Forest Regeneration Following Emerald Ash Borer (Agrilus planipennis Fairemaire) Enhances Mesophication in Eastern Hardwood Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Data Collection
2.3. Analyses
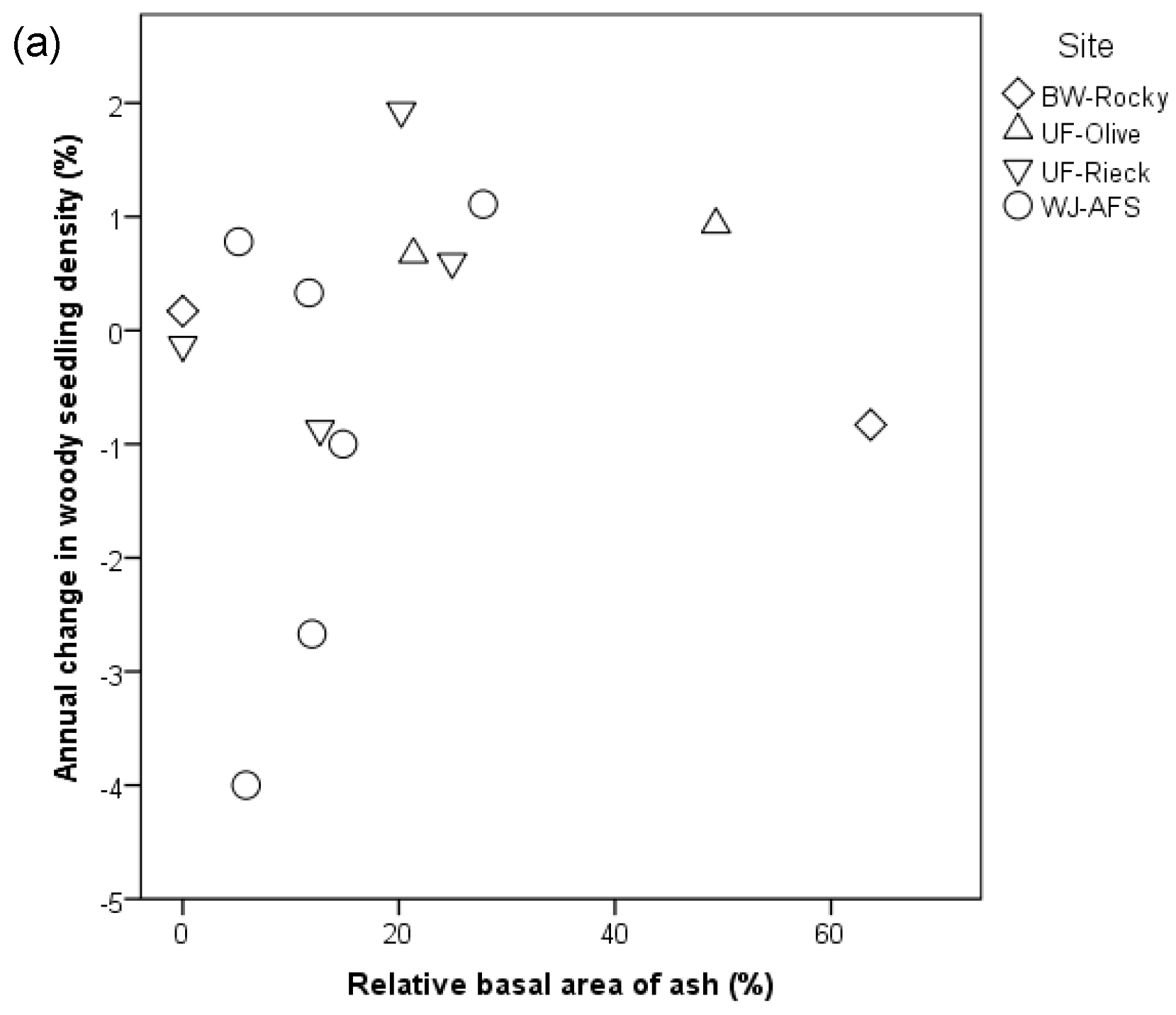
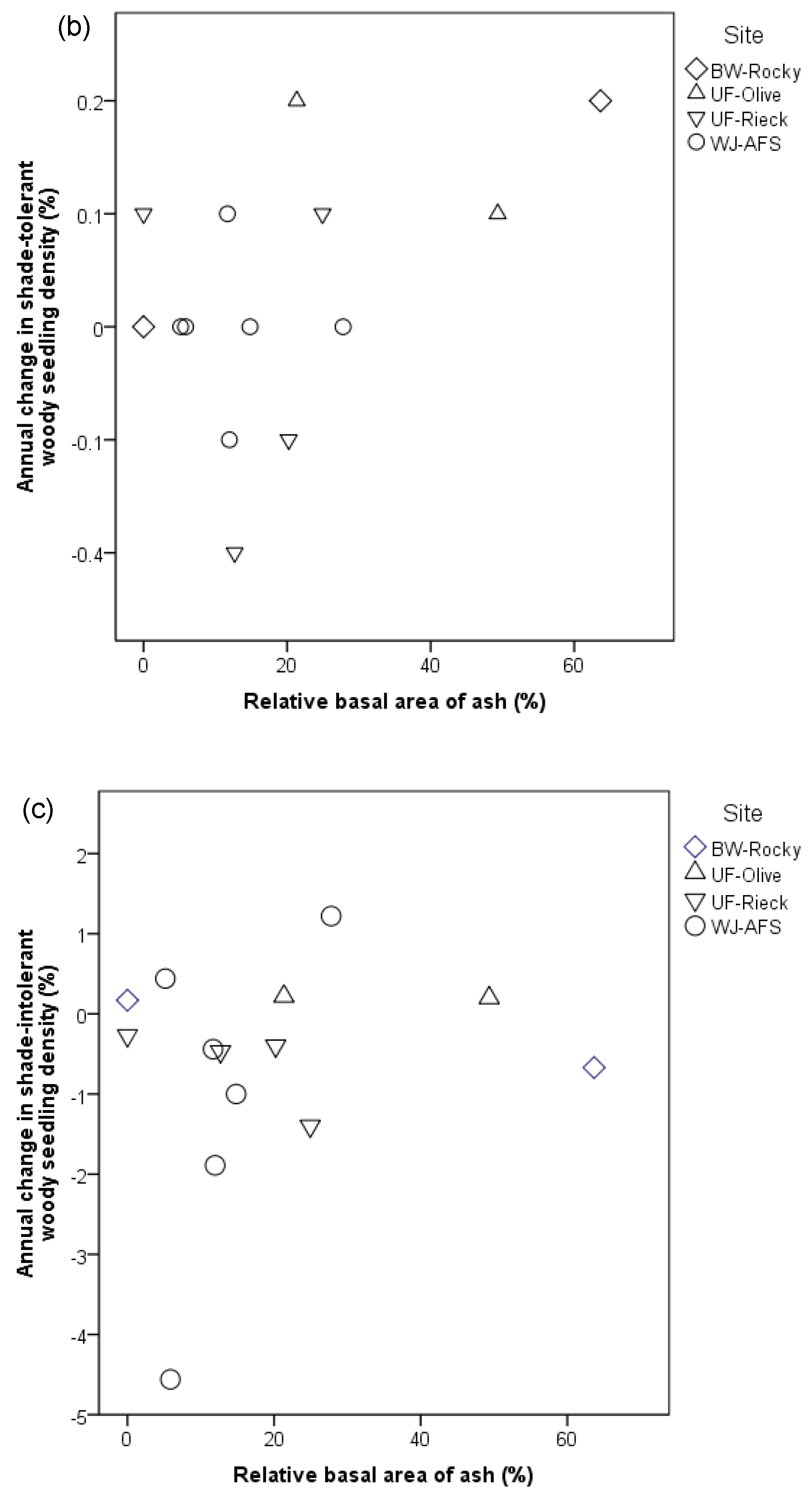
3. Results
4. Discussion
4.1. Canopy Cover
4.2. Understory Composition and Density
4.3. Invasive Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haack, R.A.; Jendak, E.; Houping, L.; Marchant, K.R.; Petrice, T.R.; Poland, T.M.; Ye, H. The emerald ash borer: A new exotic pest in North America. Newsl. Mich. Entomological Soc. 2002, 47, 1–5. [Google Scholar]
- Poland, T.M.; McCullough, D.G. Emerald Ash Borer: Invasion of the Urban Forest and the Threat to North America’s Ash Resource. J. For. 2006, 104, 118–124. [Google Scholar] [CrossRef]
- Animal and Plant Health Inspection Service, USDA. Emerald Ash Borer: Quarantine and Regulations. Available online: https://www.federalregister.gov/d/03-25881 (accessed on 15 April 2018).
