1. Introduction
Measures of above-ground biomass are needed for estimating site productivity, and stand and tree growth and yield [
1]. Estimates of individual tree and component biomass are of interest to researchers, managers and policymakers [
2]. For example, crown biomass estimates are necessary for determining the amount of logging residues, planning prescribed fire, and for biomass accounting in bioenergy production [
3,
4]. Accurate estimates of tree biomass are essential to understanding and predicting forest carbon (C) stocks and dynamics [
5,
6].
Often, local functions used to estimate tree biomass rely on the stem diameter over-bark at 1.37 m height (DBH) [
2,
7], or DBH and total tree height (HT) as explanatory variables [
8,
9]. These models are widely used but limited to certain stand characteristics and geographical areas, particularly those from which the data originated. However, inclusion of additional stand variables in the models such as stand age, density and/or productivity may result in general models that provide more accurate predictions [
10,
11,
12]. Moreover, Gonzalez-Benecke et al. [
10] concluded that stand characteristics used as covariates in general biomass functions resulted in significant improvements in model fitting and prediction ability for all tree components, especially when age and HT were unknown.
Longleaf pine (
Pinus palustris Mill.) was once a dominant forest type in the southeast U.S., occupying about 36 million ha prior to European settlement [
13]. Currently, there are only about 1.2 million ha of longleaf pine forest, and various organizations have begun promoting longleaf plantation establishment for forest products and a variety of ecosystem services [
14]. To our knowledge, although local equations (i.e., only using DBH and/or HT) are available for longleaf pine, no general biomass model (i.e., in addition to DBH and/or HT, including age and stand attributes) has been developed for the species. Due to stand age, structure and productivity effects on crown architecture, stem tapering and wood density, we expect that having general biomass functions (i.e., functions that include stand attributes as age, density and productivity metrics) is possible to improve the accuracy of biomass predictions, expanding the applicability of the models to broader geographic areas. Therefore, the objective of this study was to develop a set of local and general biomass equations to estimate total above-stump dry mass, and component biomass for longleaf pine trees in order to provide optional models that the user can apply depending on data availability. These local and general individual-tree models can be applied to trees growing over a large geographical area and having a wide range of ages and stand characteristics. Thus, they can be used in a variety of ecological, silvicultural and economics applications. These applications include estimations of C budgets for life cycle analysis and regional assessments of net primary productivity. The set of equations presented in this study provide a consistent basis for evaluations of longleaf pine forest biomass, improving the confidence in multi-scale analysis of C exchange between the forest and atmosphere.
