Zika Virus Surveillance at the Human–Animal Interface in West-Central Brazil, 2017–2018

 ,
,  , , , ,
, , , ,  , , ,
, , ,  , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Material and Methods
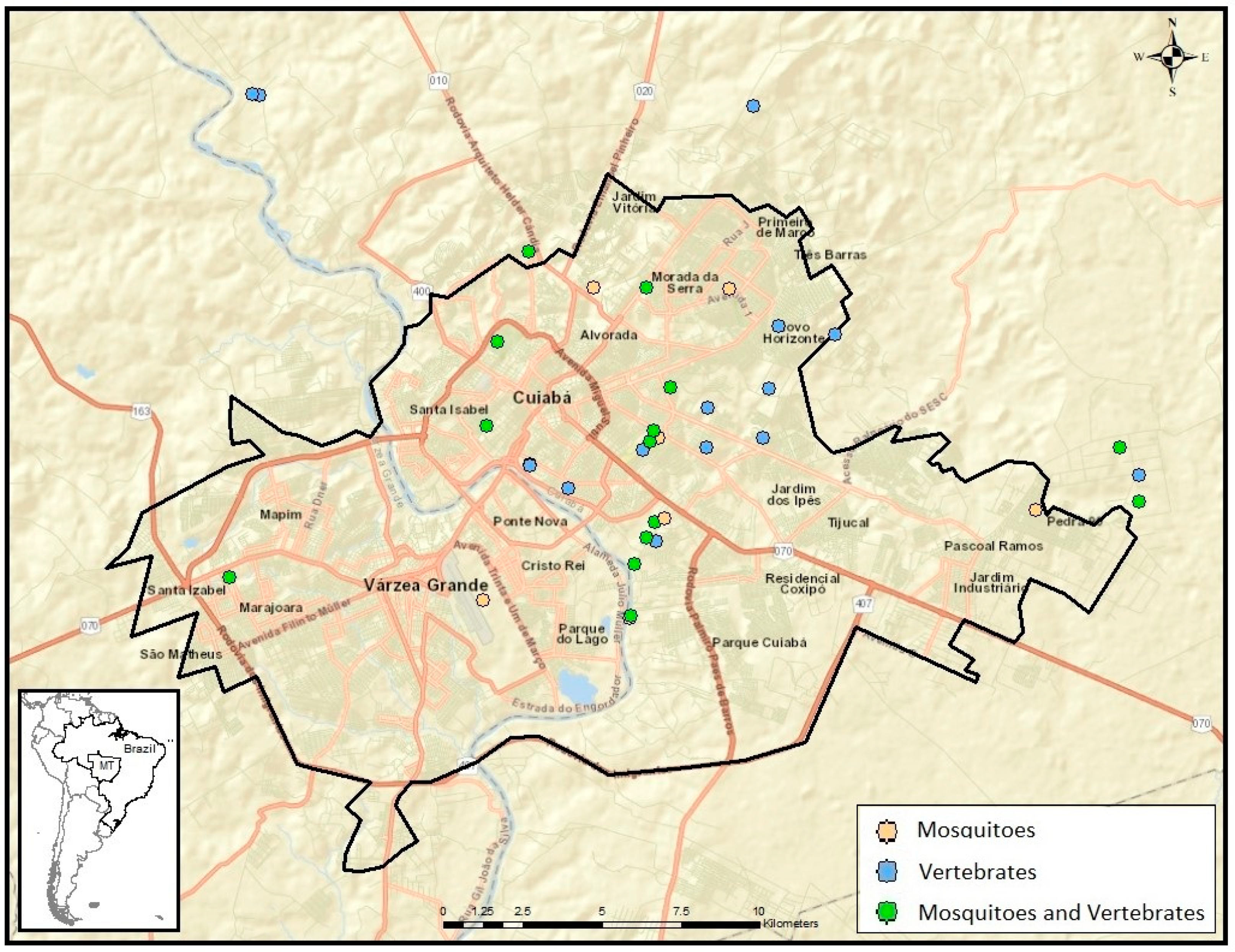
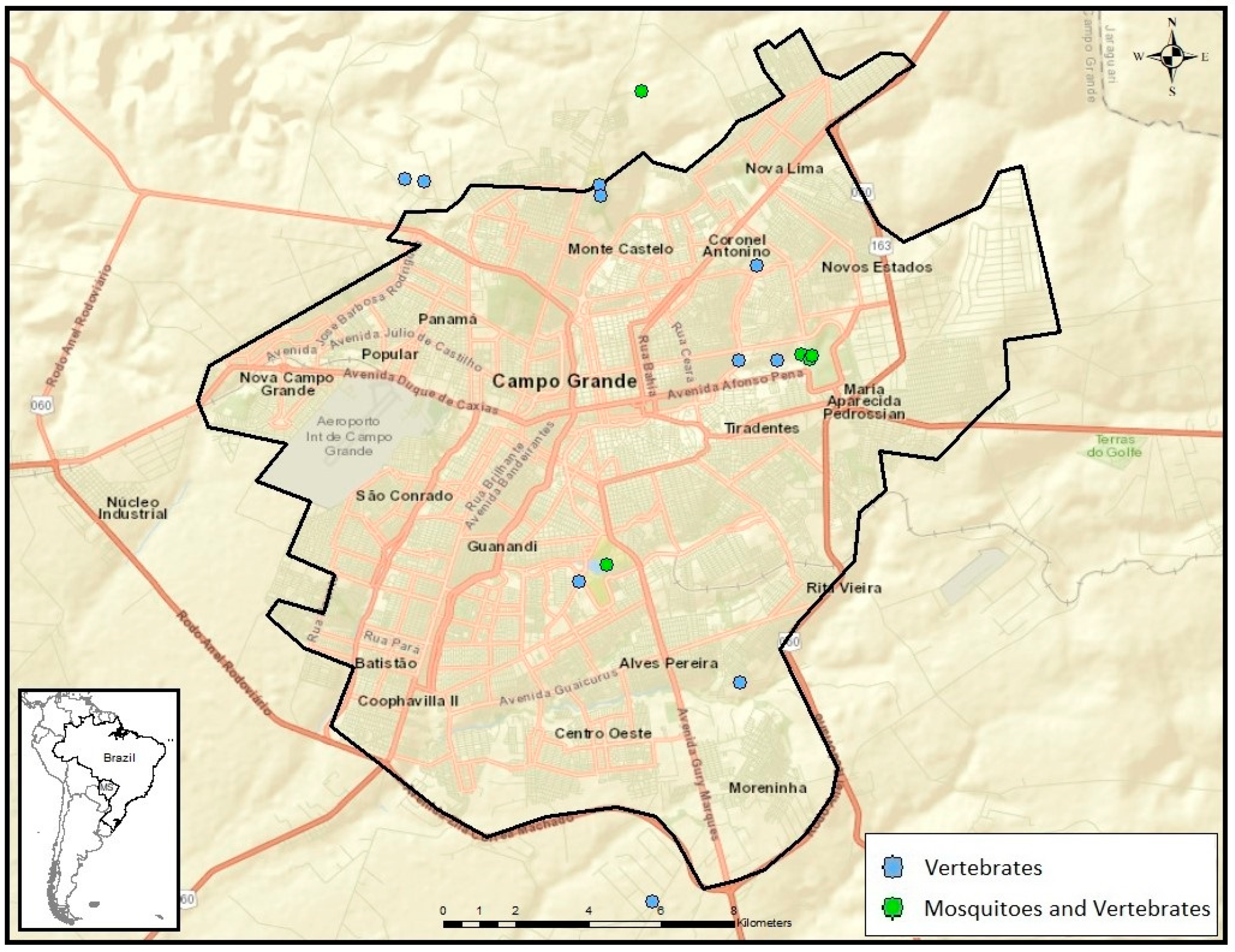
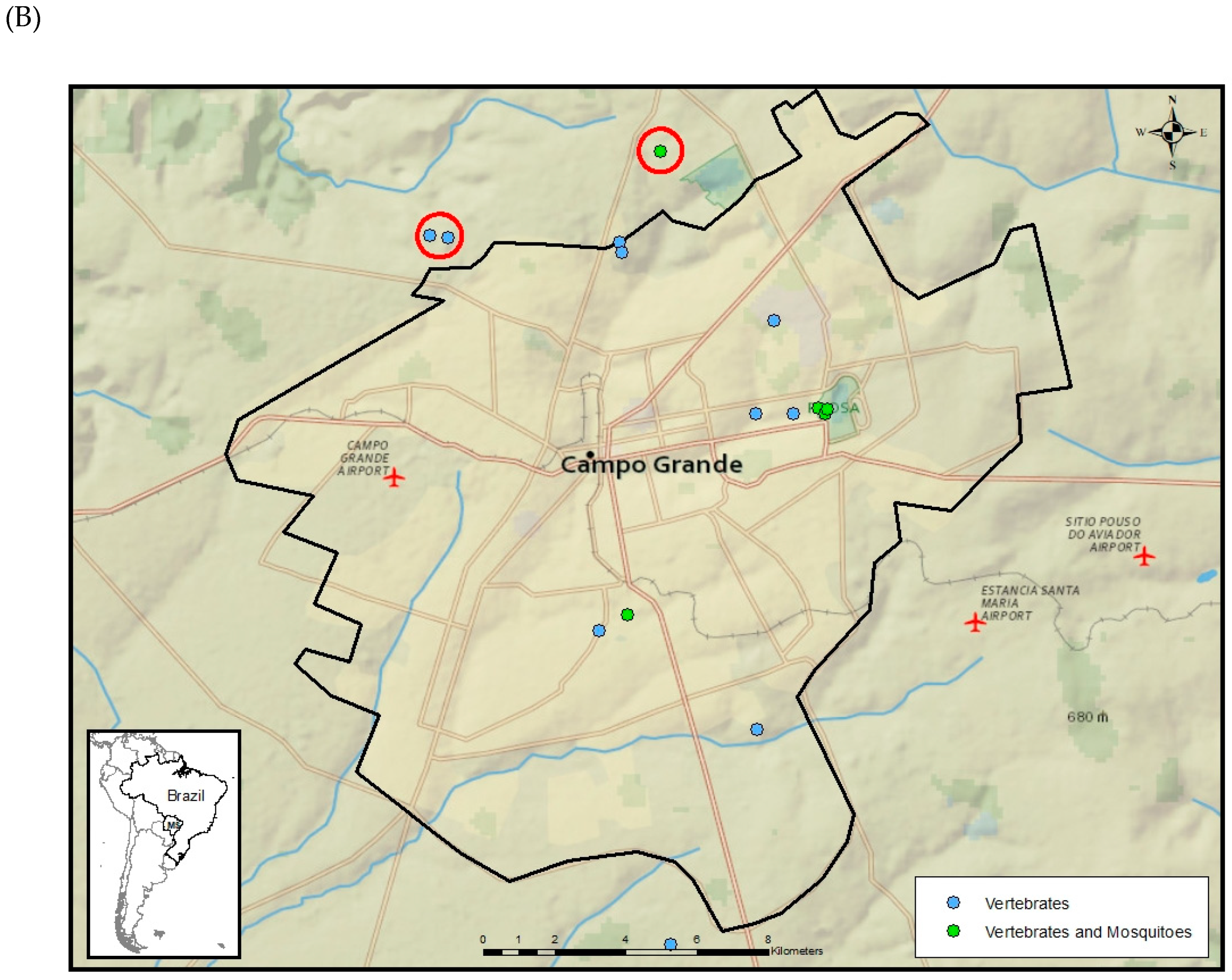
2.1. Study Sites
2.2. Mosquito Sampling
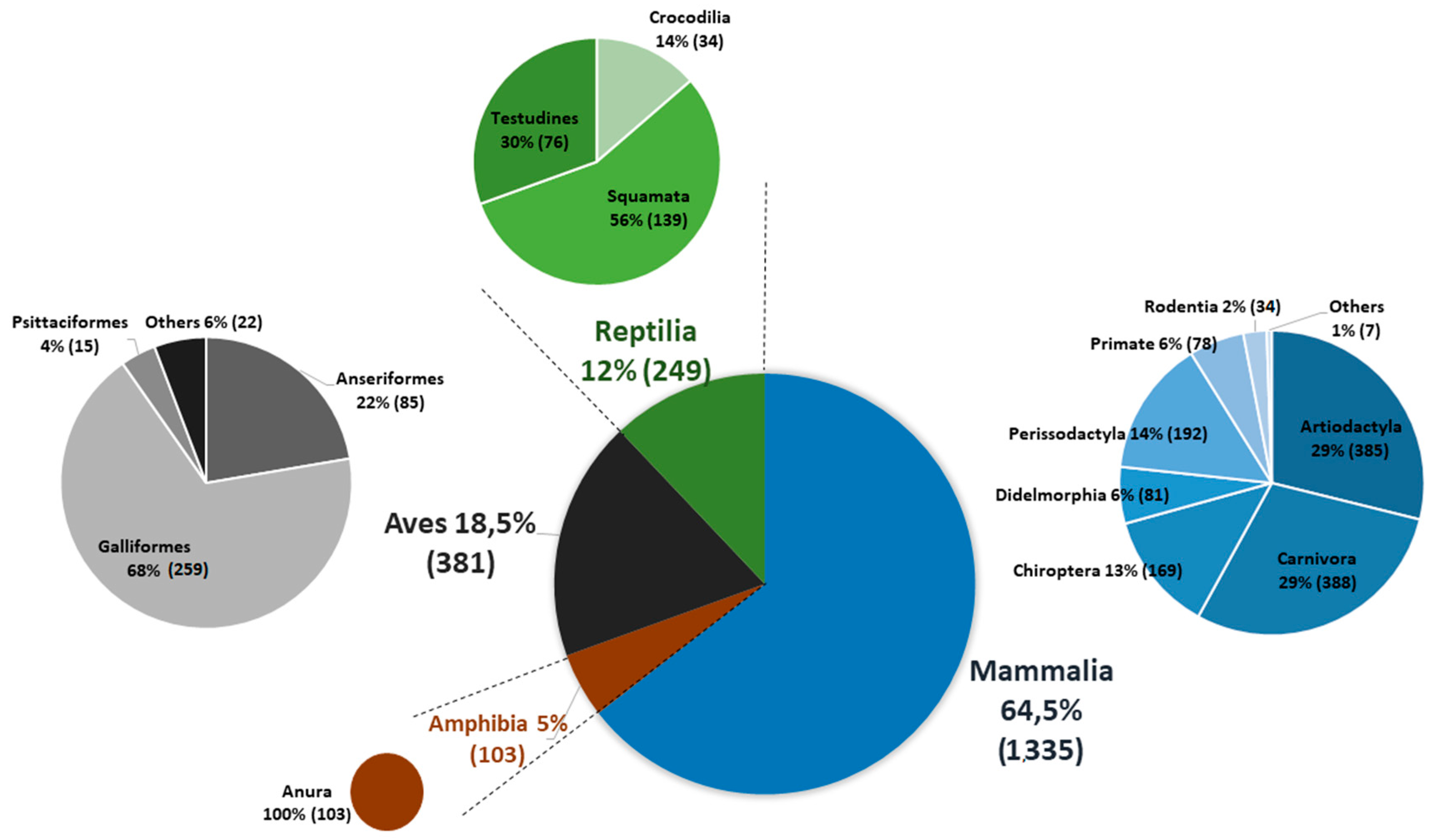
2.3. Vertebrate Sampling
2.4. Data Collection and Analysis
2.5. Laboratory Testing
2.5.1. Real-Time Reverse Transcriptase Polymerase Chain Reaction (RT-PCR) for Flaviviruses
2.5.2. Real-Time RT-PCR for ZIKV and Sanger Nucleotide Sequencing
2.5.3. Plaque Reduction Neutralization Test (PRNT90)
2.6. Ethics Clearance
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimers
References
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika Virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 10, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Ayres, C. Identification of Zika virus vectors and implications for control. Lancet Infect. Dis. 2016, 16, 278–279. [Google Scholar] [CrossRef]