- Herms, D.A.; McCullough, D.G. Emerald Ash Borer Invasion of North America: History, Biology, Ecology, Impacts, and Management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Perry, K.I.; Herms, D.A.; Klooster, W.S.; Smith, A.; Hartzler, D.M.; Coyle, D.R.; Gandhi, K.J.K. Downed Coarse Woody Debris Dynamics in Ash (Fraxinus spp.) Stands Invaded by Emerald Ash Borer (Agrilus planipennis Fairmaire). Forests 2018, 9, 191. [Google Scholar] [CrossRef]
- Ricketts, M.P.; Flower, C.E.; Knight, K.S.; Gonzalez-Meler, M.A. Evidence of Ash Tree (Fraxinus spp.) Specific Associations with Soil Bacterial Community Structure and Functional Capacity. Forests 2018, 9, 187. [Google Scholar] [CrossRef]
- Marché, J.D., II. The Green Menace: Emerald Ash Borer and the Invasive Species Problem; Oxford University Press: Oxford, UK, 2017; ISBN 978-0-19-066892-1. [Google Scholar]
- Flower, C.E.; Knight, K.S.; Gonzalez-Meler, M.A. Impacts of the emerald ash borer (Agrilus planipennis Fairmaire) induced ash (Fraxinus spp.) mortality on forest carbon cycling and successional dynamics in the eastern United States. Biol. Invasions 2013, 15, 931–944. [Google Scholar] [CrossRef]
- Hoven, B.M.; Gorchov, D.L.; Knight, K.S.; Peters, V.E. The effect of emerald ash borer-caused tree mortality on the invasive shrub Amur honeysuckle and their combined effects on tree and shrub seedlings. Biol. Invasions 2017, 19, 2813–2836. [Google Scholar] [CrossRef]
- McCullough, D.G.; Siegert, N.W. Estimating potential emerald ash borer (Coleoptera: Buprestidae) populations using ash inventory data. J. Econ. Entomol. 2007, 100, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Vankat, J.L. A classification of the forest types of North America. Vegetatio 1990, 88, 53–66. [Google Scholar] [CrossRef]
- MacFarlane, D.W.; Meyer, S.P. Characteristics and distribution of potential ash tree hosts for emerald ash borer. For. Ecol. Manag. 2005, 213, 15–24. [Google Scholar] [CrossRef]
- Anagnostakis, S.L. Chestnut blight: The classical problem of an introduced pathogen. Mycologia 1987, 79, 23–37. [Google Scholar] [CrossRef]
- Karnosky, D.F. Dutch elm disease: A review of the history, environmental implications, control, and research needs. Environ. Conserv. 1979, 6, 311–322. [Google Scholar] [CrossRef]
- Morin, R.S.; Liebhold, A.M. Invasions by two non-native insects alter regional forest species composition and successional trajectories. For. Ecol. Manag. 2015, 341, 67–74. [Google Scholar] [CrossRef]
- Elliott, K.J.; Swank, W.T. Long-term changes in forest composition and diversity following early logging (1919–1923) and the decline of American chestnut (Castanea dentata). Plant Ecol. 2007, 197, 155–172. [Google Scholar] [CrossRef]
- Stephenson, S.L. Changes in a former chestnut-dominated forest after a half century of succession. Am. Midl. Nat. 1986, 116, 173–179. [Google Scholar] [CrossRef]
- Myers, B.R.; Walck, J.L.; Blum, K.E. Vegetation change in a former chestnut stand on the Cumberland Plateau of Tennessee during an 80-year period (1921–2000). Castanea 2004, 69, 81–91. [Google Scholar] [CrossRef]
- Barnes, B.V. Succession in deciduous swamp communities of southeastern Michigan formerly dominated by American elm. Can. J. Bot. 1976, 54, 19–24. [Google Scholar] [CrossRef]
- Parker, G.R.; Leopold, D.J. Replacement of Ulmus americana L. in a mature east-central Indiana woods. Bull. Torrey Bot. Club 1983, 110, 482. [Google Scholar] [CrossRef]
- Costilow, K.C.; Knight, K.S.; Flower, C.E. Disturbance severity and canopy position control the radial growth response of maple trees (Acer spp.) in forests of northwest Ohio impacted by emerald ash borer (Agrilus planipennis). Ann. For. Sci. 2017, 74, 10. [Google Scholar] [CrossRef]
