Vertical Transmission of Fusarium circinatum Mitoviruses FcMV1 and FcMV2-2 via Microconidia

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. F. circinatum Isolates
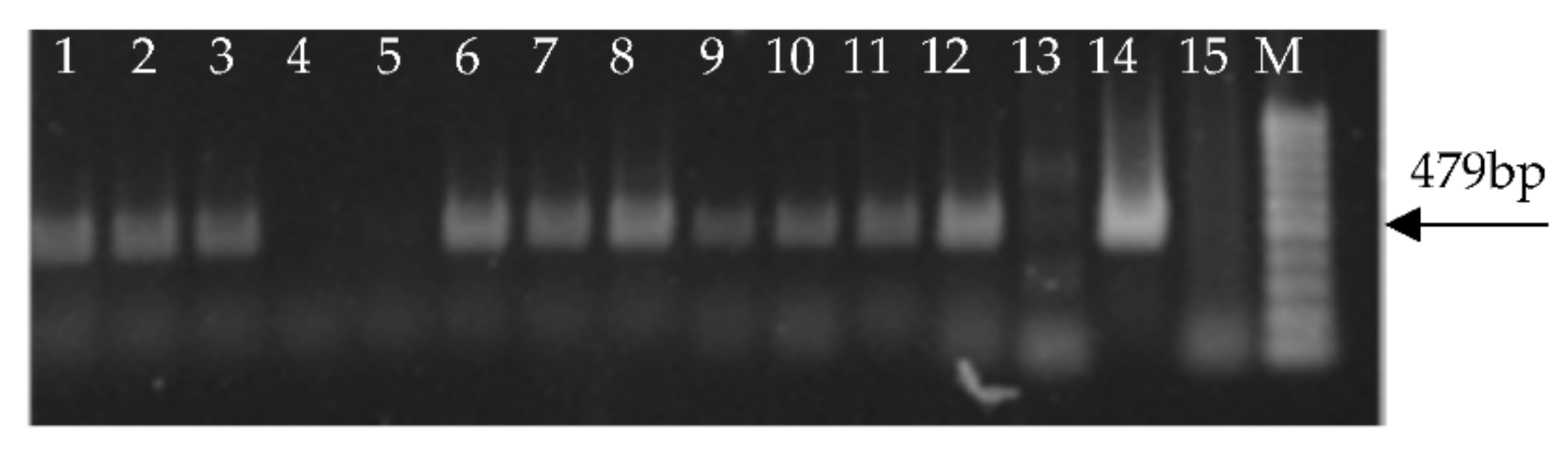
2.2. Total RNA Extraction and PCR
2.3. Statistical Analysis
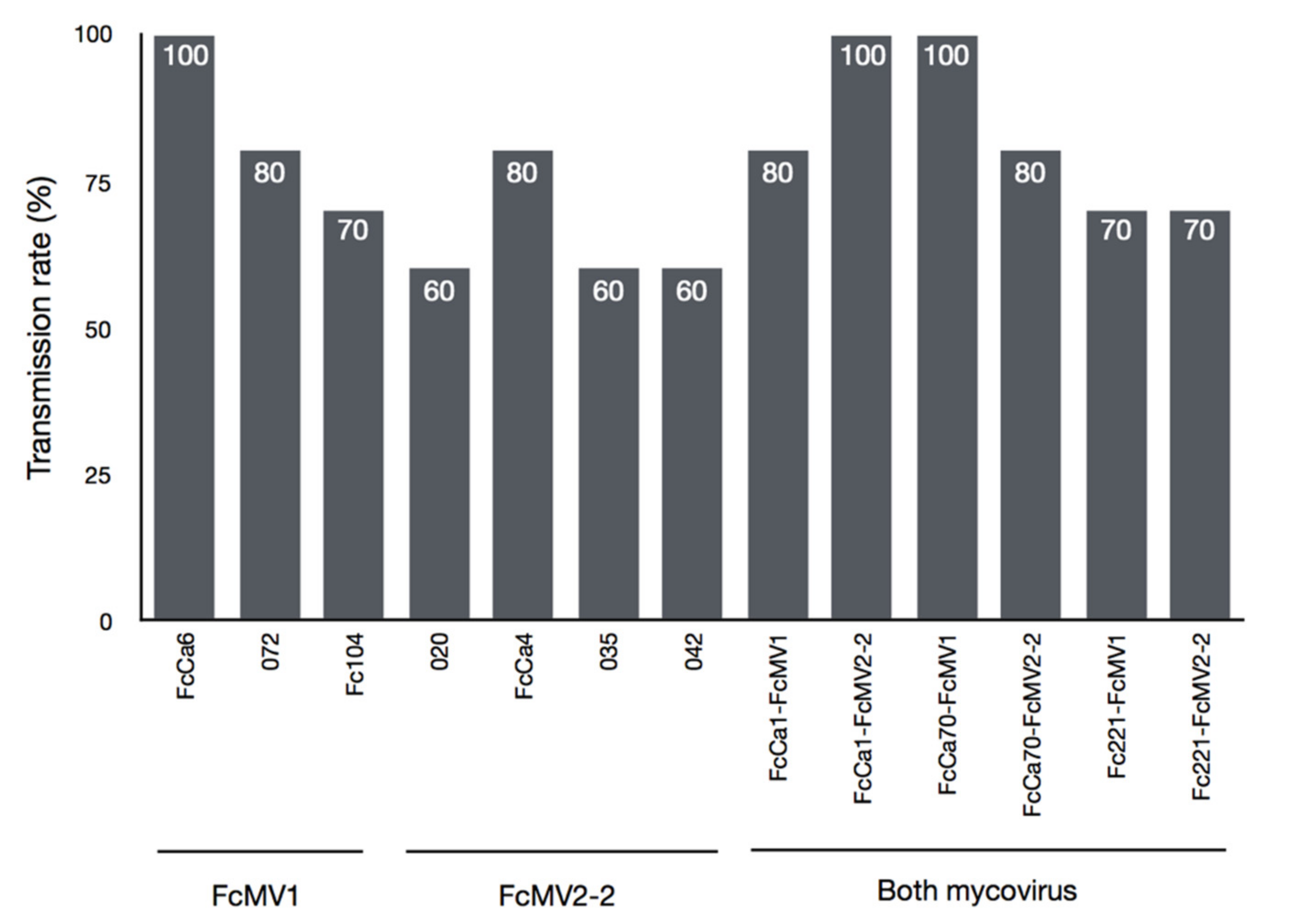
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martín-Rodrigues, N.; Espinel, S.; Sanchez-Zabala, J.; Ortíz, A.; González-Murua, C.; Duñabeitia, M.K. Spatial and temporal dynamics of the colonization of Pinus radiata by Fusarium circinatum, of conidiophora development in the pith and of traumatic resin duct formation. New Phytol. 2013, 198, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, M.J.; Jacobs, A.; Coutinho, T.A.; Ahumada, R.; Wingfield, B.D. First report of the pitch canker fungus, Fusarium circinatum, on pines in Chile. Plant Pathol. 2002, 51, 397. [Google Scholar] [CrossRef]
- Bezos, D.; Martínez-Alvarez, P.; Fernández, M.; Diez, J.J. Epidemiology and management of pine pitch canker disease in Europe—A review. Balt. For. 2017, 23, 279–293. [Google Scholar]
- Wingfield, M.J.; Hammerbacher, A.; Ganley, R.J.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, B.D.; Coutinho, T.A. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Aust. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- EU. Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009 Establishing a Framework for Community Action to Achieve the Sustainable Use of Pesticides; EU: Brussels, Belgium, 2009. [Google Scholar]
- Agrios, G.N. Plant Pathology, 4th ed.; Academic Press: San Diego, CA, USA, 1997; p. 635. [Google Scholar]
- Martínez-Álvarez, P.; Pando, V.; Diez, J.J. Alternative species to replace Monterey pine plantations affected by pitch canker caused by Fusarium circinatum in northern Spain. Plant Pathol. 2014, 63, 1086–1094. [Google Scholar] [CrossRef]