- Gubler, D.J.; Vasilakis, N.; Musso, D. History and emergence of Zika virus. J. Infect. Dis. 2017, 216, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Campos, G.S.; Bandeira, A.C.; Sardi, S.I. Zika Virus Outbreak, Bahia, Brazil. Emerg. Infect. Dis. 2015, 21, 1885–1886. [Google Scholar] [CrossRef]
- Karabatsos, N. International Catalogue of Arboviruses Including Certain other Viruses of Vertebrates, 3rd ed.; American Society of Tropical Medicine & Hygiene: San Antonio, TX, USA, 1985; p. 1147. [Google Scholar]
- Vorou, R. Zika virus, vectors, reservoirs, amplifying hosts, and their potential to spread worldwide: What we know and what we should investigate urgently. Int. J. Infect. Dis. 2016, 48, 85–90. [Google Scholar] [CrossRef]
- Faye, O.; Faye, O.; Diallo, D.; Diallo, M.; Weidmann, M.; Sall, A.A. Quantitative real-time PCR detection of Zika virus and evaluation with field-caught Mosquitoes. Virol. J. 2013, 10, 311. [Google Scholar] [CrossRef]
- Olson, J.G.; Ksiazek, T.G.; Gubler, D.J.; Lubis, S.I.; Simanjuntak, G.; Lee, V.H.; Nalim, S.; Juslis, K.; See, R. A survey for arboviral antibodies in sera of humans and animals in Lombok, Republic of Indonesia. Ann. Trop. Med. Parasitol. 1983, 77, 131–137. [Google Scholar] [CrossRef]