- Burr, S.J.; McCullough, D.G. Condition of green ash (Fraxinus pennsylvanica) overstory and regeneration at three stages of the emerald ash borer invasion wave. Can. J. For. Res. 2014, 44, 768–776. [Google Scholar] [CrossRef]
- Kolka, R.K.; D’Amato, A.W.; Wagenbrenner, J.W.; Slesak, R.A.; Pypker, T.G.; Youngquist, M.B.; Grinde, A.R.; Palik, B.J. Review of ecosystem level impacts of emerald ash borer on black ash wetlands: What does the future hold? Forests 2018, 9, 179. [Google Scholar] [CrossRef]
- Iverson, L.; Knight, K.S.; Prasad, A.; Herms, D.A.; Matthews, S.; Peters, M.; Smith, A.; Hartzler, D.M.; Long, R.; Almendinger, J. Potential species replacements for black ash (Fraxinus nigra) at the confluence of two threats: Emerald ash borer and a changing climate. Ecosystems 2015, 19, 248–270. [Google Scholar] [CrossRef]
- Davis, J.C.; Shannon, J.P.; Bolton, N.W.; Kolka, R.K.; Pypker, T.G. Vegetation responses to simulated emerald ash borer infestation in Fraxinus nigra dominated wetlands of Upper Michigan, USA. Can. J. For. Res. 2017, 47, 319–330. [Google Scholar] [CrossRef]
- Looney, C.E.; D’Amato, A.W.; Palik, B.J.; Slesak, R.A. Canopy treatment influences growth of replacement tree species in Fraxinus nigra forests threatened by the emerald ash borer in Minnesota, USA. Can. J. For. Res. 2017, 47, 183–192. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. The demise of fire and “mesophication” of forests in the eastern United States. Bioscience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Alexander, H.D.; Arthur, M.A. Implications of a predicted shift from upland oaks to red maple on forest hydrology and nutrient availability. Can. J. For. Res. 2010, 40, 716–726. [Google Scholar] [CrossRef]
- Alexander, H.D.; Arthur, M.A. Increasing red maple leaf litter alters decomposition rates and nitrogen cycling in historically oak-dominated forests of the eastern U.S. Ecosystems 2014, 17, 1371–1383. [Google Scholar] [CrossRef]
- Kreye, J.K.; Varner, J.M.; Hiers, J.K.; Mola, J. Toward a mechanism for eastern North American forest mesophication: Differential litter drying across 17 species. Ecol. Appl. 2013, 23, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Tatina, R. Changes in Fagus grandifolia and Acer saccharum abundance in an old-growth, beech-maple forest at Warren Woods State Park, Berrien County, Michigan, USA. Castanea 2015, 80, 95–102. [Google Scholar] [CrossRef]
- Natural Resources Conservation Service, United States Department of Agriculture. Web Soil Survey. Available online: https://websoilsurvey.sc.egov.usda.gov/ (accessed on 7 July 2017).
- Initial County EAB Detections in North America 2017. Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/emerald_ash_b/downloads/MultiState.pdf (accessed on 11 February 2018).
- Kuers, K.; Lindquist, L. PFPP Protocols and Datasheets. Available online: http://erenweb.org/new-page/carbon-storage-project/permanent-plot-protocol/pfpp-protocol-files/ (accessed on 9 March 2018).
- Smith, A. Effects of community structure on forest susceptibility and response to the emerald ash borer invasion of the Huron River watershed in southeast Michigan. Master’s Thesis, The Ohio State University, Columbus, OH, USA, 2006. [Google Scholar]
- Knight, K.S.; Flash, B.P.; Kappler, R.H.; Throckmorton, J.A.; Grafton, B.; Flower, C.E. Monitoring Ash (Fraxinus spp.) Decline and Emerald Ash Borer (Agrilus planipennis) Symptoms in Infested Areas; General Technical Report NRS-139; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014; p. 18.