- Brimner, T.A.; Boland, G.J. A review of the non-target effects of fungi used to biologically control plant diseases. Agric. Ecosyst. Environ. 2003, 100, 3–16. [Google Scholar] [CrossRef]
- Ghabrial, S.A; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-Plus Years of Fungal Viruses. Virology 2015, 479, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Mccabe, P.M.; Pfeiffer, P.; Van Alfen, N.K. The influence of dsRNA viruses on the biology of plant pathogenic fungi. Trends Microbiol. 1999, 7, 377–381. [Google Scholar] [CrossRef]
- Ihrmark, K.; Johannesson, H.; Stenström, E.; Stenlid, J. Transmission of double-stranded RNA in Heterobasidion annosum. Fungal Genet. Biol. 2002, 36, 147–154. [Google Scholar] [CrossRef]
- Muñoz-Adalia, E.J.; Fernández, M.M.; Diez, J.J. The use of mycoviruses in the control of forest diseases. Biocontrol Sci. Technol. 2016, 26, 577–604. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Vainio, E.J.; Botella, L.; Hantula, J.; Diez, J.J. Three mitovirus strains infecting a single isolate of Fusarium circinatum are the first putative members of the family Narnaviridae detected in a fungus of the genus Fusarium. Arch. Virol. 2014, 159, 2153–2155. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Martínez-Álvarez, P.; Bezos, D.; Hantula, J.; Diez, J.J. Fusarium circinatum isolates from northern Spain are commonly infected by three distinct mitoviruses. Arch. Virol. 2015, 160, 2093–2098. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Adalia, E.J.; Diez, J.J.; Fernández, M.M.; Hantula, J.; Vainio, E.J. Characterization of small RNAs originating from mitoviruses infecting the conifer pathogen Fusarium circinatum. Arch. Virol. 