- Darwish, M.A.; Hoogstraal, H.; Roberts, T.J.; Ahmed, I.P.; Omar, F. A sero-epidemiological survey for certain arboviruses (Togaviridae) in Pakistan. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 442–445. [Google Scholar] [CrossRef]
- Wolfe, N.D.; Kilbourn, A.M.; Karesh, W.B.; Rahman, H.A.; Bosi, E.J.; Cropp, B.C.; Andau, M.; Spielman, A.; Gubler, D.J. Sylvatic transmission of arboviruses among Bornean orangutans. Am. J. Trop. Med. Hyg. 2001, 64, 310–316. [Google Scholar] [CrossRef]
- Foy, B.D.; Kobylinski, K.C.; Chilson Foy, J.L.; Blitvich, B.J.; Travassos da Rosa, A.; Haddow, A.D.; Lanciotti, R.S.; Tesh, R.B. Probable non-vector-borne transmission of Zika virus, Colorado, USA. Emerg. Infect. Dis. 2011, 7, 880–882. [Google Scholar] [CrossRef]
- Oliveira Melo, A.S.; Malinger, G.; Ximenes, R.; Szejnfeld, P.O.; Alves Sampaio, S.; Bispo de Filippis, A.M. Zika virus intrauterine infection causes fetal brain abnormality and microcephaly: Tip of the iceberg? Ultrasound Obstet. Gynecol. 