- Dolan, B.J.; Kilgore, J.S. Emerald Ash Borer Project. Available online: http://erenweb.org/new-page/eab/ (accessed on 11 February 2018).
- Kilgore, J.S.; Dolan, B.J. Complementary Vegetation Survey (cVeg). Available online: http://erenweb.org/new-page/cveg/ (accessed on 11 February 2018).
- Baker, F.S. A revised tolerance table. J. For. 1949, 47, 179–181. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Canham, C.D.; Denslow, J.S.; Platt, W.J.; Runkle, J.R.; Spies, T.A.; White, P.S. Light regimes beneath closed canopies and tree-fall gaps in temperate and tropical forests. Can. J. For. Res. 1990, 20, 620–631. [Google Scholar] [CrossRef]
- Gottschalk, KW. Shade, leaf growth and crown development of Quercus rubra, Quercus velutina, Prunus serotina and Acer rubrum seedlings. Tree Physiol. 1994, 14, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Parker, W.C.; Dey, D.C. Influence of overstory density on ecophysiology of red oak (Quercus rubra) and sugar maple (Acer saccharum) seedlings in central Ontario shelterwoods. Tree Physiol. 2008, 28, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Senécal, J.-F.; Doyon, F.; Messier, C. Tree death not resulting in gap creation: An investigation of canopy dynamics of northern temperate deciduous forests. Remote Sens. 2018, 10, 17. [Google Scholar] [CrossRef]
- Klooster, W.S.; Herms, D.A.; Knight, K.S.; Herms, C.P.; McCullough, D.G.; Smith, A.; Gandhi, K.J.K.; Cardina, J. Ash (Fraxinus spp.) mortality, regeneration, and seed bank dynamics in mixed hardwood forests following invasion by emerald ash borer (Agrilus planipennis). Biol. Invasions 2014, 16, 859–873. [Google Scholar] [CrossRef]
- Pušpure, I.; Laiviņš, M.; Matisons, R.; Gaitnieks, T. Understory changes in Fraxinus excelsior stands in response to dieback in Latvia. Proc. Latv. Acad. Sci. 2016, 70, 131–137. [Google Scholar] [CrossRef]
- Huenneke, L.F. Understory response to gaps caused by the death of Ulmus americana in central New York. Bull. Torrey Bot. Club 1983, 110, 170–175. [Google Scholar] [CrossRef]
- Dunn, C.P. Shrub layer response to death of Ulmus americana in southeastern Wisconsin lowland forests. Bull. Torrey Bot. Club 1986, 113, 142. [Google Scholar] [CrossRef]
- Poulson, T.L.; Platt, W.J. Replacement patterns of beech and sugar maple in Warren Woods, Michigan. Ecology 1996, 77, 1234–1253. [Google Scholar] [CrossRef]
- Yamamoto, S.-I. The gap theory in forest dynamics. Bot. Mag. Tokyo 1992, 105, 375–383. [Google Scholar] [CrossRef]
- Hart, J.L.; Grissino-Mayer, H.D. Gap-scale disturbance processes in secondary hardwood stands on the Cumberland Plateau, Tennessee, USA. Plant Ecol. 2008, 201, 131–146. [Google Scholar] [CrossRef]
- Cowell, C.M.; Mark Cowell, C.; Hoalst-Pullen, N.; Jackson, M.T. The limited role of canopy gaps in the successional dynamics of a mature mixed Quercus forest remnant. J. Veg. Sci. 2010, 21, 201–212. [Google Scholar] [CrossRef]
- Flatley, W.T.; Lafon, C.W.; Grissino-Mayer, H.D.; LaForest, L.B. Changing fire regimes and old-growth forest succession along a topographic gradient in the Great Smoky Mountains. For. Ecol. Manage. 2015, 350, 96–106. [Google Scholar] [CrossRef]
- Chapman, J.I.; McEwan, R.W. Thirty years of compositional change in an old-growth temperate forest: The role of topographic gradients in oak-maple dynamics. PLoS ONE 2016, 11, e0160238. [Google Scholar] [CrossRef] [PubMed]
- Knopp, P.D. The distribution of Quercus rubra in the Maumee Lake Plain of southeastern Michigan. Am. Midl. Nat. 2012, 168, 70–92. [Google Scholar] [CrossRef]
- Jerome, D.; Westwood, M.; Oldfield, S.; Romero-Severson, J. Fraxinus Americana. Available online: http://www.iucnredlist.org/details/61918430/0 (accessed on 11 February 2018).
