Development of Somatic Embryo Maturation and Growing Techniques of Norway Spruce Emblings towards Large-Scale Field Testing


Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of Embryogenic Lines
2.2. Experiment I
2.3. Experiment II
2.4. Experiment III
2.4.1. Thawing, Proliferation Maturation and Germination
2.4.2. Embling Production
2.5. Measurements and Data Analysis
3. Results
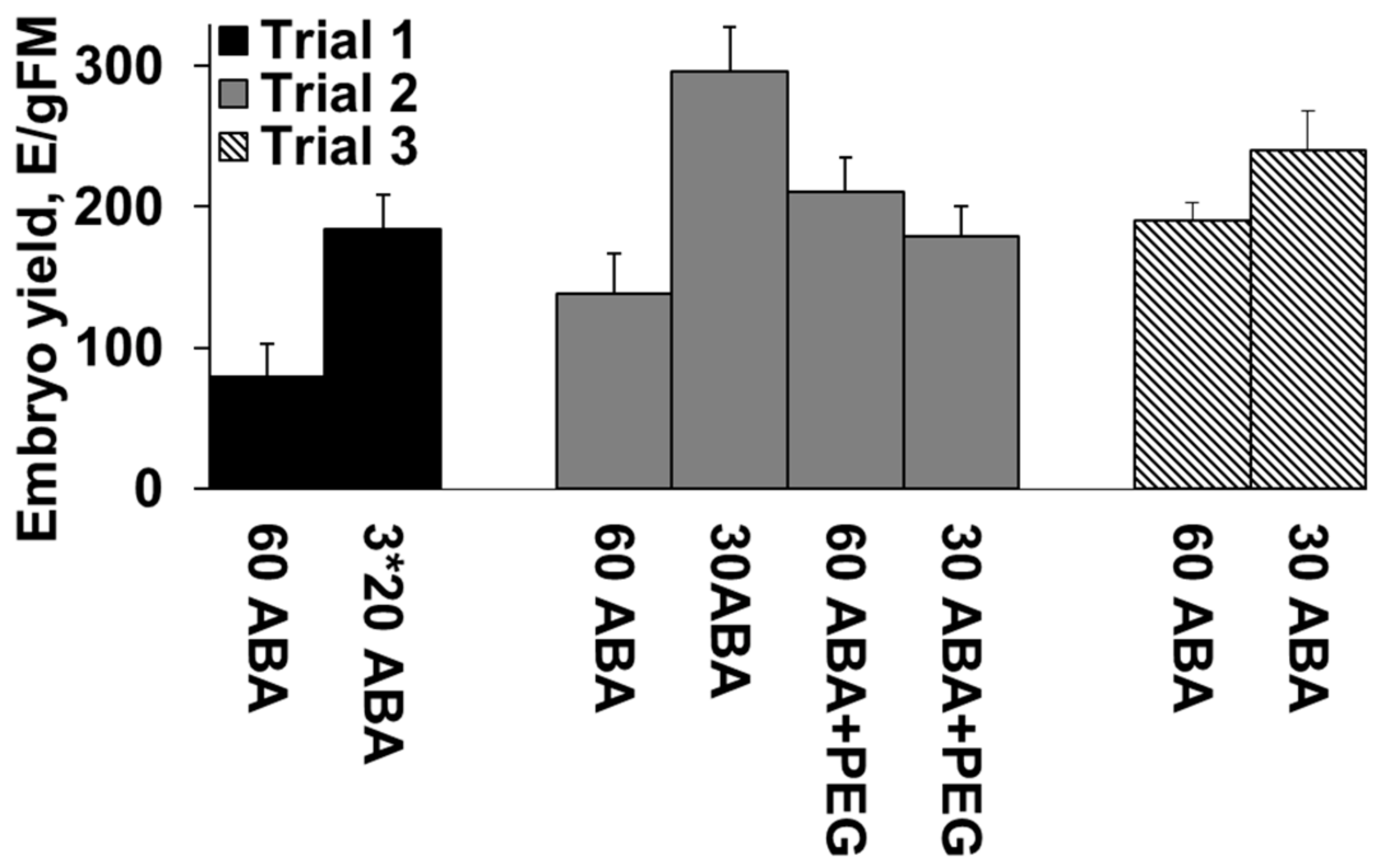
3.1. Effect of ABA Concentration and PEG in the Maturation Media
3.2. In Vitro Germination, Survival and Height Growth
3.3. Donor Plant Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Haapanen, M.; Leinonen, H.; Leinonen, K. Männyn ja kuusen siemenviljelyssiemenen taimitarhakäytön kehitys 2006–2016: Alueellinen tarkastelu. Metsätieteen Aikakauskirja 2017. [Google Scholar] [CrossRef]
- Chalupa, V. Somatic embryogenesis and plantlet regeneration from cultured immature and mature embryos of Picea abies (L.) Karst. Commun. Inst. For. Czech Repub. 1985, 14, 57–63. [Google Scholar]
- Hakman, I.; Fowke, L.C.; von Arnold, S.; Eriksson, T. The development of somatic embryos in tissue cultures initiated from immature embryos of Picea abies (Norway spruce). Plant Sci. 1985, 38, 53–59. [Google Scholar] [CrossRef]
- Bonga, J.M. Conifer clonal propagation in tree improvement programs. In Vegetative Propagation of Forest Trees; Park, Y.-S., Bonga, J.M., Moon, H.-K., Eds.; Korea Forest Research Institute: Seoul, Korea, 2016; pp. 3–31. [Google Scholar]
- Burdon, R.D.; Aimers-Halliday, Y.J. Risk management for clonal forestry with Pinus radiate–Analysis and review 1: Strategic issues and risk spread. N. Z. J. For. Sci. 2003, 33, 156–180. [Google Scholar]
- Grossnickle, S.C.; Cyr, D.; Polonenko, D.R. Somatic embryogenesis tissue culture for the propagation of conifer seedlings: A technology comes of age. Tree Plant. Notes 1996, 47, 48–57. [Google Scholar]