4. Discussion
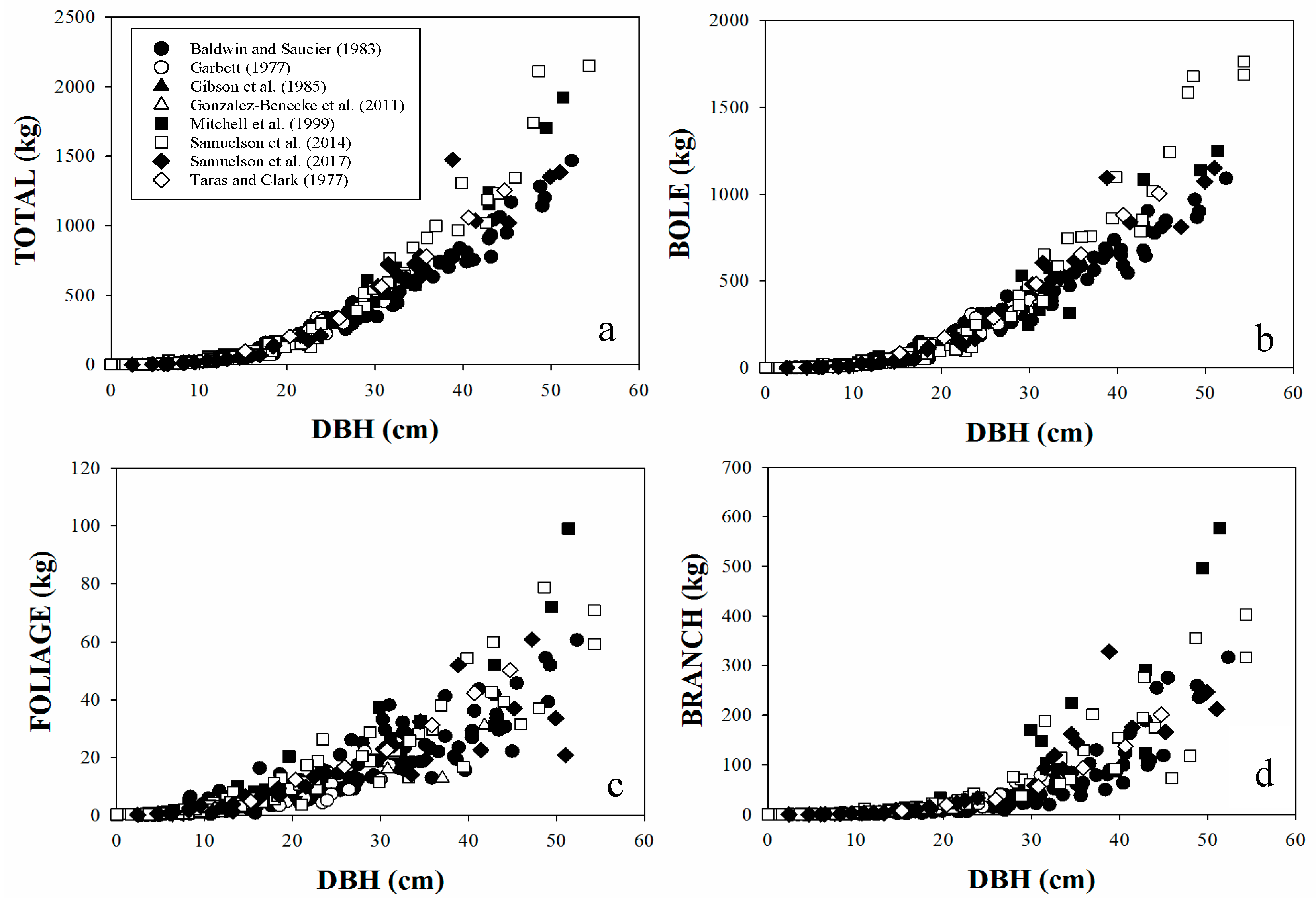
General and local additive models are useful to determine bole, living branches, foliage and total above-ground tree components. The additive models used in this study ensure that total biomass will be equal to the sum of the parts. Depending on data availability and level of accuracy desired, the users should decide which model to apply.
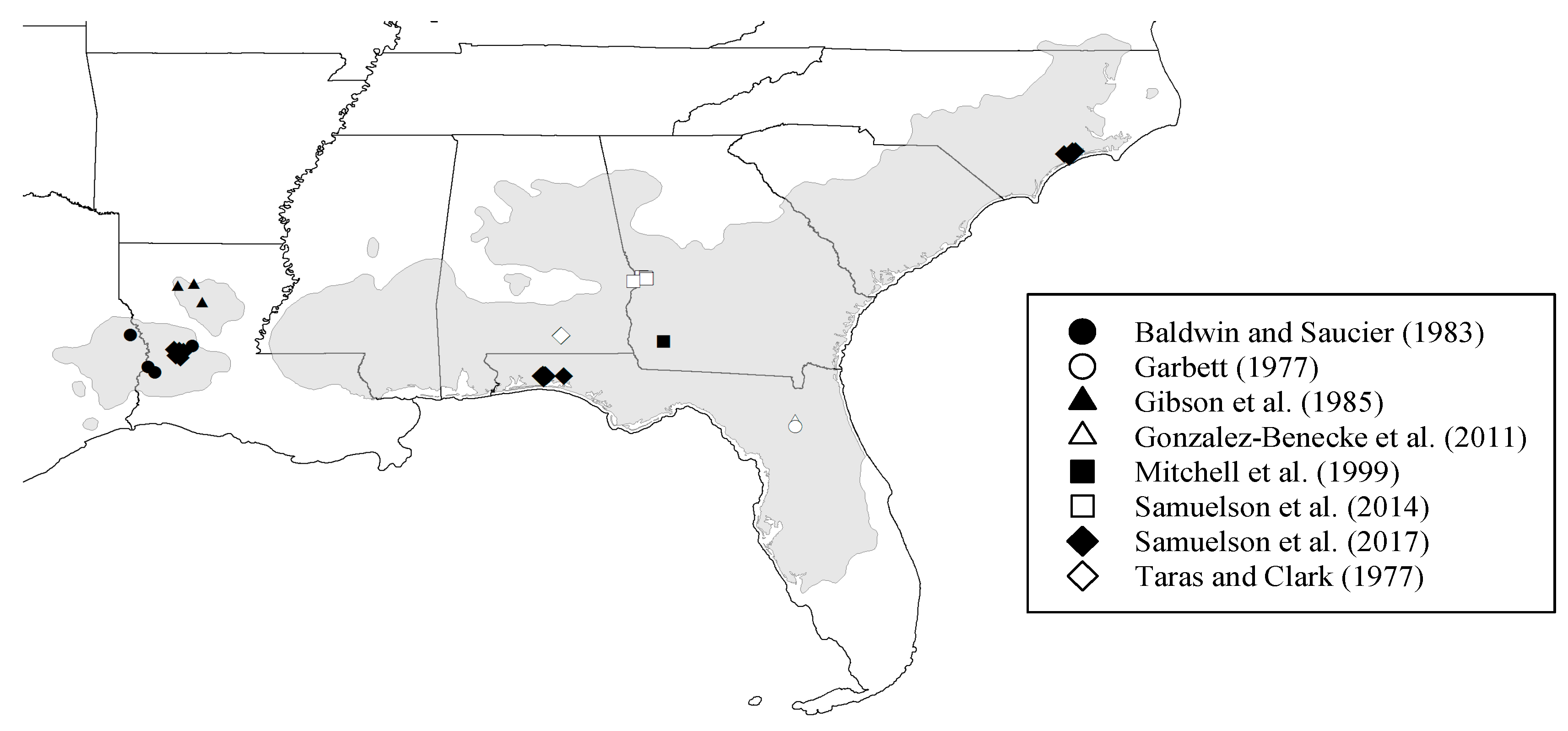
The set of biomass equations were developed using data from trees growing on sites distributed over the range of the species distribution across the southeastern U.S., spanning a wide range of ages and stand characteristics. This comprehensive data analysis provides confidence that the biomass functions reported here can be used over a broad range of stand development and management conditions. As thinning may affect stand structure, and we do not have records of years since thinning on the sampled trees, we recommend using our general models with caution on conditions soon after thinning.
Similar to Zhao et al. [
21] and Poudel and Temesgen [
22], in this study we used the biomass additivity approach, which guarantees that total above-ground biomass will be equal to the sum of the parts (bole, living branches and foliage). As Parresol [
20] concluded, biomass additivity is a preferred attribute of a biomass equations system. All previous biomass equations reported for longleaf pine including the biomass equations of Baldwin and Saucier [
