Blackcurrant Leaf Chlorosis Associated Virus: Evidence of the Presence of Circular RNA during Infections


Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Source
2.2. Double-Stranded RNA Extraction, Complementary (c)DNA Library Creation, and Illumina NGS Sequencing
2.3. Total RNA Extractions
2.4. Complementary (c)DNA Production, Cloning, Sanger Sequencing
2.5. Genome Assembly and Sequence Analysis
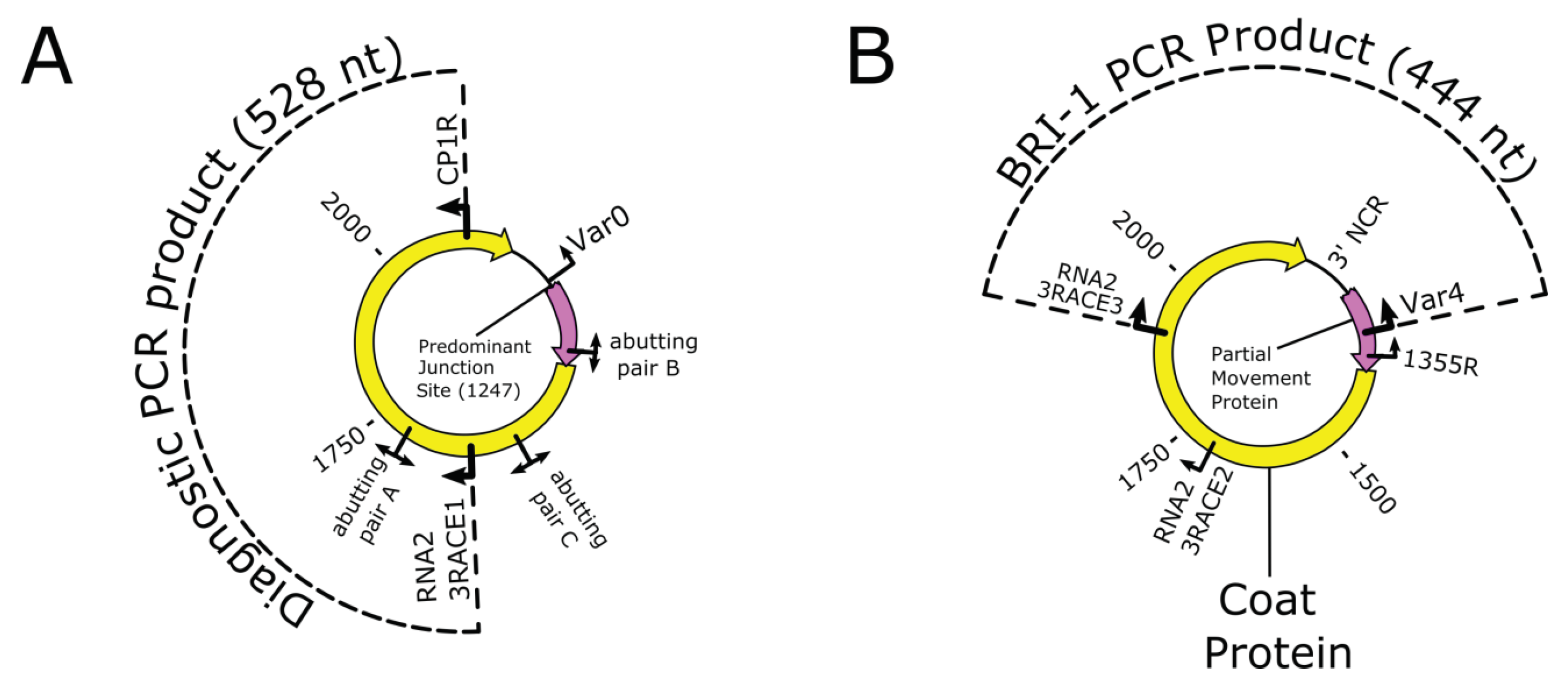
2.6. BCLCaV Diagnostic RT-PCR
2.7. RT-PCR to Verify NGS Generated Bridge Reads
2.8. RT-PCR with Abutting Primers
2.9. RNase R Enzyme Treatments Followed by RT-PCR Analysis
3. Results
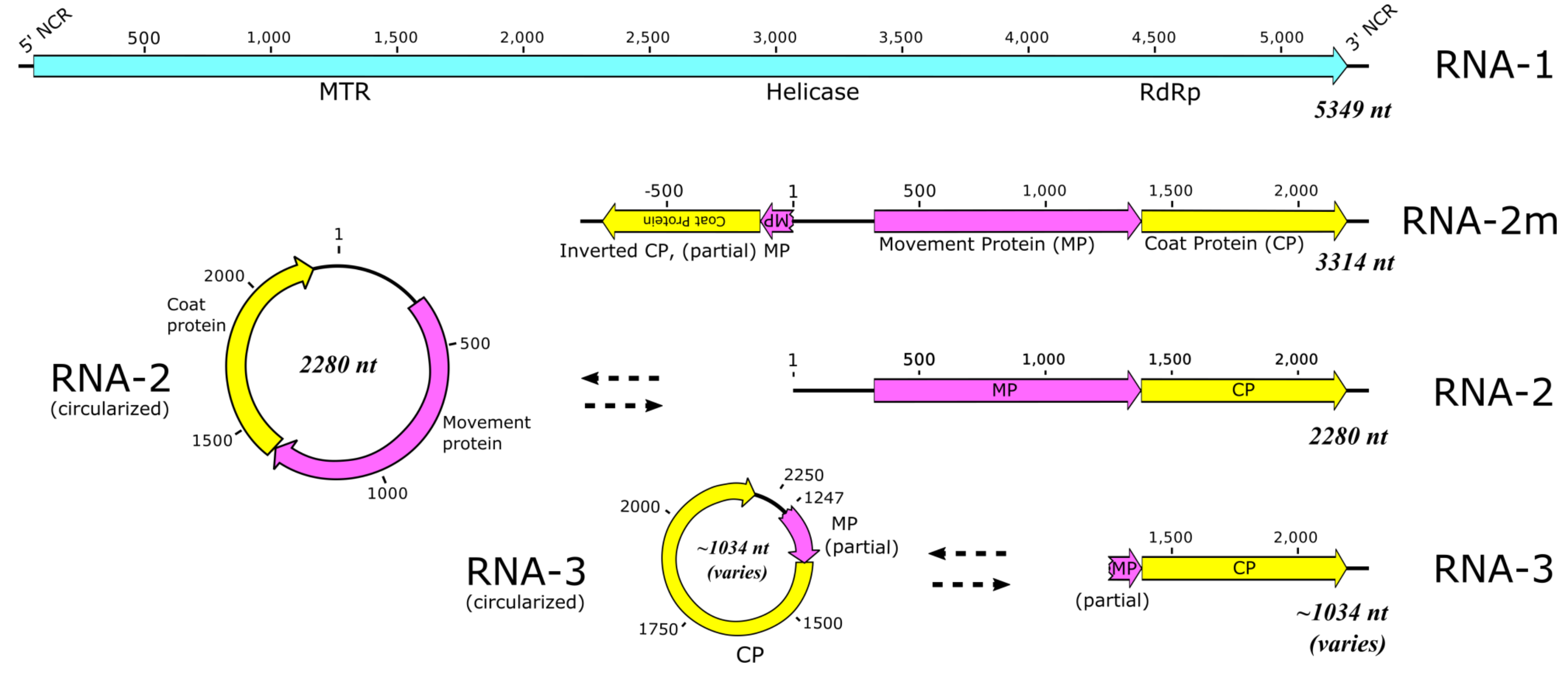
3.1. Sequence Data, Coverage, and Analysis
3.2. RT-PCR Verification of Bridge Reads
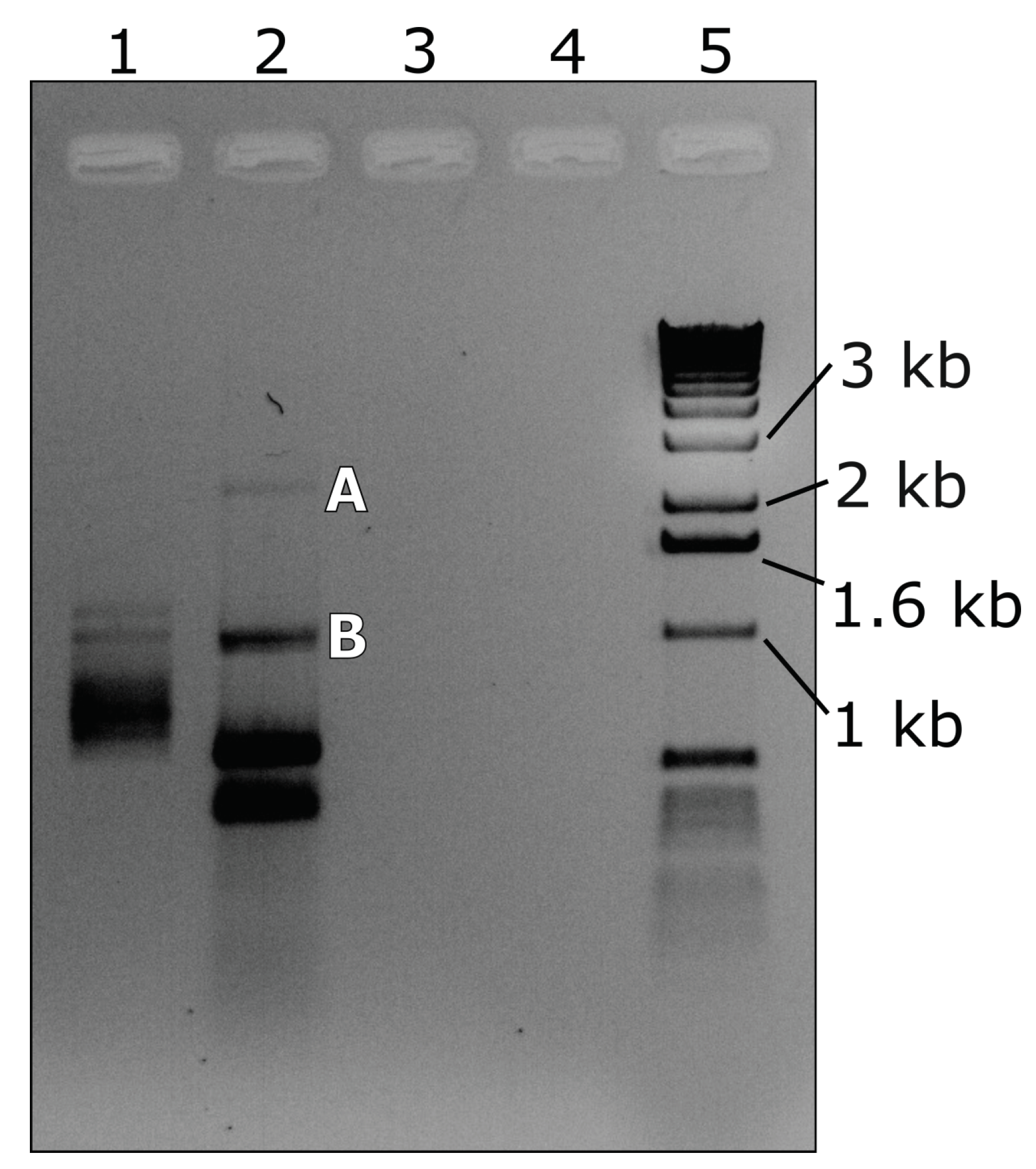
3.3. RT-PCR with Abutting Primers
3.4. RNase R Enzyme Treatments Followed by RT-PCR Analysis
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- James, D.; Phelan, J. Complete genome sequence and analysis of blackcurrant leaf chlorosis associated virus, a new member of the genus Idaeovirus. Arch. Virol. 2017, 161, 1705–1709. [Google Scholar] [CrossRef] [PubMed]
- Thekke-Veetil, T.; Ho, T.; Postman, J.D.; Tzanetakis, I.E. Characterization and detection of a novel idaeovirus infecting blackcurrant. Eur. J. Plant Pathol. 2017, 149, 751–757. [Google Scholar] [CrossRef]
- Navarro, B.; Loconsole, G.; Giampetruzzi, A.; Aboughanem-Sabanadzovic, N.; Ragozzino, A.; Ragozzino, E.; Di Serio, F. Identification and characterization of privet leaf blotch-associated virus, a novel idaeovirus. Mol. Plant Pathol. 2017, 18, 925–936. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, S.A. Genus Idaeovirus. In Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: London, UK, 2011; pp. 1173–1175. [Google Scholar]
- Cadman, C.H. Raspberry viruses and virus diseases in Britain. Hort. Res. 1961, 1, 47. [Google Scholar]
