlatch Lot ID#: DF-CL-Z7, 1000–1200 m elevation) and western larch (Larix occidentalis Nutt.) (Potlatch Lot ID#: WL-09-75 Improved BC, 1100–1350 elevation) seed was obtained in March 2012. Seed was cold, moist stratified for 30 days at 1.7 °C, and sown five per cell into 10 Styroblock® trays (Beaver Plastics, Edmonton, AB, Canada). Recognizing the operational range of seedling containers used in the region, two different tray sizes (Styroblock® 415C and 515A tray models) were used to obtain four container volume treatments which had approximate starting cell volumes of 130 and 250 cm3. Two experimental container volumes were created by cutting off a 50 cm3 portion of 415C and 515A models, resulting in cell volumes of 80 and 200 cm3 to complement the original 130 and 250 cm3 cells (Table 1).

Table 1.
Container specifications used to produce Douglas fir and western larch seedlings. Two types of containers were used to create four container treatment volumes.
To account for differences in growing space by container size, sowing was staggered by one week for each container size, beginning with the 250 cm3 containers in late April for Douglas fir and mid-May for western larch. Growing medium consisted of sphagnum peat moss:vermiculite:aged fine bark (2:1:1, v:v:v; Sun Gro Horticulture, Bellevue, WA, USA) with a bulk density of 0.14 g cm−3. Following sowing, media surface and seeds were covered with a medium-sized forestry nursery grit (Target Products Ltd., Burnaby, BC, Canada). Seedlings were grown in a greenhouse at the University of Idaho Pitkin Forest Nursery, Moscow, Idaho (46.7255° N, 116.9563° W) from April to November 2011. Culturing temperatures averaged 15 and 26 °C (minimum and maximum, respectively) over this time period. Unless otherwise noted, all methods were identical for both species.
Trays were arranged on nursery benches in a randomized complete block design (RCBD), with the arrangement repeated for each species. Blocking was determined by proximity to a heating tube underneath the nursery benches, with two Styroblocks® of the same cell volume serving as a single block. Each of the four tested container volumes consisted of five blocks and individual seedlings were considered to be experimental units (3020 total seedlings per species). Within each block, container locations were re-randomized every two weeks.
Irrigation and fertigation timing was individually tailored to each block. The saturated weight of one Styroblock® in each pair was determined at the onset of nursery culture and re-weighed daily to determine gravimetric water content [28]. Throughout the growing season, seedlings were fertigated as they reached gravimetric targets (see below), usually 2–3 times per week.
Immediately following sowing, containers were misted using an overhead boom twice daily for two weeks and thinned to 2–3 trees per cell after this period. After the initial two weeks, seedlings were thinned to one tree per cell and received applications of Peters® Conifer StarterTM (10:20:30 [N:P2O5:K2O], The Scotts Company, Marysville, OH, USA) at 42 mg N L−1, for the following three weeks, as blocks reached 85% of their saturated weight. During the rapid growth phase, seedlings received applications of Wilbur Ellis® Pro-GrowerTM (20:7:19, Wilbur Ellis, Walnut Creek, CA, USA) at 60 mg N L−1 for western larch and 100 mg N L−1 for Douglas fir, and calcium nitrate (CN; 15:0:0) as containers dried to 80–85% of their saturated weight. Alternating applications of fertilizer during this stage was supplemented with Peters® S.T.E.M.TM (Soluble Trace Element Mix) micronutrient mix. As seedlings within container volume groups reached approximately two-thirds of their target heights (15 cm), excess nutrients were leached using a water flush, and seedlings were moved to the hardening phase fertilizer regime. During this phase, seedlings received Wilbur Ellis® Pro-FinisherTM (4:25:35) at 24 mg N L−1, applied when blocks dried down to 65 and eventually 55% to initiate budset.
2.2. Nursery Phase Sampling
Height and root-collar diameter (RCD) measurements were obtained for a random subsample of 20 seedlings per Styroblock® tray every other week beginning 10 weeks after sowing, through the end of the season. These measurements were used to determine fertilization phase timing.
Assessment of morphological characteristics for each seedling at the end of the nursery culture (December 2011) included height and RCD and the respective growth increments for each variable. In addition, a random subset of 25 seedlings from each container volume × block combination was subject to destructive sampling (n = 500). After carefully washing roots to remove media, root volume (RV) was determined using the water displacement method described by Burdett [29]. The sampled seedlings were oven dried at 60 °C for 72 h, after which root and shoot dry masses were measured. The root-to-shoot (R:S) biomass ratios were calculated for all destructively-sampled seedlings. Remaining seedlings were lifted, placed into plastic bags inside wax boxes, and stored at −1.4 °C (±0.5) for five months, in line with standard nursery protocol.
2.3. Outplanting
A total of 1600 seedlings were outplanted between 25 and 31 May 2012 on a 15-hectare cut site in the East Hatter Creek Unit of the University of Idaho Experimental Forest (46.8445° N, 116.7960° W, 860 m a.s.l.). The site had a south-facing aspect, ranged in slope from 5–20%, and was logged in August 2011, with approximately 100 trees remaining to meet leave-tree obligations of the Idaho Forest Practices Act. The residual slash was broadcast burned in October 2011. Tree species present prior to harvest included western larch and Douglas fir, as well as ponderosa pine (Pinus ponderosa Dougl. Ex Laws. var. ponderosa). Soils were classified in the Santa series of Alfisols, described as moderately well-drained and moderately deep, formed in deep loess with a small amount of volcanic ash in the upper horizons [30].
For each species, a split-plot, randomized complete block design was used. Each block was the whole-plot, which consisted of two 21 × 10 m areas that were randomly assigned one of the two vegetative competition treatments (glyphosate-sprayed or high competition). In the glyphosate-sprayed (GS) treatments, competing vegetation was removed with two applications (7 and 29 June 2012) of glyphosate (41%, Glystar® Plus, Albaugh, Inc., Ankeny, IA, USA) using a backpack sprayer (prior to tree planting), at a rate of 3.4 kg acid equivalent ha−1. Additional vegetation was removed manually throughout the season. In the high-competition (HC) treatments, the natural vegetation community was allowed to establish. In addition to this, we planted blue wildrye (Elymus glaucus Buckley (Clearwater Seed, Spokane, WA, USA)) seeds, a grass species native to the region, at an approximate density of 75 plants m−2 in May 2012. Our split-plot factor was the four container volumes. Within the GS and HC treatments, the four container volumes (80, 130, 200, 250 cm3) were randomly assigned to four rows, each row contained 20 seedlings from one of the four container sizes. Spacing was 1 and 2 m between seedlings and rows, respectively. To minimize browse damage, seedlings were surrounded with 1 m tall yellow mesh protection tubes (Forestry Suppliers, Inc., Jackson, MS, USA); additionally, a 2 m buffer of animal repellant (Plantskydd®, concentration of 0.125 kg L−1, Tree World, St. Joseph, MO, USA) was sprayed around each vegetation treatment using a backpack sprayer.