8] and Samuelson et al. [
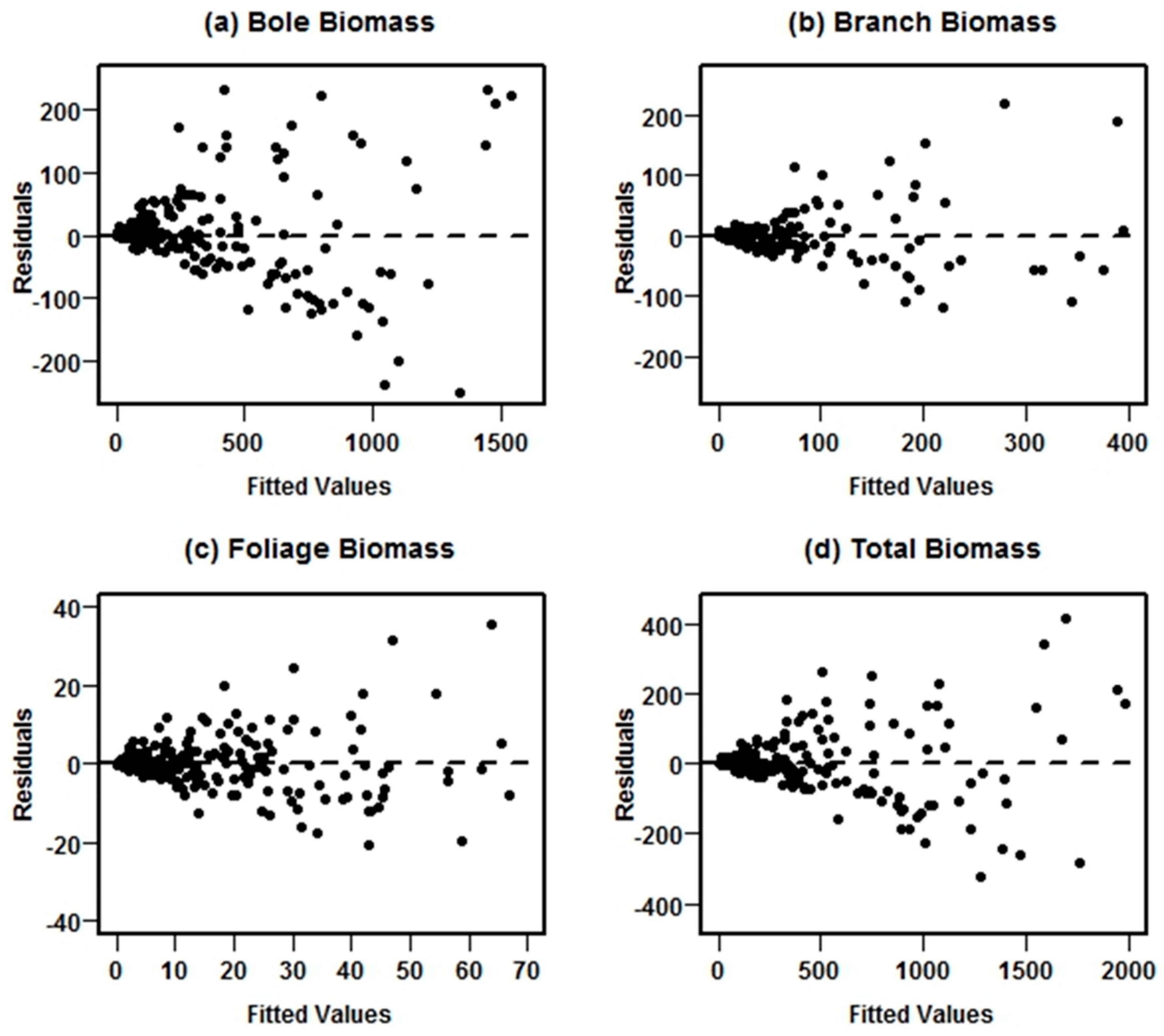
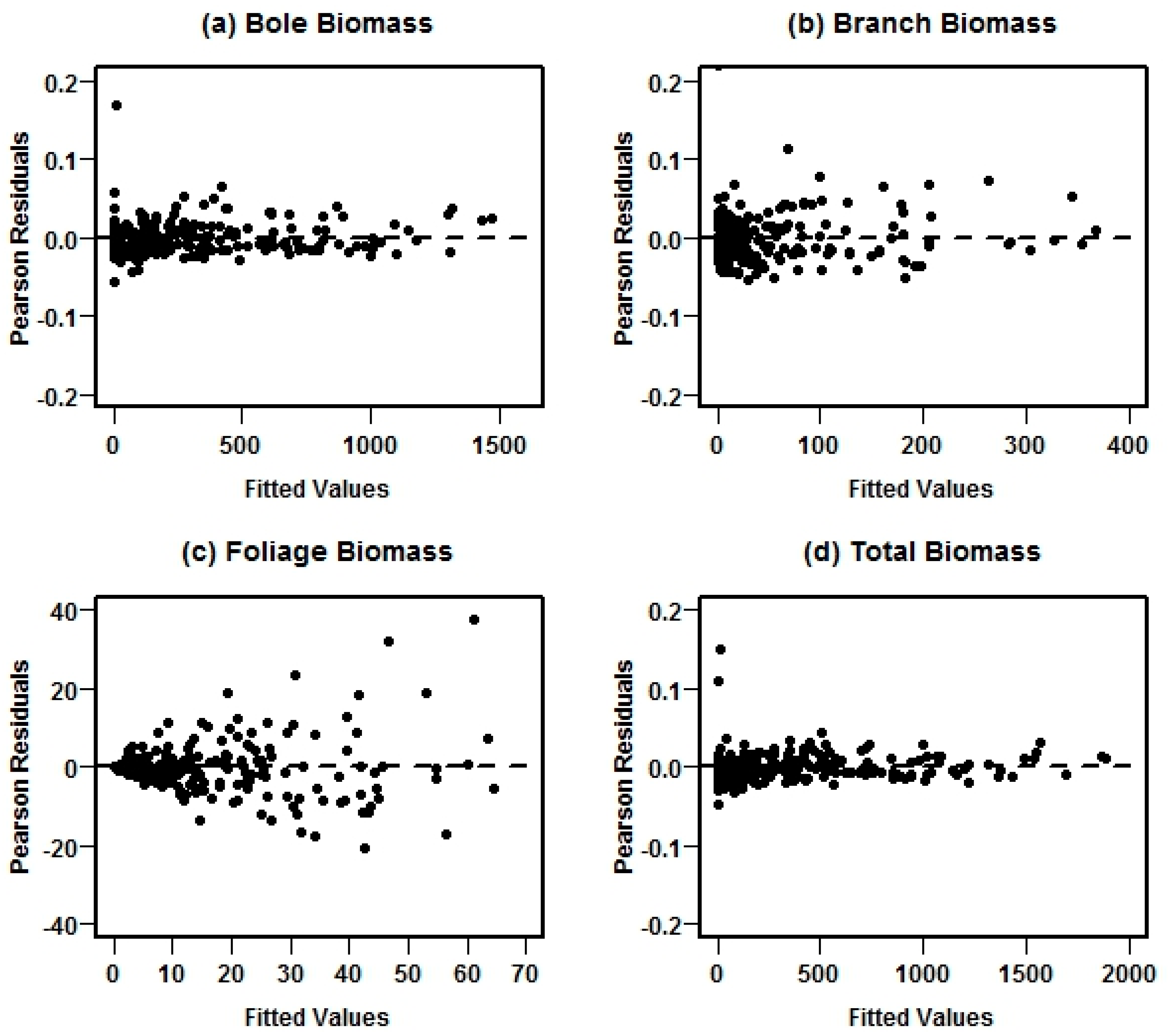
19] used in this study for comparison are not additive, as they were fitted separately for each component and total biomass. This model structure was used to ensure the additivity property of nonlinear models. The four-step fitting approach used in this study improved model estimation, increasing accuracy and precision of biomass predictions. Furthermore, the estimated weighting function eliminated heteroscedasticity for each of the biomass functions.
The models that estimated foliage biomass showed little improvement when HT (model system II) and/or tree age were included. Similar response was observed by Gonzalez-Benecke et al. [
10] for
Pinus taeda L. and
P. elliottii Engelm. trees. Compared with system I (based on DBH only), system III additionally included tree age which was significant only in the bole biomass equation, but also improved the branch and total biomass predictions. This response may be related to changes in wood density as trees age [
9,
22] and the interdependence of additive models. When A was included in the models that used DBH and HT (model system IV), it was significant in bole, branch and foliage equations, improving bole, branch and total biomass predictions. Although BA, rather than A, was significant in the branch biomass equation in system VI, that system did not predict component and total biomass better than system IV. When BA was significant in model systems V and VI, the parameter estimate for BA for foliage and branch was always negative, implying that for the same stem size and age, trees growing in stands with more intraspecific competition had smaller crown biomass than trees growing in stands with less intraspecific competition.
Even though the functions reported in this study can be used to estimate above-ground biomass directly, these equations have potential use in a variety of management and ecological applications. Several forest ecologists have used allometric biomass equations to quantify processes in forest ecosystems, such as nutrient dynamics, carbon sequestration, growth and competition (e.g., [
10,
23,
24,
25,
26,
27]), and better biomass predictions would find practical applications in nascent carbon-trading markets [
28]. Our new biomass equations should improve these types of analyses in longleaf pine forest ecosystems. Process-based models, such as 3-PG [
29] will also benefit from these more accurate and adaptable functions. Modeling systems that incorporate these equations can also be used to explore the effects of variable tree-level allometry on stand dynamics [
30].
All previously reported, biomass functions for longleaf pine trees rely on DBH only [
7,
15,
19] or DBH and HT [
8,
9,
15,
16,
17,
19]. When compared with our model system VI, the functions of Baldwin and Saucier [
8] and Samuelson et al. [
19] showed a tendency to estimate biased results. In addition to more accurate estimations, our models provide the additivity property to the biomass determinations.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}