Different Influences of Vegetation Greening on Regional Water-Energy Balance under Different Climatic Conditions

Abstract
1. Introduction
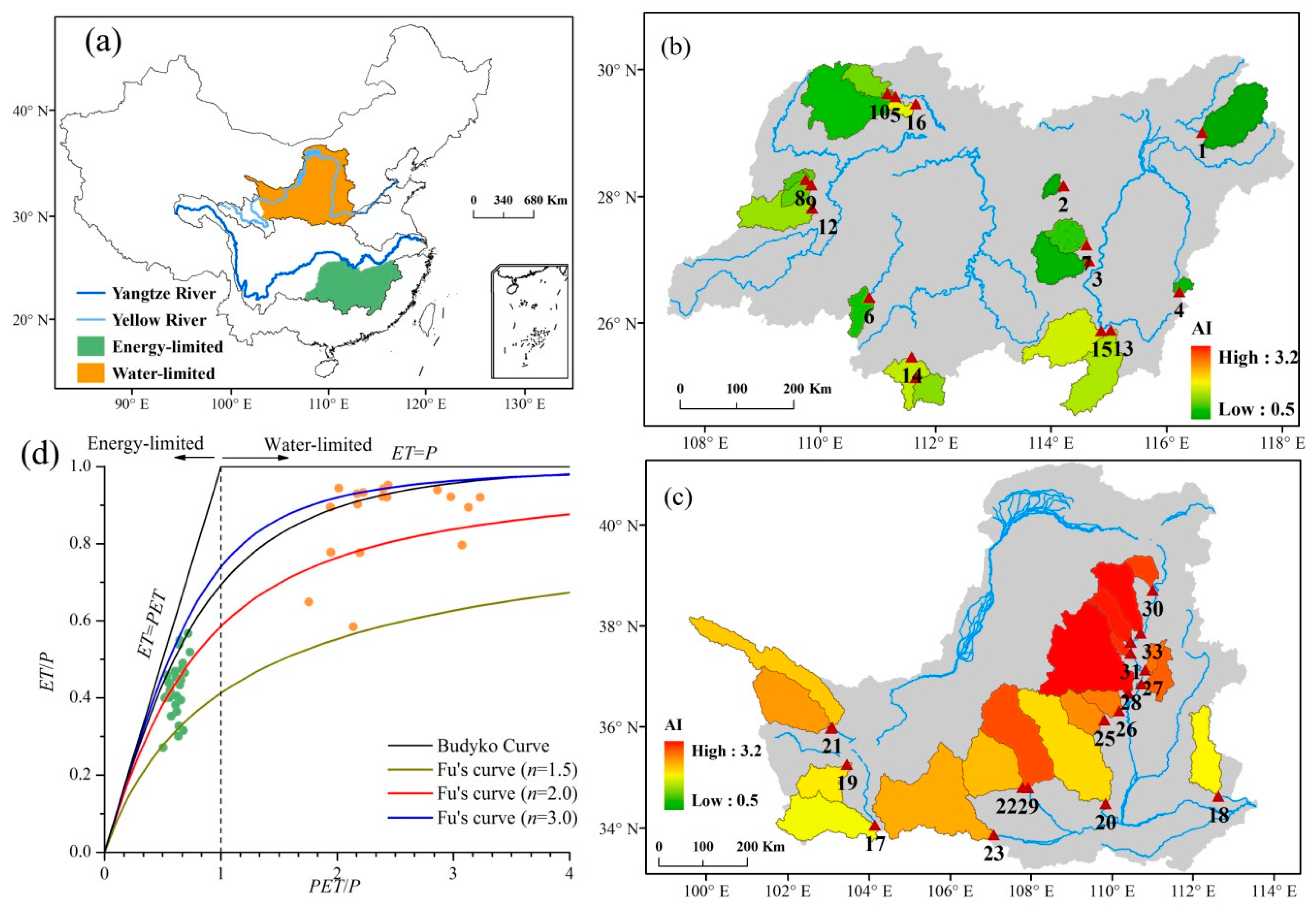
2. Study Area
3. Materials and Methods
3.1. Materials
3.2. Water-Energy Balance Index Considering Vegetation Change
3.3. Sensitivity Analysis
3.4.Contribution Method
4. Results
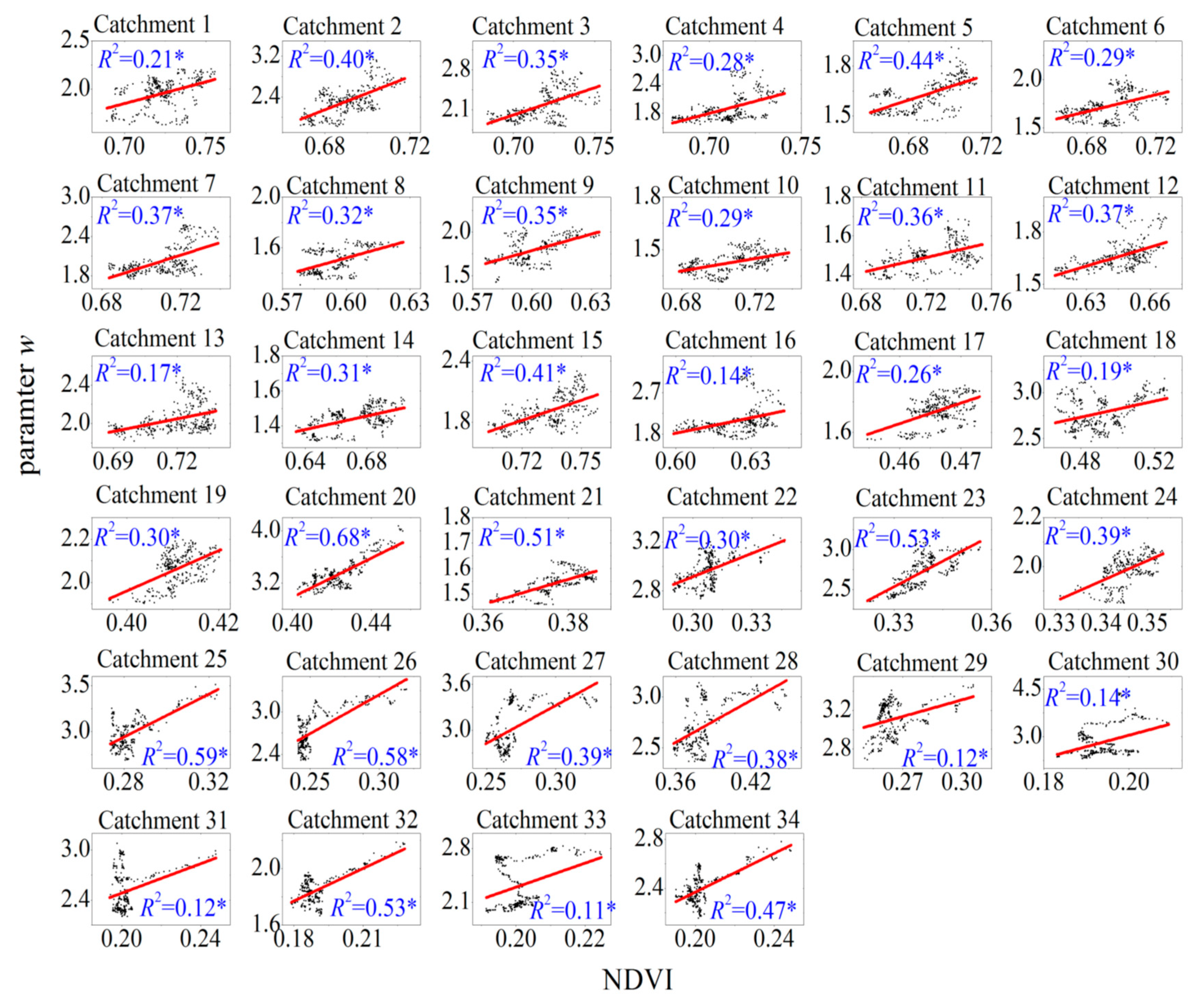
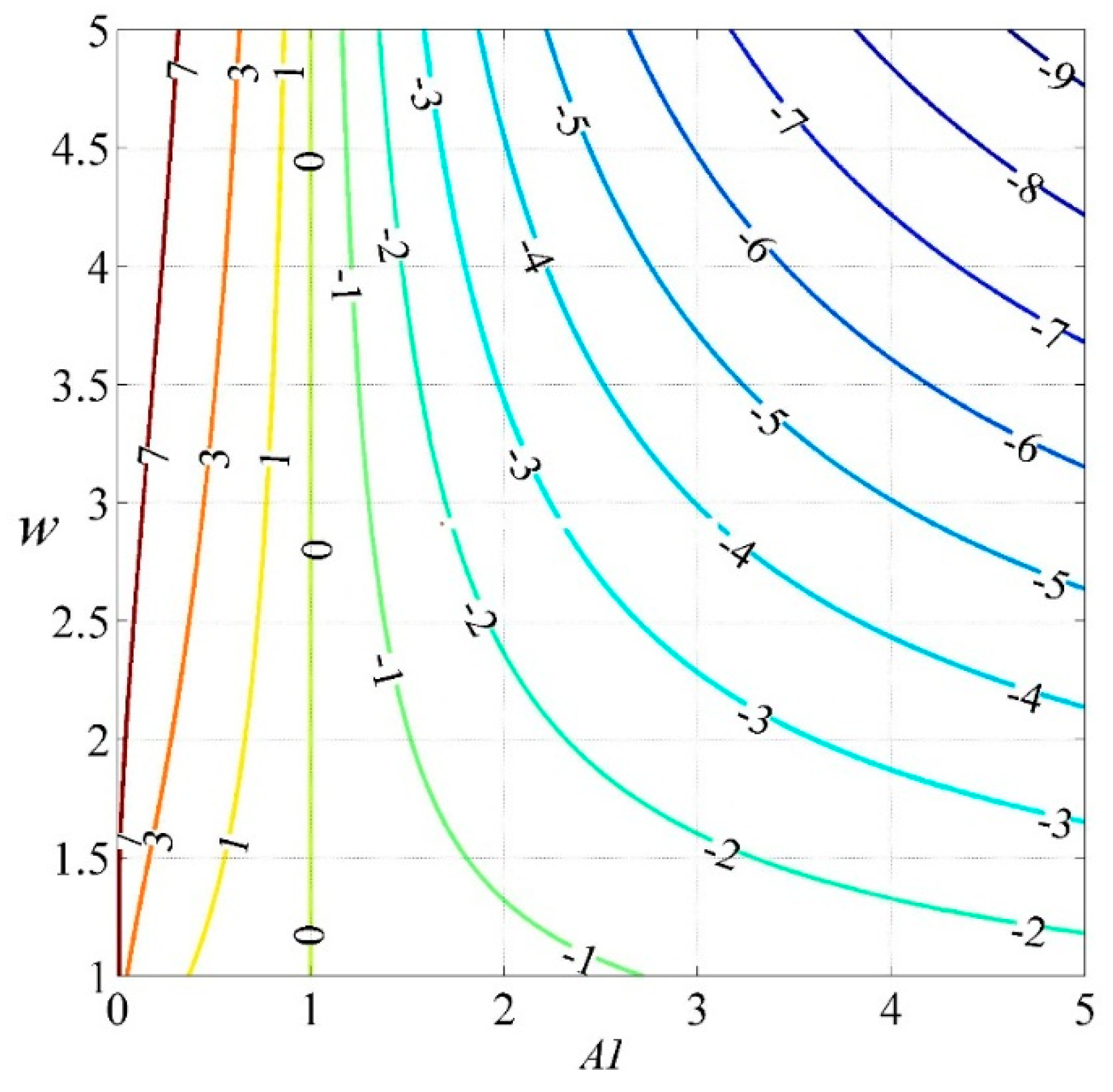
4.1. Relationship between Vegetation and the Land-Surface Parameter w
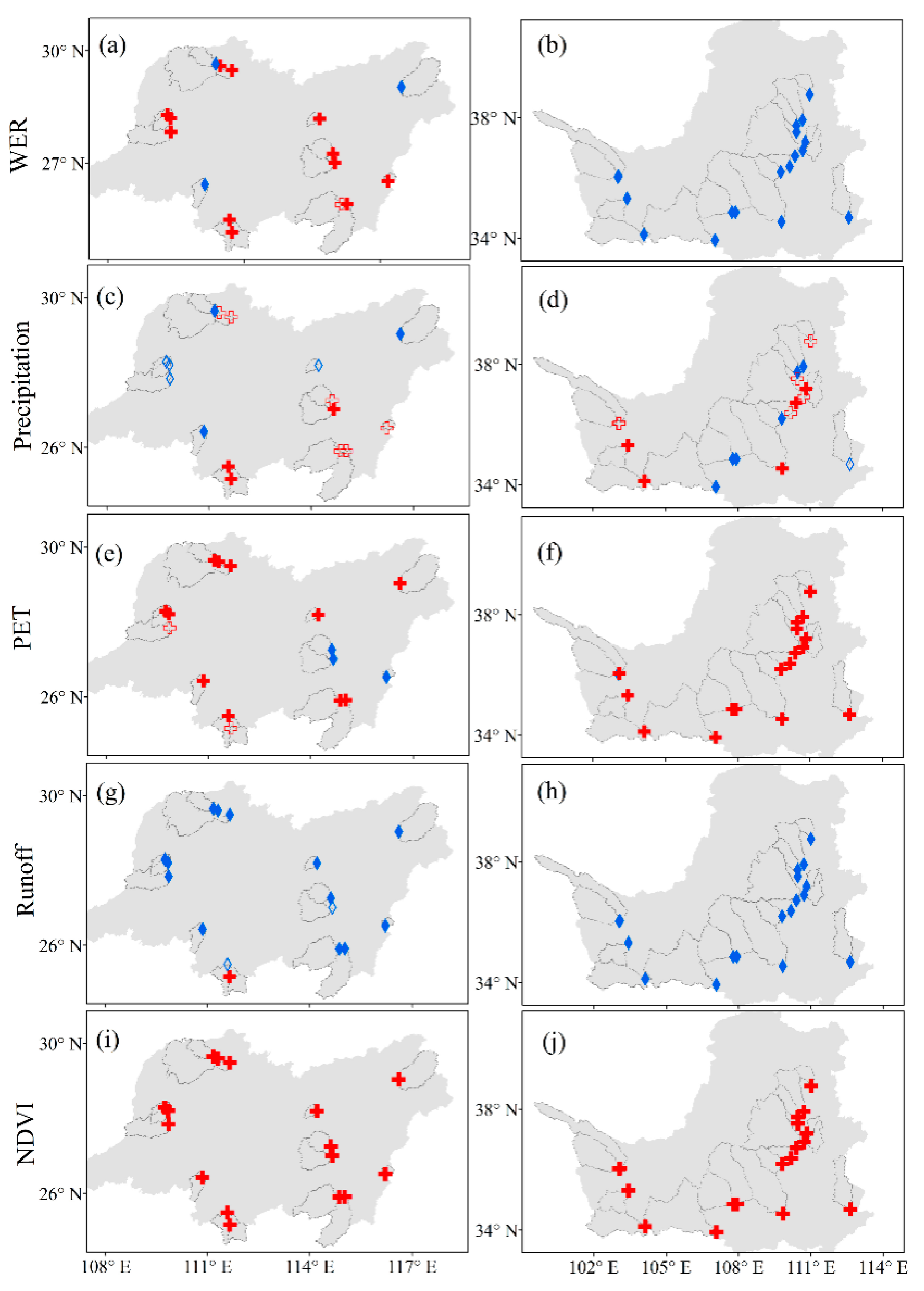
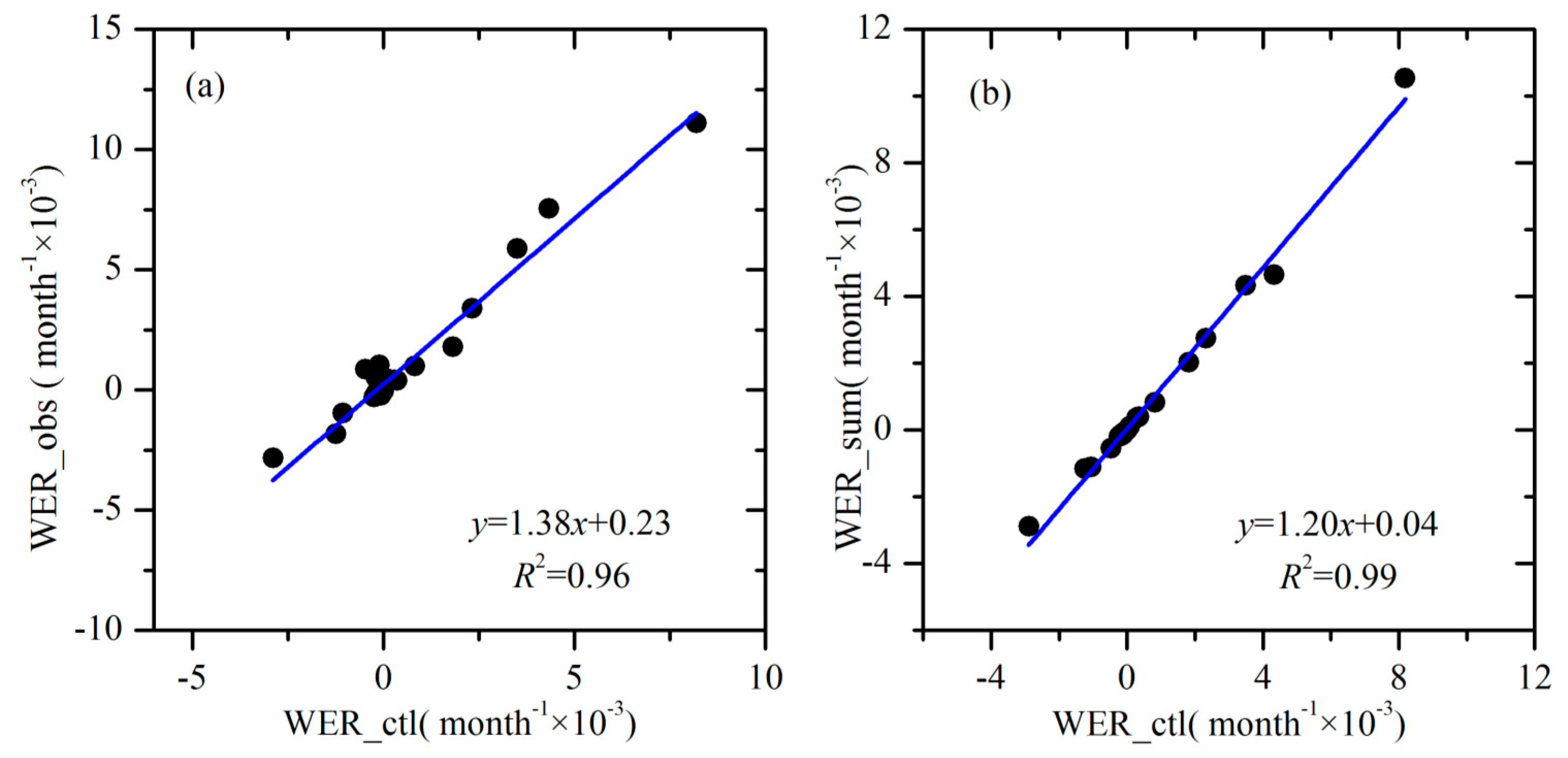
4.2. Changes in WER, Climate Variables and Vegetation
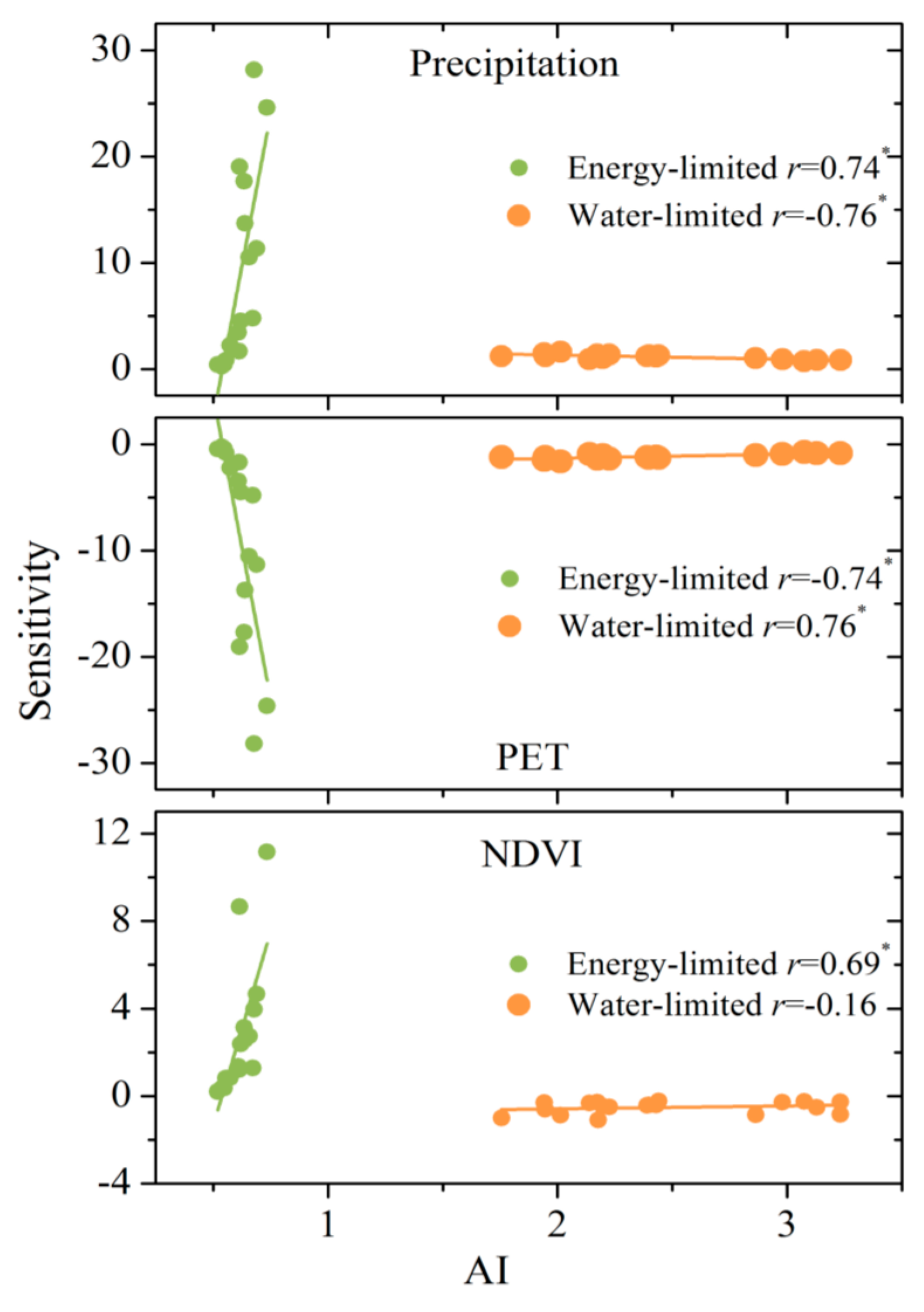
4.3. Sensitivity of WER to Climate Variables and Vegetation
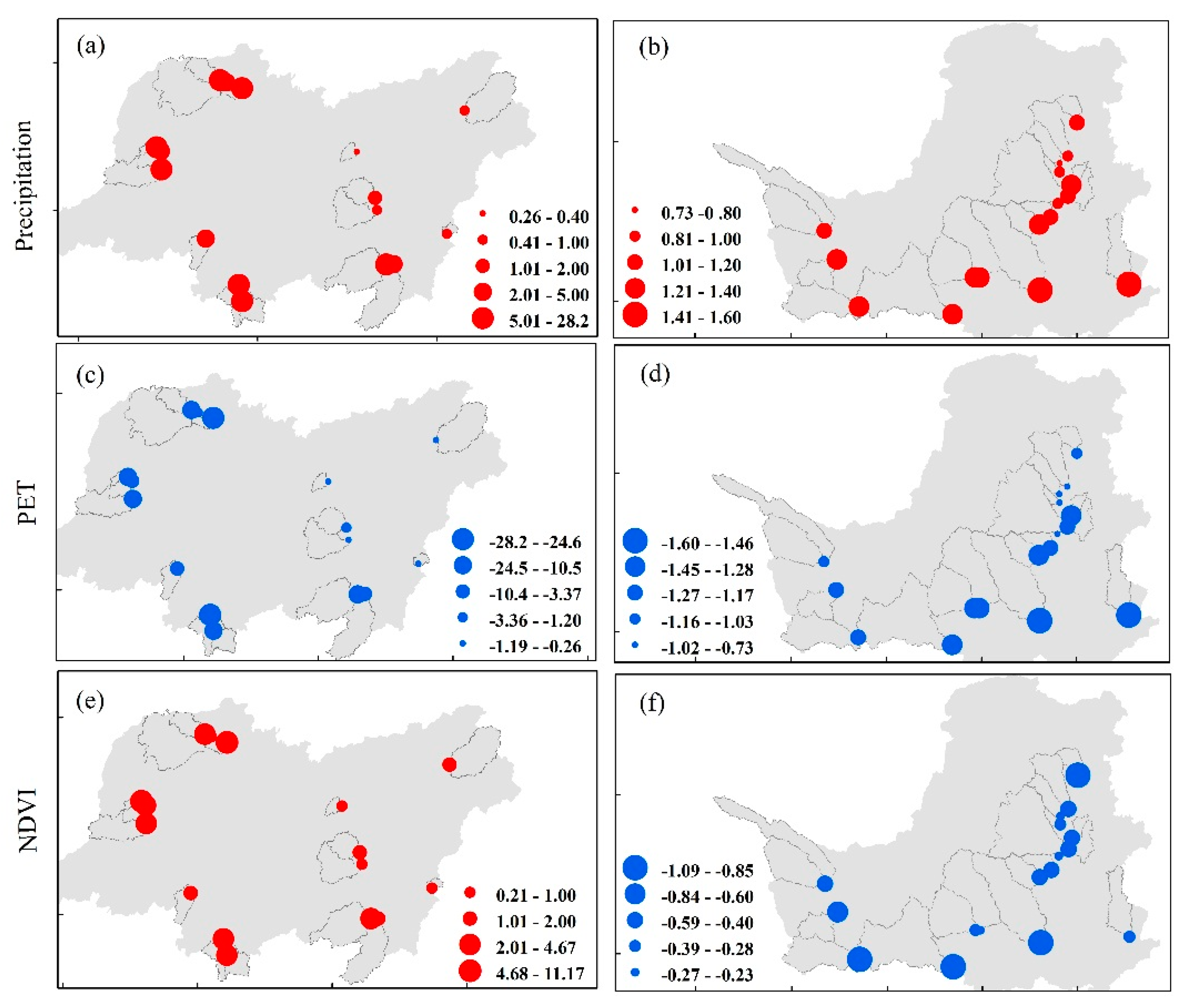
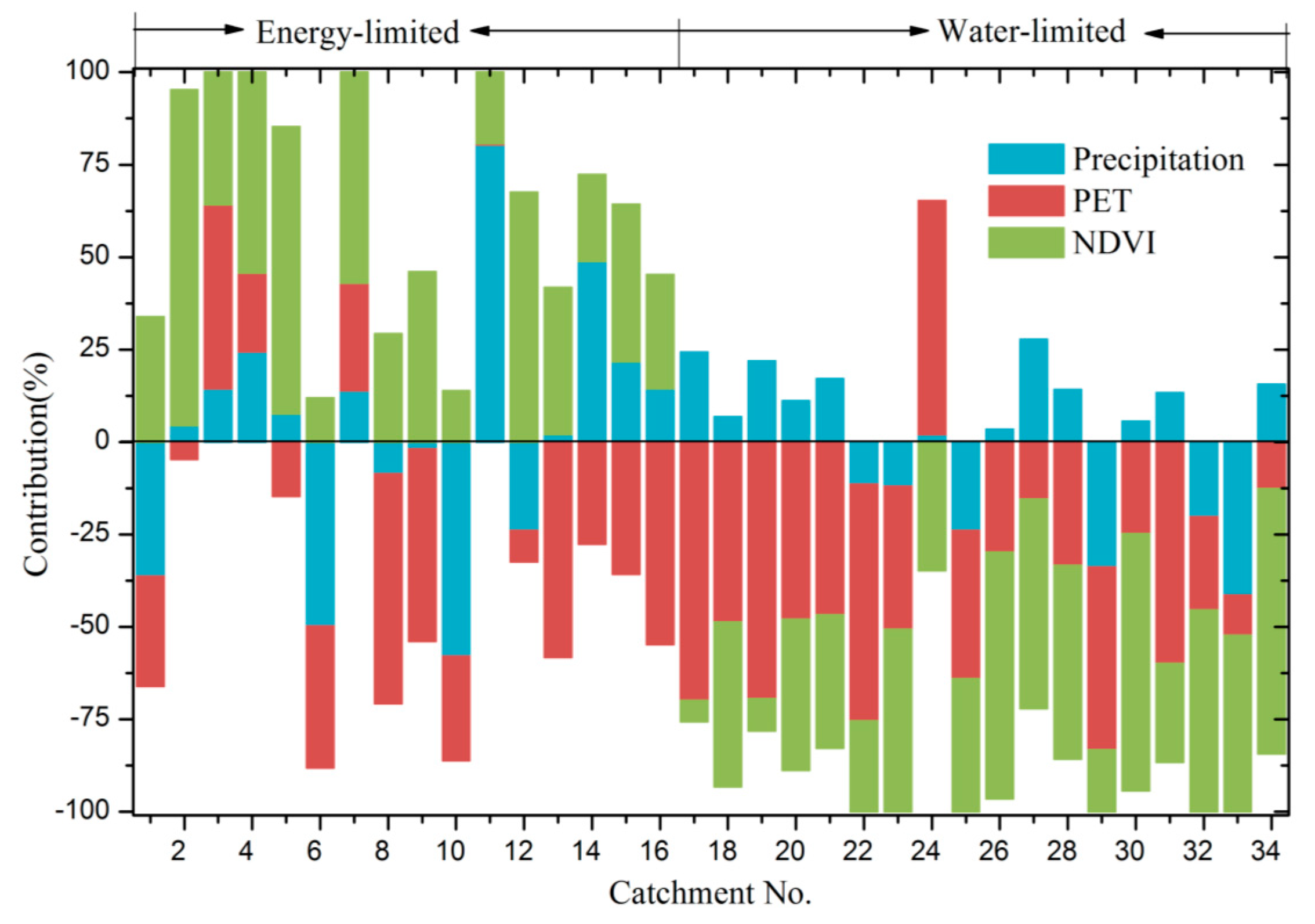
4.4. Quantification of Climate Variation and Vegetation Greening to WER
5. Discussion
5.1. Why Do the Influences of Vegetation Greening on WER Vary?
5.2. Uncertainties
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ivanov, V.Y.; Bras, R.L.; Vivoni, E.R. Vegetation-hydrology dynamics in complex terrain of semiarid areas: 2. Energy-water controls of vegetation spatiotemporal dynamics and topographic niches of favorability. Water Resour. Res. 2008, 44, 380–384. [Google Scholar] [CrossRef]
- Liu, X.; Liu, C.; Brutsaert, W. Investigation of a generalized nonlinear form of the complementary principle for evaporation estimation. J. Geophys. Res. Atmos. 2018, 123, 3933–3942. [Google Scholar] [CrossRef]
- Zhang, D.; Hong, H.; Zhang, Q.; Nie, R. Effects of climatic variation on pan-evaporation in the Poyang Lake Basin, China. Clim. Res. 2014, 61, 29–40. [Google Scholar] [CrossRef]