Existing vegetation cover was quantified using sampling approach described by Daubenmire [31]. A diagonal transect was established running from the northwestern-most corner of the plot to the southeastern-most corner. A 20 × 50 cm frame was placed at 4-, 8-, 12-, 16-, and 20-m intervals along the transect. At each sampling point, total cover of live vegetation in the frame, using the projected cover of foliage onto the ground below, was recorded as a percentage.
2.4. Edaphic and Atmospheric Monitoring
A weather station (model 2900ET, Spectrum Technologies, Inc., Plainfield, IL, USA), installed on site, was used to monitor hourly air temperature (°C), precipitation (mm), and relative humidity (%) during the first growing season. ECH20-TE soil moisture probes (Decagon Devices, Inc., Pullman, WA, USA) were installed in HC and GS plots that most closely represented the diversity of slope, soil moisture, and initial vegetative cover on the site. Volumetric soil moisture (θ, m3 m−3) measurements were collected hourly at three different soil depths (5, 15, and 30 cm) using an Em50 data logger (Decagon Devices, Inc., Pullman, WA, USA) from June to October 2012. In situ soil calibrations were performed to increase the accuracy of the volumetric soil moisture measurements.
2.5. Survival and Morphology Measurements
Height (cm) and RCD (mm) were measured immediately after planting (early June 2012), at the end of the first growing season (October 2012), and at the end of the second growing season (October 2013). Seedling survival was assessed at the end of each growing season. Height and RCD increment for all surviving trees was calculated by subtracting the initial from the final measurement for each surviving, un-sampled seedling.
2.6. Seedling Gas Exchange and Water Potential Measurements
Both seedling gas exchange and pre-dawn water potential (Ψpd) were measured, for each species, on one randomly-selected seedling from the two vegetative competition treatments × four container sizes × five replication blocks (n = 40, per species), three times throughout the season. These measurements corresponded with pre-drought (7 and 11 July), early drought (7 and 8 August), and late drought (25 and 26 September) periods. A fourth, post-drought (21 October), set of measurements was taken on Douglas fir seedlings only, since western larch seedlings had experienced high levels of mortality, and those surviving the first season had begun to senesce. For each measurement period, a new seedling was chosen and was no longer included in future measurements.
Pre-dawn water potential was measured using a pressure chamber (model 1505D-EXP, PMS Instrument Company, Corvallis, OR, USA) between 0000 and 0400 h. A small woody lateral branch from each seedling was excised and used for measurement (nb: seedlings with excised biomass were no longer used in subsequent measurements). Gas exchange measurements were conducted on the same seedlings using a portable photosynthesis instrument (model LI-6400XT, LI-COR Environmental, Lincoln, NE, USA) equipped with a lighted conifer chamber (model 6400–22L), a RGB light source, and a CO2 injector. Measurements began in the morning once photosynthetically active radiation (PAR) reached ≥800 μmol m−2 s−1 and were completed between 0800 and 1230 h. The upper 7 cm portion of the terminal leader was placed into the conifer chamber while still attached to the seedling for measurements because at the start of the season this was the only portion of the seedling tall enough to reach into the LI-6400 chamber. The chamber environment was set to 1400 μmol m−2 s−1 PAR, 400 μmol mol−1 CO2 with a flow rate of 400 μmol s−1, as described by Pinto et al. [15]. The temperature was initially set to 25 °C, but was raised, as the outside temperature rose (maximum 28 °C), to extend battery life. The portion of the branch placed inside the conifer chamber during measurements was then severed from the seedling. Leaf tissue was scanned on a flatbed scanner, and quantified using Image J software (Version 10.2, National Institutes of Health, Bethesda, MD, USA).
2.7. Statistical Analysis
All analyses, except where noted, were performed separately for each species. As well, for all analyses of variance (ANOVA), block effects were removed from models once found not to be significant. ANOVA using SAS (Version 9.3, SAS Institute, Inc., Cary, NC, USA) PROC MIXED for a RCBD (four container volumes × five blocks) was used to determine if differences in seedling morphology existed for stocktype treatments (p < 0.05) after nursery culture. Residual plots were used to assure data met model assumptions. Post-hoc means separations were performed using the Tukey HSD (α = 0.05).
Treatment differences for response variables from the outplanting experiment (seedling height, RCD, height and RCD increments, photosynthesis (A), and Ψpd) were analyzed using an ANOVA model including four container volumes × two vegetative competition treatments × five blocks within a RCBD split-plot design. Competition level served as the whole-plot factor while container volume was the split-plot factor. The design initially contained 20 seedlings (for height and RCD measurements) for all container × competition × replication combinations (n = 800, for each species). Height and RCD measurements one and two years following outplanting were collected from all surviving, un-sampled seedlings in each combination. For the physiological measurements, one tree per container volume × competition × block was used (n = 40, per species).
Differences between the effects of vegetative competition, on soil moisture throughout the season were analyzed via repeated measures ANOVA using PROC MIXED. Seedling survival was analyzed using logistic regression and a binomial distribution in PROC GLIMMIX. The model included the effects of container, competition, and their interaction. To avoid complete separation in this model, Douglas fir seedlings in the GS plot × 130 cm3 container volume treatment were excluded due to 0% mortality. Post-hoc, pair-wise comparisons were made using Tukey HSD (α = 0.05).