- Westwood, M.; Oldfield, S.; Jerome, D.; Romero-Severson, J. Fraxinus quadrangulata. Available online: http://www.iucnredlist.org/details/61919112/0 (accessed on 11 February 2018).
- Jerome, D.; Westwood, M.; Oldfield, S.; Romero-Severson, J. Fraxinus nigra. Available online: http://www.iucnredlist.org/details/61918683/0 (accessed on 11 February 2018).
- Westwood, M.; Oldfield, S.; Jerome, D.; Romero-Severson, J. Fraxinus pennsylvanica. Available online: http://www.iucnredlist.org/details/61918934/0 (accessed on 11 February 2018).
- Westwood, M.; Jerome, D.; Oldfield, S.; Romero-Severson, J. Fraxinus profunda. Available online: http://www.iucnredlist.org/details/61919022/0 (accessed on 11 February 2018).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Institution | County | State | MGS (days) | MAP (mm) | MAT (°C) | Mean Elevation (m∙asl) | Predominant Soil Texture | Dominant Tree Species (in Order of Dominance) | Mean Plot Basal Area (m2∙ha−1) | Year of Detected EAB |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rocky River 41.411217° N, 81.881817° W | Baldwin-Wallace University (BW) | Cuyahoga | OH | 173 | 1132 | 11.67 | 233 | Silty loam | Fraxinus americana L., Ulmus rubra Muhl., Quercus rubra L., Carya glabra Miller, Acer saccharum Marshall | 36.0 | 2012 |
| Olive Street 41.002292° N, 83.642219° W | University of Findlay (UF) | Hancock | OH | 159 | 847 | 10.81 | 240 | Silty clay loam | Celtis occidentalis L., Acer saccharum, Fraxinus pennsylvanica Marshall, Carya cordiformis (Wangenh.) K.Koch, Juglans nigra L., Ulmus americana L. | 25.7 | 2010 |
| Rieck Center 40.950716° N, 83.549870° W | University of Findlay (UF) | Hancock | OH | 159 | 857 | 10.81 | 250 | Silty loam | Acer saccharum, Fraxinus pennsylvanica, Fagus grandifolia Ehrh., Ulmus americana | 31.9 | 2010 |
| Abernathy Field Station (AFS) 40.134110° N, 80.183625° W | Washington & Jefferson College (WJ) | Washington | PA | 169 | 970 | 9.6 | 367 | Silty loam | Prunus serotina Ehrh., Acer saccharum, Fraxinus americana, Carya ovata (Mill.) K.Koch | 37.1 | 2014 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dolan, B.; Kilgore, J. Forest Regeneration Following Emerald Ash Borer (Agrilus planipennis Fairemaire) Enhances Mesophication in Eastern Hardwood Forests. Forests 2018, 9, 353. https://doi.org/10.3390/f9060353
Dolan B, Kilgore J. Forest Regeneration Following Emerald Ash Borer (Agrilus planipennis Fairemaire) Enhances Mesophication in Eastern Hardwood Forests. Forests. 2018; 9(6):353. https://doi.org/10.3390/f9060353
Chicago/Turabian StyleDolan, Benjamin, and Jason Kilgore. 2018. "Forest Regeneration Following Emerald Ash Borer (Agrilus planipennis Fairemaire) Enhances Mesophication in Eastern Hardwood Forests" Forests 9, no. 6: 353. https://doi.org/10.3390/f9060353
APA StyleDolan, B., & Kilgore, J. (2018). Forest Regeneration Following Emerald Ash Borer (Agrilus planipennis Fairemaire) Enhances Mesophication in Eastern Hardwood Forests. Forests, 9(6), 353. https://doi.org/10.3390/f9060353