- Arneth, A. Climate science: Uncertain future for vegetation cover. Nature 2015, 524, 44–45. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, N.; Harper, R.; Li, Q.; Liu, K.; Wei, X.; Ning, D.; Hou, Y.; Liu, S. A global review on hydrological responses to forest change across multiple spatial scales: Importance of scale, climate, forest type and hydrological regime. J. Hydrol. 2016, 546, 44–59. [Google Scholar] [CrossRef]
- Hansen, M.; Potapov, P.; Margono, B.; Stehman, S.; Turubanova, S.; Tyukavina, A. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A. Greening of the Earth and its drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Liu, Y.Y.; de Jeu, R.A.; McCabe, M.F.; Evans, J.P.; van Dijk, A.I. Global long-term passive microwave satellite-based retrievals of vegetation optical depth. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Dijk, A.I.; McCabe, M.F.; Evans, J.P.; Jeu, R.A. Global vegetation biomass change (1988–2008) and attribution to environmental and human drivers. Glob. Ecol. Biogeogr. 2013, 22, 692–705. [Google Scholar] [CrossRef]
- Piao, S.; Wang, X.; Ciais, P.; Zhu, B.; Wang, T.; Liu, J. Changes in satellite-derived vegetation growth trend in temperate and boreal Eurasia from 1982 to 2006. Glob. Chang. Biol. 2011, 17, 3228–3239. [Google Scholar] [CrossRef]
- McDowell, N.G.; Coops, N.C.; Beck, P.S.; Chambers, J.Q.; Gangodagamage, C.; Hicke, J.A.; Huang, C.Y.; Kennedy, R.; Krofcheck, D.J.; Litvak, M.; et al. Global satellite monitoring of climate-induced vegetation disturbances. Trends Plant Sci. 2015, 20, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Nan, H.; Huntingford, C.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Peng, S.; Ahlström, A.; Canadell, J.G.; Cong, N. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Gerten, D.; Schaphoff, S.; Haberlandt, U.; Lucht, W.; Sitch, S. Terrestrial vegetation and water balance—Hydrological evaluation of a dynamic global vegetation model. J. Hydrol. 2004, 286, 249–270. [Google Scholar] [CrossRef]