2018, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Adalia, E.J.; Flores-Pacheco, J.A.; Martínez-Álvarez, P.; Martín-García, J.; Fernández, M.; Diez, J.J. Effect of mycoviruses on the virulence of Fusarium circinatum and laccase activity. Physiol. Mol. Plant Pathol. 2016, 94, 8–15. [Google Scholar] [CrossRef]
- Flores-Pacheco, J.A.; Muñoz-Adalia, E.J.; Martínez-Álvarez, P.; Pando, V.; Diez-Casero, J.J.; Martín-García, J. Short communication: Effect of mycoviruses on growth, spore germination and pathogenicity of the fungus Fusarium circinatum. For. Syst. 2017, 26. [Google Scholar] [CrossRef]
- Vainio, E.J.; Korhonen, K.; Hantula, J. Genetic variation in Phlebiopsis gigantea as detected with random amplified microsatellite (RAMS) markers. Mycol. Res. 1998, 102, 187–192. [Google Scholar] [CrossRef]
- Vainio, E.J.; Hakanpää, J.; Dai, Y.-C.; Hansen, E.; Korhonen, K.; Hantula, J. Species of Heterobasidion host a diverse pool of partitiviruses with global distribution and interspecies transmission. Fungal Biol. 2011, 115, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Jurvansuu, J.; Kashif, M.; Vaario, L.; Vainio, E.; Hantula, J. Partitiviruses of a fungal forest pathogen have species-specific quantities of genome segments and transcripts. Virology 2014, 462, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, G. Chapter 3: Logit Models for Binary Data. In Lecture Notes on Generalized Linear Models. 2007. Available online: http://data.princeton.edu/wws509/notes/ (accessed on 15 January 2018).
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://www.R-project.org/ (accessed on 8 January 2018).
- Harrell, F.E., Jr. Hmisc: Harrell Miscellaneous. R Package Version 4.1-1. 2018. Available online: http://biostat.mc.vanderbilt.edu/Hmisc (accessed on 10 January 2018).