- Högberg, K.-A.; Bozhkov, P.V.; Grönroos, R.; von Arnold, S. Critical Factors Affecting Ex Vitro Performance of Somatic Embryo Plants of Picea abies. Scand. J. For. Res. 2001, 16, 295–304. [Google Scholar] [CrossRef]
- Varis, S.; Ahola, S.; Jaakola, L.; Aronen, T. Reliable and practical methods for cryopreservation of embryogenic cultures and cold storage of somatic embryos of Norway spruce. Cryobiology 2017, 76, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Cyr, D.R.; Lazaroff, W.R.; Grimes, S.M.; Quan, G.; Bethune, T.D.; Dunstan, D.I.; Roberts, D.R. Cryopreservation of interior spruce (Picea glauca engelmannii complex) embryogenic cultures. Plant Cell Rep. 1994, 13, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Find, J.; Floto, F.; Krogstrup, P.; Møller, J.D.; Nørgaard, J.V.; Kristensen, M.M.H. Cryopreservation of an embryogenic suspension culture of Picea sitchensis and subsequent plant regeneration. Scand. J. For. Res. 1993, 8, 156–162. [Google Scholar] [CrossRef]
- Hazubska-Przybył, T.; Chmielarz, P.; Michalak, M.; Bojarczuk, K. Cryopreservation of embryogenic tissues of Picea omorika (Serbian spruce). Plant Cell Tissue Organ Cult. 2010, 102, 35–44. [Google Scholar] [CrossRef]
- Häggman, H.M.; Ryynänen, L.A.; Aronen, T.S.; Krajnakova, J. Cryopreservation of embryogenic cultures of Scots pine. Plant Cell Tissue Organ Cult. 1998, 54, 45–53. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Ward, C.; Cheliak, W. Cryopreservation and plant regeneration from embryogenic cultures of larch (larix X eurolepis) and black spruce (Picea mariana). J. Exp. Bot. 1992, 43, 73–79. [Google Scholar] [CrossRef]
- Kong, L.; von Aderkas, P. A novel method of cryopreservation without a cryoprotectant for immature somatic embryos of conifer. Plant Cell Tissue Organ Cult. 2011, 106, 115–125. [Google Scholar] [CrossRef]
- Nørgaard, J.V.; Duran, V.; Johnsen, Ø.; Krogstrup, P.; Baldursson, S.; von Arnold, S. Variations in cryotolerance of embryogenic Picea abies cell lines and the association to genetic, morphological, and physiological factors. Can. J. For. Res. 1993, 23, 2560–2567. [Google Scholar] [CrossRef]