- Sivapalan, M.; Yaeger, M.A.; Harman, C.J.; Xu, X.; Troch, P.A. Functional model of water balance variability at the catchment scale: 1. Evidence of hydrologic similarity and space-time symmetry. Water Resour. Res. 2011, 47, 2144–2150. [Google Scholar] [CrossRef]
- Budyko, M.I. Climate and Life; Academic Press: London, UK, 1974. [Google Scholar]
- Zhang, L.; Dawes, W.R.; Walker, G.R. Response of mean annual evapotranspiration to vegetation changes at catchment scale. Water Resour. Res. 2001, 37, 701–708. [Google Scholar] [CrossRef]
- Liu, M.; Xu, X.; Xu, C.; Sun, A.Y.; Wang, K.; Scanlon, B.R.; Zhang, L. A new drought index that considers the joint effects of climate and land surface change. Water Resour. Res. 2017, 53. [Google Scholar] [CrossRef]
- Azzali, S.; Menenti, M. Mapping vegetation-soil-climate complexes in southern Africa using temporal Fourier analysis of NOAA-AVHRR NDVI data. Int. J. Remote Sens. 2000, 21, 973–996. [Google Scholar] [CrossRef]
- Donohue, R.J.; Roderick, M.L.; Mcvicar, T.R. On the importance of including vegetation dynamics in Budyko’s hydrological model. Hydrol. Earth Syst. Sci. 2006, 3, 983–995. [Google Scholar] [CrossRef]
- Gentine, P.; D’Odorico, P.; Lintner, B.R.; Sivandran, G.; Salvucci, G. Interdependence of climate, soil, and vegetation as constrained by the Budyko curve. Geophys. Res. Lett. 2012, 39. [Google Scholar] [CrossRef]
- Donohue, R.J.; Roderick, M.L.; Mcvicar, T.R. Can dynamic vegetation information improve the accuracy of Budyko’s hydrological model? J. Hydrol. 2010, 390, 23–34. [Google Scholar] [CrossRef]
- Li, D.; Pan, M.; Cong, Z.; Zhang, L.; Wood, E. Vegetation control on water and energy balance within the Budyko framework. Water Resour. Res. 2013, 49, 969–976. [Google Scholar] [CrossRef]