- Mayo, M.A.; Jolly, C.A.; Murant, A.F.; Raschke, J.H. Nucleotide sequence of raspberry bushy dwarf virus RNA-3. J. Gen. Virol. 1991, 72, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Natsuaki, T.; Mayo, M.A.; Jolly, C.A.; Murant, A.F. Nucleotide sequence of raspberry bushy dwarf virus RNA-1: A bicistronic component of a bipartite genome. J. Gen. Virol. 1991, 72, 2183–2189. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.; Natsuaki, T.; Mayo, M.A.; Jolly, C.A.; Murant, A.F. Nucleotide sequence of raspberry bushy dwarf virus RNA-1. J. Gen. Virol. 1992, 73, 3213–3218. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.; Mayo, M.A.; Murant, A.F. Proposed classification of the bipartite-genomed raspberry bushy dwarf idaeovirus, with tripartite genomed viruses in the family Bromoviridae. Arch. Virol. 1993, 131, 483–488. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, S.A.; McGavin, W.J. Genome activation by raspberry bushy dwarf virus coat protein. J. Gen. Virol. 2009, 90, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Powell, M.L. Translational termination-reinitiation in RNA viruses. Biochem. Soc. Trans. 2010, 38, 1558–1564. [Google Scholar] [CrossRef] [PubMed]
- Quito-Avila, D.F.; Ibarra, M.A.; Alvarez, R.; Peralta, E.L.; Martin, R.R. A raspberry bushy dwarf virus isolate from Ecuadorean Rubus glaucus contains an additional RNA that is a rearrangement of RNA-2. Arch. Virol. 2014, 159, 2519–2521. [Google Scholar] [CrossRef] [PubMed]
- Herold, J.; Andino, R. Poliovirus RNA replication requires genome circularization through a protein-protein bridge. Mol. Cell 2001, 7, 581–591. [Google Scholar] [CrossRef]
- Greene, J.; Baird, A.-M.; Brady, L.; Lim, M.; Gray, S.G.; McDermott, R.; Finn, S.P. Circular RNAs: Biogenesis, function and role in human diseases. Front. Mol. Biosci. 2017, 4, 38. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.L.; Bao, Y.; Yee, M.-C.; Barrett, S.P.; Hogan, G.J.; Olsen, M.N.; Dinneny, J.R.; Brown, P.O.; Salzman, J. Circular RNA is expressed across the eukaryotic tree of life. PLoS ONE 2014, 9, e90859. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Minoia, S.; Carbonell, A.; Gisel, A.; Delgado, S.; López-Carrasco, A.; Navarro, B.; Di Serio, F. Viroids, the simplest RNA replicons: How they manipulate their hosts for being propagated and how their hosts react for containing the infection. Virus Res. 2015, 209, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef] [PubMed]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Yang, B.; Chen, B.J.; Bliim, N.; Ueberham, U.; Arendt, T.; Janitz, M. The emerging role of circular RNAs in transcriptome regulation. Genomics 2017, 109, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Lasda, E.; Parker, R. Circular RNAs: Diversity of form and function. RNA 2014, 20, 1829–1842. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Prescott, A.G.; Lunness, P.; Chamberlin, L.C.L.; Markham, P.G. Rapid production of full-length, infectious geminivirus clones by abutting primer PCR (AbP-PCR). J. Virol. Methods 1993, 43, 7–20. [Google Scholar] [CrossRef]
- Zhang, Z.; Qi, S.; Tang, N.; Zhang, X.; Chen, S.; Zhu, P.; Ma, L.; Cheng, J.; Xu, Y.; Lu, M.; et al. Discovery of replicating circular RNAs by RNA-seq and computational algorithms. PLoS Pathog. 2014, 10, e1004553. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Zuo, Y.; Wang, J.; Zhang, M.Q.; Malhotra, A.; Mayeda, A. Characterization of RNase R-digested cellular RNA source that consists of lariat and circular RNAs from pre-mRNA splicing. Nucleic Acids Res. 2006, 34, e63. [Google Scholar] [CrossRef] [PubMed]
- Hecker, K.H.; Roux, K.H. High and low annealing temperatures increase both specificity and yield in touchdown and stepdown PCR. Biotechniques 1996, 20, 478–485. [Google Scholar] [PubMed]
- Korbie, D.J.; Mattick, J.S. Touchdown PCR for increased specificity and sensitivity in PCR amplification. Nat. Protoc. 2008, 3, 1452–1456. [Google Scholar] [CrossRef] [PubMed]
- James, D.; Phelan, J. Complete genome sequence of a strain of Actinidia virus X detected in Ribes nigrum cv. Baldwin showing unusual symptoms. Arch. Virol. 2016, 162, 507–511. [Google Scholar] [CrossRef] [PubMed]
- James, D.; Phelan, J. Detection and analysis of a filamentous virus isolated from blackcurrant (Ribes nigrum cv. Baldwin) showing symptoms of leaf chlorosis and deformity. PSJ Plant Virus Dis. Workshop Rep. 2016, 12, 17–29. [Google Scholar]