- Adams, G.W.; Kunze, H.A.; McCartney, A.; Millican, S.; Park, Y.-S. An industrial perspective on the use of advanced reforestation stock technologies. In Vegetative Propagation of Forest Trees; Park, Y.-S., Bonga, J.M., Moon, H.-K., Eds.; Korea Forest Research Institute: Seoul, Korea, 2016; pp. 323–334. [Google Scholar]
- Lelu-Walter, M.-A.; Thompson, D.; Harvengt, L.; Sanchez, L.; Toribio, M.; Pâques, L.E. Somatic embryogenesis in forestry with a focus on Europe: State-of-the-art, benefits, challenges and future direction. Tree Genet. Genomes 2013, 9, 883–899. [Google Scholar] [CrossRef]
- Bonga, J.M. A comparative evaluation of the application of somatic embryogenesis, rooting of cuttings, and organogenesis of conifers. Can. J. For. Res. 2015, 45, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Carson, M.; Carson, S.; Te Riini, C. Successful Varietal Forestry with Radiata Pine in New Zealand. N. Z. J. For. 2015, 60, 8–11. [Google Scholar]
- Thompson, D. Challenges for the large-scale propagation of forest trees by somatic embryogenesis—A review. In Proceedings of the 3rd International Conference of the IUFRO Unit 2.09.02 on “Woody Plant Production Integrating Genetic and Vegetative Propagation Technologies”, Vitoria-Gasteiz, Spain, 8–12 September 2014; pp. 81–91. [Google Scholar]
- Högberg, K.-A.; Ekberg, I.; Norell, L.; von Arnold, S. Integration of somatic embryogenesis in a tree breeding programme: A case study with Picea abies. Can. J. For. Res. 1998, 28, 1536–1545. [Google Scholar] [CrossRef]
- Majada, J.P.; Sierra, M.I.; Sanchez-Tames, R. Air exchange rate affects the in vitro developed leaf cuticle of carnation. Sci. Hortic. 2001, 87, 121–130. [Google Scholar] [CrossRef]
- Tikkinen, M.; Varis, S.; Peltola, H.; Aronen, T. Improved germination conditions for Norway spruce somatic cotyledonary embryos increased survival and height growth of emblings. Trees. submitted.