3. Results
3.1. Nursery Culture
3.1.1. Douglas Fir
Container volume significantly (p < 0.0001) influenced seedling morphology (height, RCD, RV, R:S) (Table 2). Seedlings cultivated in the smallest (80 cm3) containers exhibited the lower height, RCD, and RV than the other three stocktypes. The largest container volume produced the tallest seedlings compared to all others. The two larger stocktypes (200 and 250 cm3) exhibited higher RCD and RV values compared to the other two sizes and significantly higher R:S values compared to those reported for the smallest containers. The R:S values of the 130 cm3 stocktype were not significantly different from the other three.
Table 2.
Mean (±SE) height, root-collar diameter (RCD), root volume (RV), root dry mass:shoot dry mass (R:S) of Douglas fir and western larch seedlings at the end of one-year nursery culture across container types. Different letters within a species column indicate significant differences at α = 0.05.
3.1.2. Western Larch
Container volume had a significant effect on western larch seedling height, RCD, and RV values (p < 0.0001; Table 2). Generally, seedlings grown in 80 cm3 containers were significantly smaller in height, RCD, and RV values compared to all other stocktypes. There was no clear relationship between the increase in container volume and the assessed morphological responses. Seedlings grown in the 200 cm3 containers exhibited greater height, RCD, and RV values. With regard to height, the differences between the 130 cm3 and the two largest stocktypes were not statistically significant. Seedling R:S did not differ by container volume (p = 0.2509).
3.2. Site Conditions
At the time of planting, air temperature and vapor pressure deficit (VPD) were 13.1 °C and 0.3 kPa, respectively. Maximum air temperature (37.0 °C) and VPD (5.8 kPa) were reached on 8 July 2011. During the study period (25 May to 31 October), air temperature and VPD averaged 15.6 °C and 1.2 kPa, respectively. Mean maximum daily air temperature for the season was 24.2 °C, with mean maximum VPD of 2.6 kPa (Figure 1A,B).
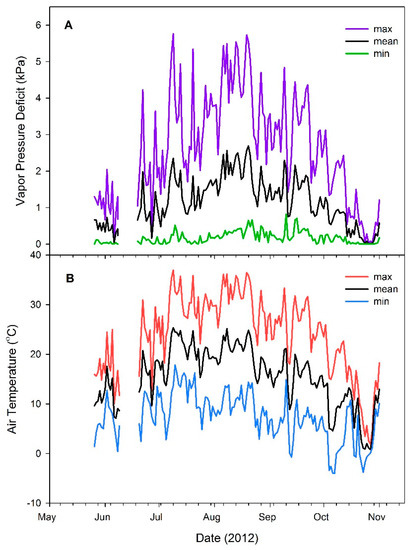
Figure 1.
Daily mean, minimum, and maximum vapor pressure deficit (A) and air temperature (B) conditions during the 2012 growing season at the outplanting site on the University of Idaho Experimental Forest, Moscow, ID, USA. Data from 9–18 June are missing due to ungulate-induced weather station damage.
From 25 May and 31 October, precipitation totaled 104 mm. No precipitation fell from 21 July through 12 October, with 63 mm falling before this drought period and 41 mm after (Figure 2A). Due to animal damage, no data were recorded from the weather station 9–18 June. Volumetric soil moisture content (θ) in the plots were compared at five dates aimed at capturing moisture content differences during the critical establishment period (i.e., seedling planting, pre-, early, late, and post-drought). For all three soil depths, the interaction of date × vegetative competition treatment was significant (p < 0.0001), as were the main effects of date (p < 0.0001) and vegetative competition (p < 0.04). In June and July 2012, θ did not differ by competition treatment at all depths. However, during the most pronounced drought period in August and September, θ was significantly higher in GS than in HC plots at all three soil depths (Figure 2). After precipitation resumed in October 2012, the two plot types again showed equal measures of θ for all three depths.
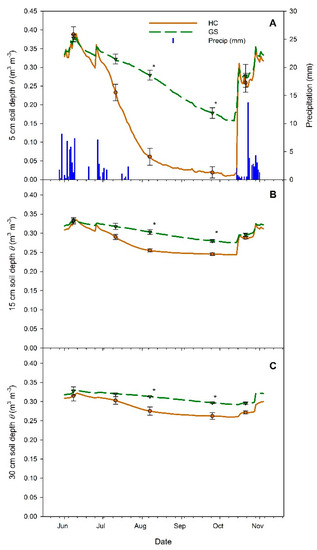
Figure 2.
Volumetric soil moisture content (θ) of both vegetative competition treatments (high competition (HC) = solid brown lines, circles; glyphosate-sprayed (GS) = dashed green lines, triangles) at 5 (A), 15 (B) and 30 cm (C) depths, and daily precipitation totals (A, vertical bars on the x-axis) during the 2012 growing season. Points with vertical error bars indicate dates where seedling physiological measurements were performed, as well as seedling planting in June. Points with asterisks (*) indicate a significant difference between HC and GS treatments for that date (α = 0.05, n = 3). Precipitation data from 9–18 June are missing due to weather station damage.
Differences in vegetation cover, assessed in August, existed between competition treatments. For Douglas-fir, GS plots had significantly less cover (6.6%) than HC plots (45.3%) (p = 0.0002). Similarly for western larch, GS plots had significantly less cover (10.1%) than HC plots (59.4%) (p = 0.0006).
3.3. Field Survival and Growth
3.3.1. Douglas Fir
By the end of the first growing season in the field, the interaction between container volume × vegetative competition did not significantly affect any of the morphological responses (Table 3). Container volume alone had a significant impact on height, height increment, and RCD, while vegetative competition influenced RCD, RCD increment, and mortality. Height, height increment, and RCD of seedlings cultured in 80 cm3 cavities were significantly smaller than the other three sizes (Table 3). There were no differences in RCD growth (increment) or mortality among stocktypes. Height and height growth (increment) did not differ significantly due to vegetative competition, but seedlings with larger RCD and RCD increment values were observed in GS plots compared to HC plots.
Table 3.
Mean (±SE) height, height increment, root-collar diameter (RCD), and RCD increment of surviving Douglas fir seedlings at end of the first and second field season (October 2012 and 2013) across container types and vegetative competition treatments (GS = glyphosate-sprayed; HC = high competition). The proportion of mortality at the end of the first and second field seasons are also shown. Different letters within a field season column indicate significant differences at α = 0.05.