- Yang, D.; Sun, F.; Liu, Z.; Cong, Z.; Ni, G.; Lei, Z. Analyzing spatial and temporal variability of annual water-energy balance in nonhumid regions of China using the Budyko hypothesis. Water Resour. Res. 2007, 43, 436–451. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, H.; Yang, D.; Jayawardena, A.W. Quantifying the effect of vegetation change on the regional water balance within the Budyko framework. Geophys. Res. Lett. 2016, 43. [Google Scholar] [CrossRef]
- Jackson, R.B.; Jobbágy, E.G.; Avissar, R.; Roy, S.B.; Barrett, D.J.; Cook, C.W.; Farley, K.A.; Maitre, D.C.L.; Mccarl, B.A.; Murray, B.C. Trading Water for Carbon with Biological Carbon Sequestration. Science 2005, 310, 1944–1947. [Google Scholar] [CrossRef] [PubMed]
- Lü, Y.; Zhang, L.; Feng, X.; Zeng, Y.; Fu, B.; Yao, X.; Li, J.; Wu, B. Recent ecological transitions in China: Greening, browning, and influential factors. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Sun, G.; Liu, S.; Jiang, H.; Zhou, G.; Dai, L. The forest-streamflow relationship in China: A 40-year retrospect. J. Am. Water Resour. Assoc. 2008, 44, 1076–1085. [Google Scholar] [CrossRef]
- Feng, X.; Fu, B.; Piao, S.; Wang, S.; Ciais, P.; Zeng, Z.; Lü, Y.; Zeng, Y.; Li, Y.; Jiang, X. Revegetation in China[rsquor]s Loess Plateau is approaching sustainable water resource limits. Nat. Clim. Chang. 2016, 6, 1019–1022. [Google Scholar] [CrossRef]
- Piao, S.; Yin, G.; Tan, J.; Cheng, L.; Huang, M.; Li, Y.; Liu, R.; Mao, J.; Myneni, R.B.; Peng, S. Detection and attribution of vegetation greening trend in China over the last 30 years. Glob. Chang. Biol. 2015, 21, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Huang, Q.; Leng, G.; Zhao, M.; Meng, E. Variations in annual water-energy balance and their correlations with vegetation and soil moisture dynamics: A case study in the Wei River Basin, China. J. Hydrol. 2017, 546, 515–525. [Google Scholar] [CrossRef]