- Lelu, M.-A.; Bastien, C.; Klimaszewska, K.; Ward, C.; Charest, P.J. An improved method for somatic plantlet production in hybrid larch (Larix x leptoeuropaea). Part 1. Somatic embryo maturation. Plant Cell Tissue Organ Cult. 1994, 36, 107–115. [Google Scholar] [CrossRef]
- Find, J. Changes in endogenous ABA levels in developing somatic embryos of Norway spruce (Picea abies (L.) Karst.) in relation to maturation medium, desiccation and germination. Plant Sci. 1997, 128, 78–83. [Google Scholar] [CrossRef]
- Von Arnold, S.; Hakman, I. Regulation of somatic embryo development in Picea abies by abscisic acid (ABA). J. Plant Physiol. 1988, 132, 164–169. [Google Scholar] [CrossRef]
- Gutmann, M.; von Aderkas, P.; Label, P.; Lelu, M.-A. Effects of abscisic acid on somatic embryo maturation of hybrid larch. J. Exp. Bot. 1996, 47, 1905–1917. [Google Scholar] [CrossRef] [Green Version]
- Lelu, M.-A.; Bastien, C.; Klimaszewska, K.; Ward, C.; Charest, P.J. An improved method for somatic plantlet production in hybrid larch (Larix x leptoeuropaea). Part 2. Control of germination and plantlet development. Plant Cell Tissue Organ Cult. 1994, 36, 117–127. [Google Scholar] [CrossRef]
- Lelu, M.-A.; Label, P. Changes in the levels of abscisic acid and its glucose ester conjugate during maturation of hybrid larch (Larix x leptoeuropaea) somatic embryos, in relation to germination and plantlet recovery. Physiol. Plant. 1994, 92, 53–60. [Google Scholar] [CrossRef]
- Attree, S.M.; Fowke, L.C. Embryogeny of gymnosperms: Advances in synthetic seed technology of conifer plants. Plant Cell Tissue Organ Cult. 1993, 35, 1–35. [Google Scholar] [CrossRef]
- Attree, S.M.; Pomeroy, M.K.; Fowke, L.C. Development of white spruce (Picea glauca (Moench.) Voss) somatic embryos during culture with abscisic acid and osmoticum, and their tolerance to drying and frozen storage. J. Exp. Bot. 1995, 285, 433–439. [Google Scholar] [CrossRef]
- Kong, L.; Yeung, E.C. Effects of silver nitrate and polyethylene glycol on white spruce (Picea glauca) somatic embryo development: Enhancing cotyledonary embryo formation and endogenous ABA content. Physiol. Plant. 1995, 93, 298–304. [Google Scholar] [CrossRef]
- Merkle, S.A.; Montello, P.M.; Reece, H.M.; Kong, L. Somatic embryogenesis and cryostorage of eastern hemlock and Carolina hemlock for conservation and restoration. Trees 1994, 28, 1767–1776. [Google Scholar] [CrossRef]
- Attree, S.M.; Pomeroy, M.K.; Fowke, L.C. Manipulation of conditions for the culture of somatic embryos of white spruce for improved triacylglycerol biosynthesis and desiccation tolerance. Planta 1992, 198, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Bozhkov, P.V.; von Arnold, S. Polyethylene glycol promotes maturation but inhibits further development of Picea abies somatic embryos. Physiol. Plant. 1998, 104, 211–224. [Google Scholar] [CrossRef]
- Svobodová, H.; Albrechtová, J.; Kumstýřová, L.; Lipavská, H.; Vágner, M.; Vondráková, Z. Somatic embryogenesis in Norway spruce: Anatomical study of embryo development and influence of polyethylene glycol on maturation process. Plant Physiol. Biochem. 1999, 37, 209–221. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Lachance, D.; Pelletier, G.; Lelu, A.-M.; Seguin, A. Regeneration of transgenic Picea glauca, P. mariana, and P. abies after cocultivation of embryogenic tissue with Agrobacterium tumefaciens. In Vitro Cell. Dev. Biol. Plant 2001, 37, 748–755. [Google Scholar]
- Litvay, J.D.; Verma, D.C.; Johnson, M.A. Influence of loblolly pine (Pinus taeda L.) culture medium and its components on growth and somatic embryogenesis of wild carrot (Daucus carota L.). Plant Cell Rep. 1985, 4, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, M.F.; Yeung, E.C. The effects of reduced and oxidized glutathione on white spruce somatic embryogenesis. In Vitro Cell. Dev. Biol. Plant 2004, 40, 61–66. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Park, Y.-S.; Overton, C.; Maceacheron, I.; Bonga, J.M. Optimized somatic embryogenesis in Pinus strobus L. In Vitro Cell. Dev. Biol. Plant 2001, 37, 392–399. [Google Scholar] [CrossRef]
- Landis, T.D.; Dumroese, R.K.; Haase, D. The Container Tree Nursery Manual. Vol. 6. Seedling Propagation; Agricultural Handbook 674; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2010; pp. 3–164.
- Rikala, R. Metsäpuiden Paakkutaimien Kasvatusopas (Container Seedling Growing Manual for Forest Trees); The Finnish Forest Research Institute: Suonenjoki, Finland, 2012; 247 p.