- Tzanetakis, I.E.; Martin, R.R. A new method for extraction of double-stranded RNA from plants. J. Virol. Methods 2008, 149, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Kalinowska, E.; Chodorska, M.; Paduch-Cichal, E.; Mroczkowska, K. An improved method for RNA isolation from plants using commercial extraction kits. Acta Biochim. Pol. 2012, 59, 391–393. [Google Scholar] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. ClustalW and ClustalX version 2. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M. Mfold webserver for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.; Bernhart, S.H.; Höner zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Menzel, W.; Jelkmann, W.; Maiss, E. Detection of four apple viruses by multiplex RT-PCR assays with coamplification of plant mRNA as internal control. J. Virol. Methods 2002, 99, 81–92. [Google Scholar] [CrossRef]
- Wilusz, J.E. Repetitive elements regulate circular RNA biogenesis. Mob. Genet. Elem. 2015, 5, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Messmer, A.; Sanderson, D.; Braun, G.; Serra, P.; Flores, R.; James, D. Molecular and phylogenetic identification of unique isolates of hammerhead viroid-like RNA from ‘Pacific Gala’ apple (Malus domestica) in Canada. Can. J. Plant Pathol. 2017, 39, 342–353. [Google Scholar] [CrossRef]
- Hossain, S.T.; Malhotra, A.; Deutscher, M.P. How RNase R degrades structured RNA: Role of the helicase activity and the S1 domain. J. Biol. Chem. 2016, 291, 7877–7887. [Google Scholar] [CrossRef] [PubMed]
- Dodds, J.A.; Morris, T.J.; Jordan, R.L. Plant viral double-stranded RNA. Ann. Rev. Phytopathol. 1984, 22, 151–168. [Google Scholar] [CrossRef]
- Petkovic, S.; Müller, S. RNA circularization strategies in vivo and in vitro. Nucleic Acids Res. 2015, 43, 2454–2465. [Google Scholar] [CrossRef] [PubMed]