2016, 47, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Schuler-Faccini, L.; Ribeiro, E.M.; Feitosa, I.M.; Horovitz, D.D.; Cavalcanti, D.P.; Pessoa, A.; Doriqui, M.J.; Neri, J.I.; Neto, J.M.; Wanderley, H.Y.; et al. Brazilian Medical Genetics Society–Zika Embryopathy Task Force. Possible association between Zika virus infection and microcephaly—Brazil, 2015. MMWR 2016, 65, 59–62. [Google Scholar] [PubMed]
- Calvet, G.; Aguiar, R.S.; Melo, A.S.O.; Sampaio, S.A.; Filippis, I.; Fabri, A.; Araujo, E.S.M.; Sequeira, P.C.; Mendonça, M.C.L.; Oliveira, L.; et al. Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: A case study. Lancet Infect. Dis. 2016, 16, 653–660. [Google Scholar] [CrossRef]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodušek, V.; et al. Zika virus associated with microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Garcez, P.P.; Loiola, E.C.; Madeiro da Costa, R.; Higa, L.M.; Trindade, P.; Delvecchio, R.; Nascimento, J.M.; Brindeiro, R.; Tanuri, A.; Rehen, S.K. Zika virus impairs growth in human neurospheres and brain organoids. Science 2016, 352, 816–818. [Google Scholar] [CrossRef]
- Pierson, T.C.; Diamond, M.S. The emergence of Zika virus and its new clinical syndromes. Nature 2018, 560, 573–581. [Google Scholar] [CrossRef]