- Ning, T.; Li, Z.; Liu, W. Vegetation dynamics and climate seasonality jointly control the interannual catchment water balance in the Loess Plateau under the Budyko framework. Hydrol. Earth Syst. Sci. 2017, 21, 1–25. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, X.; Zhang, Q.; Liang, K.; Liu, C. Investigation of factors affecting intra-annual variability of evapotranspiration and streamflow under different climate conditions. J. Hydrol. 2016, 543, 759–769. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, M.; Zhuang, D.; Zhang, Z.; Deng, X. Study on spatial pattern of land-use change in China during 1995–2000. Sci. China Ser. D Earth Sci. 2003, 46, 373–384. [Google Scholar]
- McVicar, T.R.; Roderick, M.L.; Donohue, R.J.; Li, L.T.; Van Niel, T.G.; Thomas, A.; Grieser, J.; Jhajharia, D.; Himri, Y.; Mahowald, N.M. Global review and synthesis of trends in observed terrestrial near-surface wind speeds: Implications for evaporation. J. Hydrol. 2012, 416, 182–205. [Google Scholar] [CrossRef]
- Penman, H.L. Natural Evaporation from Open Water, Bare Soil and Grass; The Royal Society: London, UK, 1948; pp. 120–145. [Google Scholar]
- Shuttleworth, W.J. Evaporation. In Handbook of Hydrology; Maidment, D.R., Ed.; McGRaw Hill: New York, NY, USA, 1993. [Google Scholar]
- Beck, H.E.; McVicar, T.R.; van Dijk, A.I.; Schellekens, J.; de Jeu, R.A.; Bruijnzeel, L.A. Global evaluation of four AVHRR–NDVI data sets: Intercomparison and assessment against Landsat imagery. Remote Sens. Environ. 2011, 115, 2547–2563. [Google Scholar] [CrossRef]
- Pinzon, J.E.; Tucker, C.J. A non-stationary 1981–2012 AVHRR NDVI3g time series. Remote Sens. 2014, 6, 6929–6960. [Google Scholar] [CrossRef]
- Carmona, A.; Poveda, G.; Sivapalan, M.; Vallejo-Bernal, S.; Bustamante, E. A scaling approach to Budyko’s framework and the complementary relationship of evapotranspiration in humid environments: Case study of the Amazon River basin. Hydrol. Earth Syst. Sci. 2016, 20, 589–603. [Google Scholar] [CrossRef]