Two years following outplanting, the combined effect of container volume × vegetative competition, and competition alone did not influence seedling morphology (Table 3). However, seedling height, height increment, RCD, and RCD increment were significantly influenced by container volume. Seedlings produced in the smallest containers remained significantly shorter compared to the other stocktypes. The smallest containers also yielded the lowest mean RCD, but did not differ from the 200 cm3 stocktype; while the 130 cm3 stocktype had the largest RCD and was similar to the 250 cm3 stocktypes. Height and RCD growth (increment) was significantly higher for the 130 cm3 compared to 200 cm3 stocktypes, while the smallest and largest container volumes were statistically indistinguishable from the others.
Mortality following the first growing season was significantly influenced by vegetative competition, evidenced by lower mortality among seedlings planted in the GS plots compared to those in HC plots (3% and 28%, respectively). At the end of the second field growing season, mortality increased two- and five-fold among the HC and GS treatments (61% and 15%, respectively).
3.3.2. Western Larch
At the end of the first field-growing season, mortality among larch vegetation treatments was high; therefore, our analyses were adjusted accordingly. In the full model analysis (mortality = container, vegetative competition, container × vegetative completion), there was no interaction or container volume effect on seedling mortality (p > 0.5172). Seedling mortality was affected by vegetative competition, with seedlings grown in HC plots exhibiting significantly higher mortality than those in GS plots (p < 0.0001) (98% and 2%, respectively). As a result of the near complete mortality in the HC plots, a priori analyses on the full statistical model for height, height increment, RCD, and RCD increment was not possible; thus, post hoc analyses focused only the effect of container volume within GS plots. Based on the surviving seedlings in these plots, container volume had a significant effect on height, RCD, and their growth increments (p < 0.0107; Table 4). Height and RCD were significantly lower among the smallest stocktype compared to all other sizes. The 130 and 200 cm3 containers produced the tallest seedlings, while the RCD values for the 130, 200, 250 cm3 stocktypes were statistically indistinguishable from each other. For height and RCD growth increments, the largest stocktype was significantly smaller than the 130 cm3 stocktype, whereas all other container volume treatments did not differ significantly.
Table 4.
Mean ± (SE) height, height increment, root-collar diameter (RCD), and RCD increment of surviving western larch seedlings at end of the first and second field seasons (October 2012 and 2013). Data is from glyphosate-sprayed (GS) plots only due to near complete mortality within the high competition (HC) plots. Different letters within a field season column indicate significant differences at α = 0.05.
As with the first year outplanting data, the full model analysis on mortality indicated no container volume × vegetative competition interaction (p = 0.5819), or container volume effects (p = 0.9227) two years following outplanting. Significant differences as a result of vegetative competition persisted at the end of the second field season. Mortality increased seven-fold between the first and second year within the GS plots, but still only amounted to 14% of the total seedlings planted; HC mortality totaled 99%. Again, because of the high mortality, post hoc analyses focused on GS container effects only. Two years following outplanting, container volume significantly influenced seedling morphology and growth (Table 4). Seedlings grown in the 80 cm3 containers exhibited the smallest height, RCD, and RCD increment; however, differences in height increment no longer varied among containers.
3.4. Physiology
3.4.1. Douglas Fir
For both field-measured physiological variables (i.e., A and Ψpd), there were no significant interactions between container volume × vegetative competition (p > 0.0748). The main effect of container volume was also not significant (p > 0.1123). Effects of vegetative competition on A were not significant in July (p = 0.2256), but were significant for August, September, and October 2012 measurements (p < 0.0056) (Figure 3). At these measurement points, seedlings in the GS plots exhibited significantly higher A rates than those in the HC plots (Figure 3A). Overall, GS plots exhibited a 128% increase in A from July to September, when soils were rapidly drying in the upper soil profile (Figure 2A). Conversely, HC plots exhibited a 39% decrease in A during the same period.
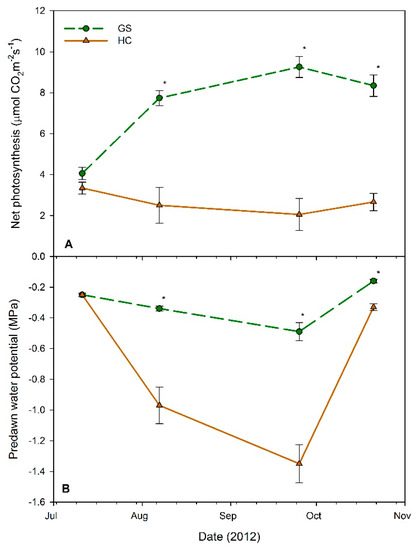
Figure 3.
(A) Net photosynthesis and (B) predawn water potential of Douglas fir seedlings at four measurement dates in 2012. Each point represents the mean (n = 19–20) across all container sizes, within each vegetative treatment, high competition (HC: solid brown lines, triangles), and glyphosate sprayed (GS: dashed green lines, circles). Error bars represent the standard error of the mean. Points with asterisks (*) indicate a significant difference between HC and GS treatments for that date (α = 0.05).
No differences in Ψpd between GS and HC were detected at the first measurement date in July (p = 1.000), but HC plots exhibited significantly lower values than GS for the remaining three measurement points (August, September, and October 2012; p < 0.0301), during and after the drought period. GS decreased 0.24 MPa (96%) from July to September, while HC decreased 1.10 MPa (440%). The September Ψpd values were most negative for both vegetative competition levels, respectively, but they both recovered to near July values by 21 October (Figure 3B), after 4.6 mm of precipitation fell between 13–21 October (Figure 2).
3.4.2. Western Larch
Physiological data for western larch was only collected for the first three measurement points (i.e., pre-, early, and late drought) due to high seedling mortality and the onset of fall senescence at the last measurement date (October 2012). For all three measurement points, both net photosynthesis and Ψpd showed no significant interactions between container volume and vegetative competition (p > 0.1580); container volume alone had no impact (p > 0.0744). Conversely, vegetative competition influenced both photosynthesis and Ψpd measurements in August (p = 0.0073 and p = 0.0068) and September (p = 0.0125 and p = 0.0018) (Figure 4). Seedlings in the HC plots exhibited a 78% decrease in net photosynthesis from July to September, while those in GS plots experienced a 68% increase in photosynthesis during the same period (Figure 4A). The Ψpd values stayed relatively similar through the three measurement periods for GS seedlings, changing a maximum of 0.06 MPa (20%), while those in the HC plots became significantly more negative during the August and September measurement points, changing a maximum of 1.98 MPa (615%) (Figure 4B). Seedlings in both vegetative competition treatments had their most negative Ψpd measurements in September.