- Nygren, M. Metsäpuiden Sisemenopas (Seedling Guide for Forest Trees); Metsäntutkimuslaitoksen Tiedonantoja: Suonenjoki, Finland, 2003; Volume 882, 144 p. [Google Scholar]
- Högberg, K.-A. Possibilities and limitations of vegetative propagation of Norway spruce. In Acta Universitatis Agriculturae Sueciae, Silvestria 294; Swedish University of Agricultural Sciences: Uppsala, Switzerland, 2003. [Google Scholar]
- Von Aderkas, P.; Kong, L.; Prior, N.A. In Vitro techniques for conifer embryogenesis. In Vegetative Propagation of Forest Trees; Park, Y.-S., Bonga, J.M., Moon, H.-K., Eds.; Korea Forest Research Institute: Seoul, Korea, 2016; pp. 335–350. [Google Scholar]
- Tikkinen, M.; Varis, S.; Peltola, H.; Aronen, T. Norway spruce Emblings as Cutting Donors for Tree Breeding and Production. Scand. J. For. Res. 2017, 33, 207–214. [Google Scholar] [CrossRef]
- Haapanen, M. Clones in Finnish tree breeding. In Proceedings of the Nordic Meeting Held in September 10th–11th 2008 at Punkaharju, Finland; Working papers of the Finnish Forest Research Institute; Finnish Forest Research Institute: Vantaa, Finland, 2009; Volume 114, pp. 16–19. [Google Scholar]


{kind=link}
| Exp./Trial | Treatment | Maturation | Germination | 1st Growing Period/Growing Season | 2nd Growing Season |
|---|---|---|---|---|---|
| I/1; 2; 3 | 60ABA | 8 weeks | |||
| I/1 | 3* 20ABA | 8 weeks | |||
| I/2 | 60ABA + PEG | 8 weeks | |||
| I/2 | 30ABA + PEG | 8 weeks | |||
| I/2; 3 | 30ABA | 8 weeks | |||
| II | GT-I | 8 weeks | 18:6 Day-night | Transplanted to Miniplugs | Grown outside since |
| 60 µM | 1 week (1) | Controlled environment | June 2017 | ||
| 30 µM ABA | Transplanted to Plantek 81f | ||||
| Winterized and cold stored | |||||
| GT-II | 8 weeks | 18:6 Day-night | Transplanted to Plantek 81f | ||
| 60 µM | 1 week (1) | Nursery greenhouse in March 2017 | |||
| 30 µM ABA | Grown outside since June 2017 | ||||
| III | Thawing lots A to D | 8 weeks | 18:6 Day-night | Transplanted to Miniplugs | Grown outside since |
| 60 µM ABA | 1 week (1) | Controlled environment | June 2017 | ||
| Winterized and cold stored | |||||
| Thawing lot E | 8 weeks | 18:6 Day-night | Transplanted to Miniplugs | ||
| 30 µM ABA | 1 week (1) | Controlled environment | |||
| Transplanted to Plantek 81f in March 2017 | |||||
| Grown outside since June 2017 | |||||
| Thawing lot F | 8 weeks | 18:6 Day-night | Transplanted to Plantek 81f | ||
| 30 µM ABA | 1 week (1) | Nursery greenhouse in March 2017 | |||
| Grown outside since June 2017 |
| Crossing | Explants | Initiated | Cryostored | Thawing | Thawed | Cryo + Maturation | E/gFW | Over 200 E/gFW | Surv. % A to D | Surv. % E to F |