- Arora, V.K. The use of the aridity index to assess climate change effect on annual runoff. J. Hydrol. 2002, 265, 164–177. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, D. Trend analysis of reference evapotranspiration in Northwest China: The roles of changing wind speed and surface air temperature. Hydrol. Process. 2013, 27, 3941–3948. [Google Scholar] [CrossRef]
- Ukkola, A.; Keenan, T.; Kelley, D.I.; Prentice, I. Vegetation plays an important role in mediating future water resources. Environ. Res. Lett. 2016, 11. [Google Scholar] [CrossRef]
- Bai, P.; Liu, X.; Liu, C. Improving hydrological simulations by incorporating GRACE data for model calibration. J. Hydrol. 2018, 557, 291–304. [Google Scholar] [CrossRef]
- Miao, C.; Ashouri, H.; Hsu, K.L.; Sorooshian, S.; Duan, Q. Evaluation of the PERSIANN-CDR Daily Rainfall Estimates in Capturing the Behavior of Extreme Precipitation Events over China. J. Hydrometeorol. 2015, 16, 1387–1396. [Google Scholar] [CrossRef]
- Miao, C.; Kong, D.; Wu, J.; Duan, Q. Functional degradation of the water–sediment regulation scheme in the lower Yellow River: Spatial and temporal analyses. Sci. Total Environ. 2016, 551, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Wang, Q. Characterization and causes of low-quality and low-efficiency forest in Ganzhou of Jiangxi Province. Prot. For. Sci. Technol. 2016. [Google Scholar] [CrossRef]
- Behling, H. Late Quaternary vegetation, climate and fire history of the Araucaria forest and campos region from Serra Campos Gerais, ParanáState (South Brazil). Rev. Palaeobot. Palynol. 1997, 97, 109–121. [Google Scholar] [CrossRef]