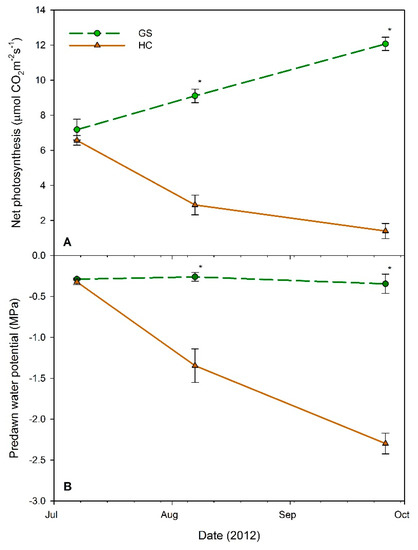
Figure 4.
(A) Net photosynthesis and (B) predawn water potential of western larch seedlings at three measurement dates in 2012. Each point represents the mean (n = 13–20) across all container sizes, within each vegetative treatment, high competition (HC: solid brown lines, triangles), and glyphosate sprayed (GS: dashed green lines, circles). Error bars represent the standard error of the mean. Points with asterisks (*) indicate a significant difference between HC and GS treatments for that date (α = 0.05).
4. Discussion
Following nursery culture, the positive correlation between seedling size and increasing container volume is well-documented [7,10,17,32]. In this study, we detected the same trend in Douglas fir and western larch with regard to the morphological traits measured. However, the largest containers were not often statistically set apart from the next smallest size. In fact, western larch seedlings followed a slightly different pattern of growth with container size. In general, morphological traits increased from the 80 to 200 cm3 container sizes, but then decreased slightly for the largest size (250 cm3). Despite our best efforts to equilibrate seedling quality across container types, according to the recommendations by Pinto et al. [27], we suspect the largest stocktype may not have reached its fullest size potential during nursery culture (for either species). It is difficult to know whether growth times or nutrient delivery contributed to this scenario. In some cases, situations like this can make outplanting performance interpretation tenuous. Lamhamedi et al. [33] experienced similar issues with large experimental stocktypes. The subsequent performance attributes show that there may have been a lasting effect among the largest stocktype in our study, but for only the first year of growth.
Despite the initial height and RCD differences among Douglas fir stocktypes at planting, the stratification of these traits began to dissipate after one field season. After two seasons, height, RCD, and their growth increments were not statistically different amongst stocktypes, with the exception of the smallest. Other studies with Douglas fir show similar trends where growth rates, as well as absolute morphological characteristics of different stocktypes, tend to equilibrate over time [11,34]. Despite the growth similarities among the Douglas fir stocktypes, we observed noticeable height and diameter growth among seedlings produced in the 130 cm3 containers following the second growing season. Although not statistically different, this may suggest that relative growth rates of smaller seedlings can be as good as, or greater than, those of larger seedlings, reducing the seedling size gap over time [19,35,36].
While seedling height and shoot growth appear to be more influenced by container volume, survival and below-ground production are likely influenced by competition. After one growing season, the positive effect of reducing vegetative competition on Douglas fir size and growth was seen only in terms of RCD and seedling survival, but not seedling height. This supports previous findings that seedling height is not a strong predictor of performance in all cases [25,26,37,38]. Conversely, RCD has been used as a better predictor of early seedling health and survival [24,39]. Seedlings with greater RCD, which is correlated to higher root mass, have been shown to exhibit increased hydraulic conductivity, reduced transplanting shock, and improved survival under moisture stress [40,41,42,43,44].
For western larch seedlings, the role of container volume and its interaction with vegetative competition could not be fully assessed due to near 100% mortality under high competition in the first year. Our investigation of the larch stocktypes grown with reduced competition (GS), yielded similar results to the Douglas fir, where the initial size differences between stocktypes largely dissipated, and growth mostly equilibrated over two years. However, despite reduced soil moisture stress as a result of lower competition, the smallest stocktype consistently underperformed and remained small.
Generally, western larch has been found to be less drought tolerant than Douglas fir [45]. This may partly explain why drought has been attributed to high levels of mortality in western larch during the first season following outplanting [46,47]. In our study, the period between July and September was likely accountable for the majority of seedling mortality in the first year. During this period, the Ψpd values fell below −2.0 MPa, which may have led to stomatal closure and the subsequent reduction in net photosynthesis to below 4 μmol CO2 m−2 s−1, indicating that seedlings were unable to effectively manage moisture stress. Pinto et al. [48] modeled similar conditions for ponderosa pine seedlings and predicted that dry soils in the upper soil profile (0–15 cm) would reduce seedling photosynthesis and transpirational relationships.
While our photosynthesis rates measured were comparable with those reported in the literature [49,50], we did not observe a combined effect of vegetative competition × container volume, or container volume alone on the net photosynthesis measurements of either species. Although seedling stocktypes differed in initial size, their carbon allocation rates did not. This is contrary to several comparable studies that observed a significant influence of stocktype × competition interaction on photosynthesis rates [14,15,25,26]. However, despite the significance of the interaction there is uniform correlation that is shared by these studies. For example, Cuesta et al. [14] observed higher photosynthesis rates for larger versus smaller stocktypes under high competition, with no differences in performance in the absence of vegetative competition—an outcome expected in this study. Mohammed et al. [26] saw increased photosynthesis rates in a smaller (57 cm3) stocktype under reduced competition, while a larger (98 cm3) stocktype exhibited no change between competition treatments. Lamhamedi et al. [25] saw no difference between black spruce produced in 110 or 300 cm3 containers either with or without competition, but noted that in the presence of vegetative competition the largest (700 cm3) stocktype performed worse compared to the smaller sizes. Finally, Pinto et al. [15] reported that high levels of vegetative competition resulted in complete mortality among smaller stocktypes (60 and 90 cm3) of ponderosa pine, whereas the largest stocktype (120 cm3) exhibited increased survival.