|---|---|---|---|---|---|---|---|---|---|---|
| E1551 × E2229 | 99 | 67 | 61 | III, IV and VI | 48 | 45 | 94 (±13) | 7 | 28 (±2) | 72 (±2) |
| E162 × E81 | 200 | 101 | 66 | II and VI | 48 | 42 | 89 (±12) | 5 | 14 (±2) | 87 (±2) |
| E18 × E436 | 200 | 133 | 127 | III and VI | 47 | 30 | 110 (±18) | 6 | 62 (±2) | 85 (±2) |
| E207 × E1373 | 200 | 145 | 140 | III, IV and VI | 50 | 36 | 81 (±11) | 3 | 54 (±2) | 70 (±5) |
| E207 × E252 | 200 | 60 | 52 | II, IV and VI | 51 | 43 | 124 (±14) | 6 | 30 (±2) | 78 (±2) |
| E2105c × E2283 | 200 | 114 | 98 | II, IV and VI | 50 | 45 | 40 (±9) | 1 | 14 (±2) | 66 (±5) |
| E212 × E54 | 200 | 127 | 92 | I, IV and V | 92 | 61 | 45 (±7) | 2 | 14 (±2) | 80 (±2) |
| E242 × E222 | 400 | 187 | 106 | I, IV, V and VI | 106 | 49 | 36 (±7) | 2 | 10 (±2) | 82 (±2) |
| E46 × E3222 | 400 | 244 | 209 | III and VI | 78 | 53 | 89 (±12) | 5 | 25 (±2) | 88 (±2) |
| E462 × E64 | 200 | 160 | 137 | I and VI | 48 | 41 | 81 (±13) | 5 | 54 (±2) | 81 (±1) |
| E799 × E1366 | 45 | 42 | 36 | II and IV | 34 | 32 | 144 (±21) | 7 | 24 (±2) | |
| E9 × E3231 | 400 | 132 | 123 | I and VI | 60 | 52 | 68 (±10) | 5 | 5 (±1) | 71 (±2) |
| Overall | 2744 | 1512 | 1247 | I to VI | 712 | 529 | 79 (±4) | 54 | 30 (±1) | 80 (±1) |
| Experiment | Measurement | Model | Variable | Sig. | Odds Ratio (95% Confidence Interval) | Correct, % | |
|---|---|---|---|---|---|---|---|
| Experiment II | After GP1 | log(p/1−p) = | 2.839 − 1.372g1 − 0.076c2 − 0.437c4 − 0.748c6 − 0.820c7 + 0.470c8 − 0.748c9 − 0.163c10 − 0.546c11 | 81 | |||
| Growing technique | <0.001 | 1 | GT-I | ||||
| 0.254 (0.177–0.363) | GT-II | ||||||
| Family | <0.001 | 1 | E1551 × E2229 | ||||
| 0.927 (0.591–1.453) | E18 × E436 | ||||||
| 0.623 (0.341–1.138) | E207 × E252 | ||||||
| 0.473 (0.293–0.765) | E212 × E54 | ||||||
| 0.44 (0.25–0.775) | E242 × E222 | ||||||
| 0.473 (0.293–0.765) | E462 × E64 | ||||||
| 1.6 (0.956–2.676) | E46 × E3222 | ||||||
| 0.849 (0.509–1.419) | E799 × E1366 | ||||||
| 0.579 (0.324–1.035) | E9 × E3231 | ||||||
| Experiment II | Autumn 2017 | log(p/1−p) = | 0.386 + 1.210g1 + 0.150c2 − 0.437c4 − 0.588c6 − 0.654c7 + 0.491c8 − 1.248c9 − 0.899c10 − 1.095c11 | 74.1 | |||
| Growing technique | <0.001 | 1 | GT-I | ||||
| 3.354 (2.684–4.191) | GT-II | ||||||
| Family | <0.001 | 1 | E1551 × E2229 | ||||
| 1.162 (0.785–1.72) | E18 × E436 | ||||||
| 0.646 (0.368–1.134) | E207 × E252 | ||||||
| 0.556 (0.361–0.855) | E212 × E54 | ||||||
| 0.52 (0.31–0.873) | E242 × E222 | ||||||
| 0.287 (0.188–0.437) | E462 × E64 | ||||||
| 1.634 (1.06–2.52) | E46 × E3222 | ||||||
| 0.407 (0.266–0.622) | E799 × E1366 | ||||||
| 0.335 (0.203–0.55) | E9 × E3231 | ||||||