- Jiang, Y.; Zhang, T.; Luo, C.; Jiang, D.; Li, G.; Li, Q.; Hsiang, T.; Huang, J. Prevalence and diversity of mycoviruses infecting the plant pathogen Ustilaginoidea virens. Virus Res. 2015, 195, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Romo, M.; Leuchtmann, A.; García, B.; Zabalgogeazcoa, I. A totivirus infecting the mutualistic fungal endophyte Epichloë festucae. Virus Res. 2007, 124, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Ding, P.; Liu, F.; Xu, C.; Wang, K. Transmission of Cryphonectria hypovirus to protect chestnut trees from chestnut blight disease. Biol. Control 2007, 40, 9–14. [Google Scholar] [CrossRef]
- Zamora, P.; Martín, A.B.; Dueñas, M.; Martin, R.S.; Diez, J.J. Cryphonectria parasitica isolates of the same vegetative compatibility type display different rates of transfer of CHV1 hypovirus. Eur. J. Plant Pathol. 2015, 143, 767–777. [Google Scholar] [CrossRef]
- Zamora, P.; González Casas, A.; Dueñas, M.; San Martin, R.; Diez, J.J. Factors influencing growth, sporulation and virus transfer in Cryphonectria parasitica isolates from Castilla and León (Spain). Eur. J. Plant Pathol. 2017, 148, 65–73. [Google Scholar] [CrossRef]
- Anagnostakis, S.L.; Chen, B.; Geletka, L.M.; Nuss, D.L. Hypovirus Transmission to Ascospore Progeny by Field-Released Transgenic Hypovirulent Strains of Cryphonectria parasitica. Phytopathology 1998, 88, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Nakamura, H.; Arakawa, M.; Matsumoto, N. Diversity and vertical transmission of double-stranded RNA elements in root rot pathogens of trees, Helicobasidium mompa and Rosellinia necatrix. Mycol. Res. 2004, 108, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Ihrmark, K.; Stenström, E.; Stenlid, J. Double-stranded RNA transmission through basidiospores of Heterobasidion annosum. Mycol. Res. 2004, 108, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Won, H.K.; Park, S.J.; Kim, D.K.; Shin, M.J.; Kim, N.; Lee, S.H.; Kwon, Y.C.; Ko, H.K.; Ro, H.S.; Lee, H.S. Isolation and characterization of a mycovirus in Lentinula edodes. J. Microbiol. 2013, 51, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Day, T.; Proulx, S.R. A General Theory for the Evolutionary Dynamics of Virulence. Am. Nat. 2004, 163, E40–E63. [Google Scholar] [CrossRef] [PubMed]
- Brusini, J.; Wayne, M.L.; Franc, A.; Robin, C. The impact of parasitism on resource allocation in a fungal host: The case of Cryphonectria parasitica and its mycovirus, Cryphonectria Hypovirus 1. Ecol. Evol. 2017, 7, 5967–5976. [Google Scholar] [CrossRef] [PubMed]
- Göker, M.; Scheuner, C.; Klenk, H.-P.; Stielow, J.B.; Menzel, W. Codivergence of Mycoviruses with Their Hosts. PLoS ONE 2011, 6, e22252. [Google Scholar] [CrossRef] [PubMed]
- Hyder, R.; Pennanen, T.; Hamberg, L.; Vainio, E.J.; Piri, T.; Hantula, J. Two viruses of Heterobasidion confer beneficial, cryptic or detrimental effects to their hosts in different situations. Fungal Ecol. 2013, 6, 387–396. [Google Scholar] [CrossRef]
- Cho, W.K.; Lee, K.-M.; Yu, J.; Son, M.; Kim, K.-H. Insight into Mycoviruses Infecting Fusarium Species. In Mycoviruses; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Wu, M.; Zhang, L.; Li, G.; Jiang, D.; Ghabrial, S. A Genome characterization of a debilitation-associated mitovirus infecting the phytopathogenic fungus Botrytis cinerea. Virology 2010, 406, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wu, S.; Liu, L.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2015, 197, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Varese, G.C.; Falk, B.W.; Turina, M. Mycoviruses of an endophytic fungus can replicate in plant cells: Evolutionary implications. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Márquez, L.M.; Redman, R.S.; Rodriguez, R.J.; Roossinck, M.J. A Virus in a Fungus in a Plant: Three-Way Symbiosis Required for Thermal Tolerance. Science 2007, 315, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Herrero, N.; Pérez-Sánchez, R.; Oleaga, A.; Zabalgogeazcoa, I. Tick pathogenicity, thermal tolerance and virus infection in Tolypocladium cylindrosporum. Ann. Appl. Biol. 2011, 159, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Iturritxa, E.; Ganley, R.J.; Wright, J.; Heppe, E.