Soil moisture in the upper profile was not markedly different between competition treatments early in the growing season. As the season progressed, however, differences between the treatments became pronounced in the upper soil profile, and were larger than those at greater depths (>15 cm; Figure 2). Due to the drastic changes observed in seedling physiology over time (seedling A and Ψpd), and consequent survival (for western larch), there is evidence to suggest that a large portion of the seedlings’ functional root system was in the upper profile. For example, our reported Ψpd measurements for Douglas fir are similar to those observed in seedlings under high moisture stress [13,51]. This corroborates findings of Pinto et al. [48], which show a strong correlation between rooting depth, soil moisture, and physiological functioning. This also shows the importance of considering the interaction of edaphic characteristics of a site with stocktype selection and site preparation.
Successful seedling establishment depends on the ability of seedlings to quickly become coupled to the site hydrological cycle and respond to environmental and silvicultural conditions [52]. The low levels of photosynthesis and Ψpd, along with increased mortality (especially in western larch) under a high level of competition indicate that the seedlings did not have ample resources and time to establish prior to the onset of summer drought. Similar findings have been observed among ponderosa pine, Aleppo pine, and Holm oak [14,15]. Along these lines, root growth is sensitive to both plant moisture stress [22,53] and soil temperature [52,54]. Our data suggests that seedlings outplanted into reduced competition conditions were able to access soil water in the upper profile, which had significantly greater θ than those observed in the high competition plots, which promoted greater root egress into the soil. Pinto et al. [15] reported that out of the three tested stocktypes that varied in volume and length, only the largest, longest stocktype survived in high competition. That same stocktype would have had roots that extended well into the zone of higher soil moisture measured within the scope of our study, even under high competition. Thus, future work should focus on examining the performance of taller containers, especially in conditions where vegetation control methods are not available.
Interestingly, Douglas fir seedlings in HC plots experienced a recovery to pre-drought levels of Ψpd in October, after rainfall resumed, but this was not accompanied by an increase in photosynthesis rates. This contrasts findings by Pinto et al. [15], who reported an increase in post-drought photosynthesis rates for the largest stocktype of ponderosa pine seedlings in high competition plots in October. The difference in these reports could be related to the number of precipitation events. Pinto et al. [15] stated that seedlings received several small rain events throughout the drought season, and prior to the final gas exchange measurement. Drought conditions within our study were more pronounced, with a complete absence of precipitation lasting 87 days during the first growing season. In addition to species-specific drought response strategies [45], other studies have shown that photosynthetic recovery is strongly influenced by the length and severity of drought [55]. The mechanism behind this damage is attributed to restricted CO2 diffusion into the plant, a depletion of carbon reserves, and general metabolic impairment over time [56,57]. However, less severe intermittent drought, followed by increased water availability, may allow for seedling acclimatization to drought and eliminate the reduction in photosynthesis rates [58].
5. Conclusions
On sites with high vegetative competition resulting in depleted upper-profile soil moisture, seedling mortality can be excessively high leading to establishment failure. This study used vegetative competition to further increase summer drought conditions in order to isolate stocktype- and species-driven differences in performance following outplanting while using uniformly-cultured nursery seedlings, in an effort to reduce confounding. Our results indicate that the largest operational stocktype evaluated in this study did not offer an advantage to overcoming drought conditions observed in the first two growing seasons. When competing vegetation was controlled, ample soil moisture remained in the upper profile, which refuted the initial differences in seedling size for all but the smallest containers. Thus, western larch and, to a lesser extent, Douglas fir seedlings planted on sites with moisture limitations require control of vegetative competition prior to spring planting. Furthermore, in the Inland Northwest, Douglas fir and western larch seedlings selected for spring planting may not require container volumes beyond 130 cm3, especially in the absence of vegetative competition.
Author Contributions
B.A.M., A.S.D., and J.R.P. conceived and designed the experiment; B.A.M. and J.R.P. implemented the experiment; B.A.M, J.R.P., and O.A.K. analyzed data; B.A.M. wrote the original paper as part of her thesis; A.S.D. coordinated the work; and all authors contributed to the final version of the manuscript.
Acknowledgments
We thank the Potlatch Corporation for funding and providing seed for the project, and Abbie Acuff, in particular, for the support of this work; University of Idaho Center for Forest Nursery and Seedling Research for nursery culturing space and logistical support; and University of Idaho Experimental Forest for the planting site. We are grateful for the data collection assistance in the nursery and field provided by Don Regan, Jake Kleinknecht, and Shannin Murphy. Thanks to Amy Ross-Davis, who contributed to an extensive revision of the manuscript in an earlier form; Douglass Jacobs and Randy Brooks provided support and insight during the study development. Further support was provided by the USDA Forest Service, Rocky Mountain Research Station (RMRS) and the National Center for Reforestation, Nurseries, and Genetic Resources. The views expressed are strictly those of the authors and do not necessarily represent the positions or policy of their respective institutions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Landis, T.D.; Tinus, R.W.; McDonald, S.E.; Barnett, J.P. Containers and growing media. In The Container Tree Nursery Manual; USDA Forest Service Agricultural Handbook 674: Washington, DC, USA, 1990; Volume 2, p. 88. [Google Scholar]
- Rose, R.; Carlson, W.C.; Morgan, P. The target seedling concept. In Target Seedling Symposium: Proceedings, Combined Meeting Western Forest Nursery Associations, Roseburg, OR, USA, 13–17 August 1990; Rose, R., Campbell, S.J., Landis, T.D., Eds.; USDA Forest Service General Technical Report RM-200; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1990. [Google Scholar]