| Experiment III | After GP1 | log(p/1−p) = | 0.859 + 0.238c1 + 0.945c2 + 0.421c3 + 0.018c4 + 0.104c5 + 0.405c6 + 0.419c7 + 1.206c8 + 0.651c9 + 0.574c10 − 0.003c11 | 78 | |||
| Family | <0.001 | 1 | E1551 × E2229 | ||||
| 1.269 (1.019–1.58) | E162 × E81 | ||||||
| 2.573 (2.035–3.252) | E18 × E436 | ||||||
| 1.523 (1.188–1.954) | E207 × E1373 | ||||||
| 1.018 (0.83–1.25) | E207 × E252 | ||||||
| 1.11 (0.832–1.48) | E2105c × E2283 | ||||||
| 1.499 (1.183–1.899) | E212 × E54 | ||||||
| 1.52 (1.197–1.931) | E242 × E222 | ||||||
| 3.34 (2.601–4.289) | E462 × E64 | ||||||
| 1.918 (1.575–2.336) | E46 × E3222 | ||||||
| 1.775 (1.371–2.298) | E799 × E1366 | ||||||
| 0.997 (0.812–1.225) | E9 × E3231 | ||||||
| Experiment III | Autumn 2017 | log(p/1−p) = | −2.652 + 0.613b1 + 1.186b2 + 0.915b3 + 3.753b4 + 3.334b5 + 0.632c1 + 1.186c2 + 0.886c3 + 0.912c4 + 0.108c5 + 0.365c6 + 0.401c7 + 1.494c8 + 0.831c9 + 0.840c10 + 0.126c11 | 76.5 | |||
| Thawing | <0.001 | 1 | A | ||||
| 1.846 (1.326–2.571) | B | ||||||
| 6.66 (4.935–8.988) | C | ||||||
| 2.497 (1.714–3.638) | D | ||||||
| 42.655 (29.934–60.782) | E | ||||||
| 28.049 (21.653–36.334) | F | ||||||
| Family | <0.001 | 1 | E1551 × E2229 | ||||
| 1.881 (1.427–2.479) | E162 × E81 | ||||||
| 3.275 (2.659–4.034) | E18 × E436 | ||||||
| 2.425 (1.931–3.046) | E207 × E1373 | ||||||
| 2.49 (1.907–3.25) | E207 × E252 | ||||||
| 1.114 (0.776–1.601) | E2105c × E2283 | ||||||
| 1.441 (0.973–2.135) | E212 × E54 | ||||||
| 1.493 (1.049–2.123) | E242 × E222 | ||||||
| 4.456 (3.266–6.08) | E462 × E64 | ||||||
| 2.297 (1.905–2.769) | E46 × E3222 | ||||||
| 2.316 (1.659–3.232) | E799 × E1366 | ||||||
| 1.134 (0.88–1.462) | E9 × E3231 | ||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tikkinen, M.; Varis, S.; Aronen, T. Development of Somatic Embryo Maturation and Growing Techniques of Norway Spruce Emblings towards Large-Scale Field Testing. Forests 2018, 9, 325. https://doi.org/10.3390/f9060325
Tikkinen M, Varis S, Aronen T. Development of Somatic Embryo Maturation and Growing Techniques of Norway Spruce Emblings towards Large-Scale Field Testing. Forests. 2018; 9(6):325. https://doi.org/10.3390/f9060325
Chicago/Turabian StyleTikkinen, Mikko, Saila Varis, and Tuija Aronen. 2018. "Development of Somatic Embryo Maturation and Growing Techniques of Norway Spruce Emblings towards Large-Scale Field Testing" Forests 9, no. 6: 325. https://doi.org/10.3390/f9060325
APA StyleTikkinen, M., Varis, S., & Aronen, T. (2018). Development of Somatic Embryo Maturation and Growing Techniques of Norway Spruce Emblings towards Large-Scale Field Testing. Forests, 9(6), 325. https://doi.org/10.3390/f9060325