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, M.J. A genetically homogenous population of Fusarium circinatum causes pitch canker of Pinus radiata in the Basque Country, Spain. Fungal Biol. 2011, 115, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Berbegal, M.; Pérez-Sierra, A.; Armengol, J.; Grünwald, N.J. Evidence for Multiple Introductions and Clonality in Spanish Populations of Fusarium circinatum. Phytopathology 2013, 103, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Mycoviruses, RNA Silencing, and Viral RNA Recombination. Adv. Virus Res. 2011, 80, 25–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brusini, J.; Robin, C. Mycovirus transmission revisited by in situ pairings of vegetatively incompatible isolates of Cryphonectria parasitica. J. Virol. Methods 2013, 187, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortesi, P.; McCulloch, C.E.; Song, H.; Lin, H.; Milgroom, M.G. Genetic control of horizontal virus transmission in the chestnut blight fungus, Cryphonectria parasitica. Genetics 2001, 159, 107–118. [Google Scholar] [PubMed]
- Dawe, A.L.; Nuss, D.L. Hypovirus Molecular Biology. From Koch’s Postulates to Host Self-Recognition Genes that Restrict Virus Transmission. Adv. Virus Res. 2013, 86, 109–147. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.H.; Dawe, A.L.; Churbanov, A.; Smith, M.L.; Milgroom, M.G.; Nuss, D.L. Molecular characterization of vegetative incompatibility genes that restrict hypovirus transmission in the chestnut blight fungus Cryphonectria parasitica. Genetics 2012, 190, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.X.; Spiering, M.J.; Dawe, A.L.; Nuss, D.L. Vegetative incompatibility loci with dedicated roles in allorecognition restrict mycovirus transmission in chestnut blight fungus. Genetics 2014, 197, 701–714. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sierra, A.; Landeras, E.; León, M.; Berbegal, M.; García-Jiménez, J.; Armengol, J. Characterization of Fusarium circinatum from Pinus spp. in northern Spain. Mycol. Res. 2007, 111, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Melzer, M.S.; Ikeda, S.S.; Boland, G.J. Interspecific transmission of double-stranded RNA and hypovirulence from Sclerotinia sclerotiorum to S. minor. Phytopathology 2002, 92, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Charlton, N.D.; Cubeta, M.A. Transmission of the M2 double-stranded RNA in Rhizoctonia solani anastomosis group 3 (AG-3). Mycologia 2007, 99, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Yaegashi, H.; Nakamura, H.; Sawahata, T.; Sasaki, A.; Iwanami, Y.; Ito, T.; Kanematsu, S. Appearance of mycovirus-like double-stranded RNAs in the white root rot fungus, Rosellinia necatrix, in an apple orchard. FEMS Microbiol. Ecol. 2013, 83, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Hantula, J. Taxonomy, biogeography and importance of Heterobasidion viruses. Virus Res. 2016, 219, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Liu, Y.-C.; Hillman, B.I.; Milgroom, M.G. Recombination and migration of Cryphonectria hypovirus 1 as inferred from gene genealogies and the coalescent. Genetics 2004, 166, 1611–1629. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-M.; Yu, J.; Son, M.; Lee, Y.-W.; Kim, K.-H. Transmission of Fusarium boothii mycovirus via protoplast fusion causes hypovirulence in other phytopathogenic fungi. PLoS ONE 2011, 6, e21629. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Inoue, K.; Kida, C.; Uwamori, T.; Sasaki, A.; Kanematsu, S.; Park, P. Potentiation of mycovirus transmission by zinc compounds via attenuation of heterogenic incompatibility in Rosellinia necatrix. Appl. Environ. Microbiol. 2013, 79, 3684–3691. [Google Scholar] [CrossRef] [PubMed]
- Bezos, D. (University of Valladolid, Palencia, Spain); Muñoz-Adalia, E.J. (University of Valladolid, Palencia, Spain) Personal communication, 2017.