- Thiffault, N. Stock type in intensive silivculture: A (short) discussion about roots and size. For. Chron. 2004, 8, 463–468. [Google Scholar] [CrossRef]
- Endean, F.; Carlson, L.W. The effect of rooting volume on the early growth of lodgepole pine seedlings. Can. J. For. Res. 1975, 5, 55–60. [Google Scholar] [CrossRef]
- Jinks, R.; Mason, B. Effects of seedling density on the growth of Corsican pine (Pinus nigra var. maritima Melv.), Scots pine (Pinus sylvestris L.) and Douglas-fir (Pseudotsuga menziesii Franco) in containers. Ann. For. Sci. 1998, 55, 407–423. [Google Scholar] [CrossRef]
- Chirino, E.; Vilagrosa, A.; Hernández, E.I.; Matos, A.; Vallejo, V.R. Effects of a deep container on morpho-functional characteristics and root colonization in Quercus suber L. seedlings for reforestation in Mediterranean climate. For. Ecol. Manag. 2008, 256, 779–785. [Google Scholar] [CrossRef]
- Carlson, L.W.; Endean, F. The effect of rooting volume and container configuration on the early growth of white spruce seedlings. Can. J. For. Res. 1976, 6, 221–224. [Google Scholar] [CrossRef]
- Simpson, D. Nursery growing density and container volume affect nursery and field growth of Douglas-fir and lodgepole Pine seedlings. In National Proceedings: Forest and Conservation Nursery Associations, Williamsburg, VA, USA, 11–14 July 1994; Landis, T.D., Dumroese, R.K., Tech. Coords., Eds.; USDA Forest Service General Technical Report RM-GTR-257; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1994. [Google Scholar]
- Aphalo, P.; Rikala, R. Field performance of silver-birch planting-stock grown at different spacing and in containers of different volume. New For. 2003, 25, 93–108. [Google Scholar] [CrossRef]
- Dominguez-Lerena, S.; Herrerosierra, N.; Carrascomanzano, I.; Ocanabueno, L.; Penuelasrubira, J.; Mexal, J. Container characteristics influence seedling development in the nursery and field. For. Ecol. Manag. 2006, 221, 63–71. [Google Scholar] [CrossRef]
- Haase, D.L.; Rose, R.; Trobaugh, J. Field Performance of Three Stock Sizes of Douglas-fir Container Seedlings Grown with Slow-release Fertilizer in the Nursery Growing Medium. New For. 2006, 31, 1–24. [Google Scholar] [CrossRef]
- Puértolas, J.; Jacobs, D.F.; Benito, L.F.; Peñuelas, J.L. Cost–benefit analysis of different container capacities and fertilization regimes in Pinus stock-type production for forest restoration in dry Mediterranean areas. Ecol. Eng. 2012, 44, 210–215. [Google Scholar] [CrossRef]
- Carlson, W.C.; Miller, D.E. Target seedling root system size, hydraulic conductivity, and water use during seedling establishment. In Target Seedling Symposium: Proceedings, Combined Meeting Western Forest Nursery Associations, Roseburg, OR, USA, 13–17 August 1990; Rose, R., Campbell, S.J., Landis, T.D., Eds.; USDA Forest Service General Technical Report RM-200; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1990. [Google Scholar]
- Cuesta, B.; Villar-Salvador, P.; Puértolas, J.; Jacobs, D.F.; Rey Benayas, J.M. Why do large, nitrogen rich seedlings better resist stressful transplanting conditions? A physiological analysis in two functionally contrasting Mediterranean forest species. For. Ecol. Manag. 2010, 260, 71–78. [Google Scholar] [CrossRef]
- Pinto, J.R.; Marshall, J.D.; Dumroese, R.K.; Davis, A.S.; Cobos, D.R. Photosynthetic response, carbon isotopic composition, survival, and growth of three stock types under water stress enhanced by vegetative competition. Can. J. For. Res. 2012, 42, 333–344. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Puértolas, J.; Cuesta, B.; Peñuelas, J.L.; Uscola, M.; Heredia-Guerrero, N.; Rey Benayas, J.M. Increase in size and nitrogen concentration enhances seedling survival in Mediterranean plantations. Insights from an ecophysiological conceptual model of plant survival. New For. 2012, 43, 755–770. [Google Scholar] [CrossRef]
- Sutherland, C.; Day, R.J. Container volume affects survival and growth of white spruce, black spruce, and jack pine seedlings: A literature review. North. J. Appl. For. 1988, 5, 185–189. [Google Scholar] [CrossRef]
- Jobidon, R.; Roy, V.; Cyr, G. Net effect of competing vegetation on selected environmental conditions and performance of four spruce seedling stock sizes after eight years in Québec (Canada). Ann. For. Sci. 2003, 60, 691–699. [Google Scholar] [CrossRef]
- Close, D.C.; Paterson, S.; Corkrey, R.; McArthur, C. Influences of seedling size, container type and mammal browsing on the establishment of Eucalyptus globulus in plantation forestry. New For. 2010, 39, 105–115. [Google Scholar] [CrossRef]
- Van den Driessche, R. Relationship between spacing and nitrogen fertilization of seedlings in the nursery, seedling mineral nutrition, and outplanting performance. Can. J. For. Res. 1984, 14, 431–436. [Google Scholar] [CrossRef]
- Pinto, J.R.; Marshall, J.D.; Dumroese, R.K.; Davis, A.S.; Cobos, D.R. Establishment and growth of container seedlings for reforestation: A function of stocktype and edaphic conditions. For. Ecol. Manag. 2011, 261, 1876–1884. [Google Scholar] [CrossRef]
- Burdett, A.N. Physiological processes in plantation establishment and the development of specifications for forest planting stock. Can. J. For. Res. 1990, 20, 415–427. [Google Scholar] [CrossRef]
- Ferguson, S.A. Climatology of the Interior Columbia River Basin; USDA Forest Service General Technical Report PNW-GTR-445; USDA Forest Service: Portland, OR, USA, 1999.
- Grossnickle, S.C. Why seedlings survive: Influence of plant attributes. New For. 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Lamhamedi, M.S.; Bernier, P.Y.; Hébert, C.; Jobidon, R. Physiological and growth responses of three sizes of containerized Picea mariana seedlings outplanted with and without vegetation control. For. Ecol. Manag. 1998, 110, 13–23. [Google Scholar] [CrossRef]
- Mohammed, G.H.; Noland, T.L.; Wagner, R.G. Physiological perturbation in jack pine (Pinus banksiana Lamb.) in the presence of competing herbaceous vegetation. For. Ecol. Manag. 1998, 103, 77–85. [Google Scholar] [CrossRef]
- Pinto, J.R.; Dumroese, R.K.; Davis, A.S.; Landis, T.D. Conducting seedling stocktype trials: A new approach to an old question. J. For. 2011, 109, 293–299. [Google Scholar]
- Dumroese, R.K.; Montville, M.E.; Pinto, J.R. Using container weights to determine irrigation needs: A simple method. Native Plants J. 2015, 16, 67–71. [Google Scholar] [CrossRef]
- Burdett, A.N. A nondestructive method for measuring the volume of intact plant parts. Can. J. For. Res. 1979, 9, 120–122. [Google Scholar] [CrossRef]
- Web Soil Survey: Natural Resources Conservation Service (NRCS) Soil Survey Staff. United States Department of Agriculture. Available online: http://websoilsurvey.nrcs.usda.gov/ (accessed on 8 February 2012).