- Lehmann, C.E.; Anderson, T.M.; Sankaran, M.; Higgins, S.I.; Archibald, S.; Hoffmann, W.A.; Hanan, N.P.; Williams, R.J.; Fensham, R.J.; Felfili, J. Savanna vegetation-fire-climate relationships differ among continents. Science 2014, 343, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Hundecha, Y.; Bárdossy, A. Modeling of the effect of land use changes on the runoff generation of a river basin through parameter regionalization of a watershed model. J. Hydrol. 2004, 292, 281–295. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catchments Characteristics | Energy-Limited | Water-Limited |
|---|---|---|
| number | 16 | 18 |
| catchment area (km2) | 656–15,307 | 1121–30,661 |
| air temperature (°C) | 14.8–19.7 | 0.9–11.3 |
| annual rainfall (mm) | 1279–1852 | 388–683 |
| aridity index | 0.5–0.8 | 1.7–3.2 |
| Experiment | Description |
|---|---|
| Control test:WER_ctr | Precipitation, PET and NDVI from 1982–2013 |
| Sensitivity test:WER_Prcp | Precipitation maintained at the initial year, the others same as the control test |
| Sensitivity test:WER_PET | PET maintained at the initial year, the others same as the control test |
| Sensitivity test:WER_NDVI | NDVI maintained at the initial year, the others same as the control test |
| Catchment No. | AI | R2 | a | b | Catchment No. | AI | R2 | a | b |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.52 | 0.21 * | 4.5 | −1.3 | 18 | 1.94 | 0.19 * | 4.39 | 0.61 |
| 2 | 0.54 | 0.40 * | 16.71 | −9.22 | 19 | 1.95 | 0.30 * | 9.38 | −1.79 |
| 3 | 0.55 | 0.35 * | 9.44 | −4.59 | 20 | 2.01 | 0.68 * | 14.9 | −2.98 |
| 4 | 0.56 | 0.28 * | 10.09 | −5.26 | 21 | 2.14 | 0.51 * | 4.91 | −0.31 |
| 5 | 0.57 | 0.44 * | 3.6 | −0.85 | 22 | 2.17 | 0.30 * | 6.41 | 0.99 |
| 6 | 0.61 | 0.29 * | 4.39 | −1.32 | 23 | 2.18 | 0.53 * | 21.44 | −4.54 |
| 7 | 0.61 | 0.37 * | 9.47 | −4.7 | 24 | 2.2 | 0.39 * | 8.29 | −0.88 |
| 8 | 0.61 | 0.32 * | 4.66 | −1.28 | 25 | 2.23 | 0.59 * | 11.7 | −0.36 |
| 9 | 0.62 | 0.35 * | 6.99 | −2.42 | 26 | 2.39 | 0.58 * | 11.02 | −0.06 |
| 10 | 0.64 | 0.29 * | 1.79 | 0.16 | 27 | 2.4 | 0.39 * | 9.91 | 0.35 |