- Palukaitis, P. Satellite RNAs and satellite viruses. Mol. Plant-Microbe Interact. 2016, 29, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Adams, I.P.; Glover, R.H.; Monger, W.A.; Mumford, R.; Jackeviciene, E.; Navalinskiene, M.; Samuitiene, M.; Boonham, N. Next-generation sequencing and metagenomics analysis: A universal diagnostic tool in plant virology. Mol. Plant Pathol. 2009, 10, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Lavezzo, E.; Militello, V.; Toppo, S.; Palu, G. Applications of next-generation sequencing technologies to diagnostic virology. Int. J. Mol. Sci. 2011, 12, 7861–7884. [Google Scholar] [CrossRef] [PubMed]
- Mokili, J.L.; Rohwer, F.; Dutilh, B.E. Metagenomics and future perspectives in virus discovery. Curr. Opin. Virol. 2012, 2, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Pettit Kneller, E.L.; Rakotondrafara, A.M.; Miller, W.A. Cap-independent translation of plant viral RNAs. Virus Res. 2006, 119, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of circRNAs. Mol. Cell 2017, 66, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.C. The expanding virosphere. Cell Host Microbe 2016, 20, 279–280. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Pair | Primer Names | 5′–3′ Sequence | Location of Primer Binding Sites | Expected RT-PCR Product Size (nt) | |
|---|---|---|---|---|---|
| NAD5 1 | NAD5-F | (F) | GATGCTTCTTGGGGCTTCTTGTT | Host mRNA; NADH dehydrogenase-5 gene | 181 |
| NAD5-R | (R) | CTCCAGTCACCAACATTGGCATAA | |||
| AVX 2 | RivCP2F | (F) | CAGCTTGTACGAGCGTATG | Actinidia virus X; coat protein region | 460 |
| RivXCP4R | (R) | AGCTAGGTTGGAGATGTAATTG | |||
| Diagnostic RNA-1 3 | RNA1 3RACE1 | (F) | GAGCCAAGCTCACAAACACTC | 4625 to 4645 (RNA1) | 710 |
| Var0 | (R) | GGCAACCATCTGGACAATAG | 5315 to 5334 (RNA1) | ||
| Diagnostic RNA-2 2 | RNA2 3RACE1 | (F) | ACCAGCATTTCGCAGTTCAG | 1599 to 1618 (RNA2) | 528 |
| CP1R | (R) | ATTCTACCCAGCGCCGTAAG | 2107 to 2126 (RNA2) |
| Primer Pair | Primer Names | 5′–3′ Sequence | Location along BCLCaV RNA2 | Expected RT-PCR Product Size (nt) 1 | |
|---|---|---|---|---|---|
| BRI-0 | RNA2 3RACE3 | (F) | TGTTGCGGTGGTTGAAGTTG | 1909 to 1928 | 722 (full RNA-2 target) |
| RNA2 5RACE3 | (R) | CGAACGCATACTCACTGAAC | 331 to 350 | ||
| BRI-1 | RNA2 3RACE3 | (F) | TGTTGCGGTGGTTGAAGTTG | 1909 to 1928 | 444 (RNA-3 target) |
| Var4 | (R) | GGCTGACTTGCACTACCTTTC | 1298 to 1318 | ||