- Daubenmire, R. A canopy-coverage method of vegetational analysis. Northwest Sci. 1959, 33, 43–64. [Google Scholar]
- Paterson, J. Growing environment and container type influence field performance of black spruce container stock. New For. 1997, 13, 329–339. [Google Scholar] [CrossRef]
- Lamhamedi, M.S.; Bernier, P.Y.; Herbert, C. Effect of shoot size on the gas exchange and growth of containerized Picea mariana seedlings under different watering regimes. New For. 1997, 13, 207–221. [Google Scholar] [CrossRef]
- Rose, R.; Haase, D.L.; Kroiher, F.; Sabin, T. Root volume and growth of Ponderosa pine and Douglas-fir seedlings: A summary of eight growing seasons. West. J. Appl. For. 1997, 12, 69–73. [Google Scholar] [CrossRef][Green Version]
- Van den Driessche, R. Absolute and relative growth of Douglas-fir seedlings of different sizes. Tree Physiol. 1992, 10, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Faure-Lacroix, J.; Tremblay, J.P.; Thiffault, N.; Roy, V. Stock type performance in addressing top-down and bottom-up factors for the restoration of indigenous trees. For. Ecol. Manag. 2013, 307, 333–340. [Google Scholar] [CrossRef]
- Overton, W.S.; Ching, K.K. Analysis of differences in height growth among populations in a nursery selection study of Douglas-fir. For. Sci. 1978, 24, 497–509. [Google Scholar] [CrossRef]
- Rose, R.; Gleason, J.F.; Atkinson, M. Morphological and water-stress characteristics of three Douglas-fir stocktypes in relation to seedling performance under different soil moisture conditions. New For. 1993, 7, 1–17. [Google Scholar] [CrossRef]
- Chavasse, C.G.R. The significance of planting height as an indicator of subsequent seedling growth. N. Z. J For. Sci. 1977, 22, 283–296. [Google Scholar]
- Carlson, W.C. Root system considerations in the quality of loblolly pine seedlings. South. J. Appl. For. 1986, 10, 87–92. [Google Scholar] [CrossRef]
- Mexal, J.G.; Landis, T.D. Target seedling concepts: Height and diameter. In Target Seedling Symposium: Proceedings, Combined Meeting Western Forest Nursery Associations, Roseburg, OR, USA, 13–17 August 1990; Rose, R., Campbell, S.J., Landis, T.D., Eds.; USDA Forest Service General Technical Report RM-200; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1990; pp. 17–35. [Google Scholar]
- Haase, D.L.; Rose, R. Soil moisture stress induces transplant shock in stored and unstored 2 + 0 Douglas-fir seedlings of varying root volumes. For. Sci. 1993, 39, 275–294. [Google Scholar]
- South, D.; Harris, S.; Barnett, J.; Hainds, M.; Gjerstad, D. Effect of container type and seedling size on survival and early height growth of seedlings in Alabama, U.S.A. For. Ecol. Manag. 2005, 204, 385–398. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Piñol, J.; Sala, A. Ecological implications of xylem cavitation for several Pinaceae in the Pacific Northern USA. Funct. Ecol. 2000, 14, 538–545. [Google Scholar] [CrossRef]
- Schmidt, W.C.; Shearer, R.C.; Roe, A.L. Ecology and Silviculture of Western Larch Forests; Technical Bulletin. 1520; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1976; 96p.
- Schmidt, W.C. Larix occidentalis . In Silvics of North America: Volume 1. Conifers; Burns, R.M., Honkala, B.H., Eds.; Agricultural Handbook 654; USDA Forest Service: Washington, DC, USA, 1990; p. 877. [Google Scholar]
- Pinto, J.R.; Marshall, J.D.; Dumroese, R.K.; Davis, A.S.; Cobos, D.R. Seedling establishment and physiological responses to temporal and spatial soil moisture changes. New For. 2016, 47, 223–241. [Google Scholar] [CrossRef]
- Rosenthal, S.I.; Camm, E.L. Photosynthetic decline and pigment loss during autumn foliar senescence in western larch (Larix occidentalis). Tree Physiol. 1997, 17, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Robertson, N.D.; Davis, A.S. Sulfometuron methyl influences seedling growth and leaf function of three conifer species. New For. 2011, 43, 185–195. [Google Scholar] [CrossRef]
- Dumroese, R.K.; Haase, D.L.; Landis, T.D. Seedling processing, storage, and outplanting. In The Container Tree Nursery Manual; USDA Forest Service Agricultural Handbook: Washington, DC, USA, 2010; Volume 7, p. 674. [Google Scholar]
- Rietveld, W.J. Transplanting stress in bareroot conifer seedlings: Its development and progression to establishment. North. J. Appl. For. 1989, 6, 99–107. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Importance of root growth in overcoming planting stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Sayer, M.S.; Brissette, J.C.; Barnett, J.P. Root growth and hydraulic conductivity of southern pine seedlings in response to soil temperature and water availability after planting. New For. 2005, 30, 253–272. [Google Scholar] [CrossRef]
- Rouhi, V.; Samson, R.; Lemeur, R.; Van Damme, P. Photosynthetic gas exchange characteristics in three different almond species during drought stress and subsequent recovery. Environ. Exp. Bot. 2007, 59, 117–129. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Galmes, J.; Medrano, H.; Ribas-Carbo, M. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress. Physiol. Plant. 2006, 127, 343–352. [Google Scholar] [CrossRef]
- Niinemets, U. Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: Past stress history, stress interactions, tolerance and acclimation. For. Ecol. Manag. 2010, 260, 1623–1639. [Google Scholar] [CrossRef]
- Stewart, J.D.; Zine el Abidine, A.; Bernier, P.Y. Stomatal and mesophyll limitations of photosynthesis in black spruce seedlings during multiple cycles of drought. Tree Physiol. 1995, 15, 57–64. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).