| 11 | 0.64 | 0.36 * | 1.96 | 0.08 | 28 | 2.43 | 0.38 * | 6.86 | 0.08 |
| 12 | 0.66 | 0.37 * | 3.63 | −0.68 | 29 | 2.44 | 0.12 * | 5.76 | 1.58 |
| 13 | 0.67 | 0.17 * | 4.36 | −1.09 | 30 | 2.86 | 0.14 * | 36.12 | −4.18 |
| 14 | 0.68 | 0.31 * | 1.97 | 0.11 | 31 | 2.98 | 0.12 * | 8.81 | 0.72 |
| 15 | 0.69 | 0.41 * | 6.54 | −2.89 | 32 | 3.07 | 0.53 * | 7.66 | 0.39 |
| 16 | 0.73 | 0.14 * | 11.28 | −4.95 | 33 | 3.13 | 0.11 * | 15.72 | −0.85 |
| 17 | 1.76 | 0.26 * | 13.49 | −4.55 | 34 | 3.23 | 0.47 * | 7.92 | 0.79 |
| Group | Trend | WER | Precipitation | PET | Runoff | NDVI |
|---|---|---|---|---|---|---|
| Energy-limited | Increase (p < 0.01) | 12 | 3 | 11 | 1 | 16 |
| Increase | 1 | 6 | 2 | 0 | 0 | |
| Decrease | 0 | 4 | 0 | 2 | 0 | |
| Decrease (p < 0.01) | 3 | 3 | 3 | 13 | 0 | |
| Water-limited | Increase (p < 0.01) | 0 | 5 | 17 | 0 | 18 |
| Increase | 0 | 6 | 0 | 0 | 0 | |
| Decrease | 0 | 1 | 0 | 0 | 0 | |
| Decrease (p < 0.01) | 18 | 6 | 1 | 18 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Liu, X.; Bai, P. Different Influences of Vegetation Greening on Regional Water-Energy Balance under Different Climatic Conditions. Forests 2018, 9, 412. https://doi.org/10.3390/f9070412
Zhang D, Liu X, Bai P. Different Influences of Vegetation Greening on Regional Water-Energy Balance under Different Climatic Conditions. Forests. 2018; 9(7):412. https://doi.org/10.3390/f9070412
Chicago/Turabian StyleZhang, Dan, Xiaomang Liu, and Peng Bai. 2018. "Different Influences of Vegetation Greening on Regional Water-Energy Balance under Different Climatic Conditions" Forests 9, no. 7: 412. https://doi.org/10.3390/f9070412
APA StyleZhang, D., Liu, X., & Bai, P. (2018). Different Influences of Vegetation Greening on Regional Water-Energy Balance under Different Climatic Conditions. Forests, 9(7), 412. https://doi.org/10.3390/f9070412