| BRI-4 | RNA2 3RACE2 | (F) | TGCCTGAGGGAGAGGTTGTG | 1701 to 1720 | 689 (RNA-3 target) |
| 1355R | (R) | GGCTTTCTGCGGTATTGGTC | 1336 to 1355 |
| Primer Pair | Primer Names | 5′–3′ Sequence | Location along BCLCaV RNA2 | Expected (RNA-3) RT-PCR Product Size 1 (nt) | |
|---|---|---|---|---|---|
| Abutting pair A | RNA2 3RACE2 | (F) | TGCCTGAGGGAGAGGTTGTG | 1701 to 1720 | 1034 |
| Var2 | (R) | AACCCGAGTGGTAGAGGAG | 1681 to 1699 | ||
| Abutting pair B | 1356F | (F) | CCGAACCCTATTACTAGGATCTG | 1356 to 1378 | 1034 |
| 1355R | (R) | GGCTTTCTGCGGTATTGGTC | 1336 to 1355 | ||
| Abutting pair C | 1519F | (F) | ACCCTTCGCTAGTGTGGATCTC | 1519 to 1540 | 1034 |
| Var3 2 | (R) | TCACCCGTCGCTGGATTAG | 1500 to 1518 |
| RT-PCR Test | Topology of Target | Strandedness of Target | Post Digest PCR Amplification | Amplification of All No-Enzyme Controls | ||
|---|---|---|---|---|---|---|
| After 1 Round | After 2 Rounds | After 3 Rounds | ||||
| NAD5 1 | Linear | ssRNA | not done | N 4 | not done | Y 5 |
| AVX 2 | Linear | ssRNA with dsRNA phase | Y | Y | N | Y |
| BCLCaV 3 RNA1 diagnostic | Linear, not known to be circular | ssRNA with dsRNA phase | Y | N | N | Y |
| BCLCaV RNA2 diagnostic | linear + hypothesized circular | ssRNA with dsRNA phase | Y | Y | Y | Y |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, D.; Phelan, J.; Sanderson, D. Blackcurrant Leaf Chlorosis Associated Virus: Evidence of the Presence of Circular RNA during Infections. Viruses 2018, 10, 260. https://doi.org/10.3390/v10050260
James D, Phelan J, Sanderson D. Blackcurrant Leaf Chlorosis Associated Virus: Evidence of the Presence of Circular RNA during Infections. Viruses. 2018; 10(5):260. https://doi.org/10.3390/v10050260
Chicago/Turabian StyleJames, Delano, James Phelan, and Daniel Sanderson. 2018. "Blackcurrant Leaf Chlorosis Associated Virus: Evidence of the Presence of Circular RNA during Infections" Viruses 10, no. 5: 260. https://doi.org/10.3390/v10050260
APA StyleJames, D., Phelan, J., & Sanderson, D. (2018). Blackcurrant Leaf Chlorosis Associated Virus: Evidence of the Presence of Circular RNA during Infections. Viruses, 10(5), 260. https://doi.org/10.3390/v10050260