- Bouneb, M.; Turchetti, T.; Nannelli, R.; Roversi, P.F.; Paoli, F.; Danti, R.; Simoni, S. Occurrence and transmission of mycovirus Cryphonectria hypovirus 1 from dejecta of Thyreophagus corticalis (Acari, Acaridae). Fungal Biol. 2016, 120, 351–357. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample | Host | Region | Locality | Mating Type | FcMV1 | FcMV2-2 |
|---|---|---|---|---|---|---|
| FcCa6 | Pinus radiata | Cantabria | Comillas | MAT-2 | + | − |
| 072 | P. radiata | Cantabria | Comillas | MAT-2 | + | − |
| Fc104 | P. pinaster | Asturias | Unknown | MAT-1 | + | − |
| 020 | P. radiata | Cantabria | Cabezón de la Sal | MAT-2 | − | + |
| FcCa4 | P. radiata | Cantabria | Cabezón de la Sal | MAT-2 | − | + |
| 035 | P. radiata | Cantabria | Cabezón de la Sal | MAT-2 | − | + |
| 042 | P. radiata | Cantabria | Cabezón de la Sal | MAT-2 | − | + |
| FcCa1 | P. radiata | Cantabria | Rionansa | MAT-2 | + | + |
| FcCa70 | P. radiata | Cantabria | Comillas | MAT-2 | + | + |
| Fc221 | P. radiata | Cantabria | Unknown | MAT-2 | + | + |
| Transmission Rate | Area of Fungal Colony 1 | Spore Germination 1 | Relative Length of Necrosis 1 | Area under the Disease Progress Curve 2 | Mycelial Growth 2 | |
|---|---|---|---|---|---|---|
| Transmission rate | 1 | |||||
| Area of fungal colony | r = 0.64 p = 0.13 | 1 | ||||
| Spore germination | r = 0.07 p = 0.89 | r = 0.16 p = 0.73 | 1 | |||
| Relative necrosis length | r = 0.24 p = 0.60 | r = −0.04 p = 0.94 | r = −0.36 p = 0.42 | 1 | ||
| Area under the disease progress curve | r = −0.54 p = 0.21 | r = −0.5 p = 0.25 | r = 0.31 p = 0.50 | r= −0.82 p= 0.02 | 1 | |
| Mycelial growth | r = −0.21 p = 0.66 | r = −0.14 p = 0.76 | r = −0.75 p = 0.05 | r = 0.04 p = 0.94 | r = −0.14 p = 0.76 | 1 |
| Model | Notation | Logit (𝜋ij) | Res Deviance | Res df | AIC |
|---|---|---|---|---|---|
| Null | 𝜙 | 𝜂 | 138 | 129 | |
| Isolate (i) | i | 𝜂 + 𝛼i | 135.92 | 128 | 139.92 |
| Virus (v) | v | 𝜂 + 𝛽j | 123.06 | 120 | 143.06 |
| Additive | i + v | 𝜂 + 𝛼i + 𝛽j | 123.06 | 119 | 145.06 |
| Saturated | i × v | 𝜂 + 𝛼i + 𝛽j + (𝛼𝛽)ij | 117.06 | 117 | 143.06 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romeralo, C.; Bezos, D.; Martínez-Álvarez, P.; Diez, J.J. Vertical Transmission of Fusarium circinatum Mitoviruses FcMV1 and FcMV2-2 via Microconidia. Forests 2018, 9, 356. https://doi.org/10.3390/f9060356
Romeralo C, Bezos D, Martínez-Álvarez P, Diez JJ. Vertical Transmission of Fusarium circinatum Mitoviruses FcMV1 and FcMV2-2 via Microconidia. Forests. 2018; 9(6):356. https://doi.org/10.3390/f9060356
Chicago/Turabian StyleRomeralo, Carmen, Diana Bezos, Pablo Martínez-Álvarez, and Julio Javier Diez. 2018. "Vertical Transmission of Fusarium circinatum Mitoviruses FcMV1 and FcMV2-2 via Microconidia" Forests 9, no. 6: 356. https://doi.org/10.3390/f9060356
APA StyleRomeralo, C., Bezos, D., Martínez-Álvarez, P., & Diez, J. J. (2018). Vertical Transmission of Fusarium circinatum Mitoviruses FcMV1 and FcMV2-2 via Microconidia. Forests, 9(6), 356. https://doi.org/10.3390/f9060356