- Faria, N.R.; Avezedo, R.S.S.; Kraemer, M.U.G.; Souza, R.; Cunha, M.S.; Hill, S.C.; Thézé, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika virus in the America: Early epidemiological and genetic findings. Science 2016, 352, 345–349. [Google Scholar] [CrossRef]
- Figueiredo, L.T.M. Human Urban Arboviruses Can Infect Wild Animals and Jump to Sylvatic Maintenance Cycles in South America. Front. Cell Infect. Microbiol. 2019, 9, 259. [Google Scholar] [CrossRef]
- Moreira-Soto, A.; Carneiro, I.O.; Fischer, C.; Feldmann, M.; Kümmerer, B.M.; Silva, N.S.; Santos, U.G.; Souza, B.F.C.D.; Liborio, F.A.; Valença-Montenegro, M.M.; et al. Limited Evidence for Infection of Urban and Peri-urban Nonhuman Primates with Zika and Chikungunya Viruses in Brazil. mSphere 2018, 3, e00523-17. [Google Scholar] [CrossRef]
- Favoretto, S.R.; Araujo, D.B.; Duarte, N.F.H.; Oliveira, D.B.L.; da Crus, N.G.; Mesquita, F.; Leal, F.; Machado, R.R.G.; Gaio, F.; Oliveira, W.F.; et al. Zika Virus in Peridomestic Neotropical Primates, Northeast Brazil. Ecohealth 2019, 16, 61–69. [Google Scholar] [CrossRef]
- Terzian, A.C.B.; Zini, N.; Sacchetto, L.; Rocha, R.F.; Parra, M.C.P.; Del Sarto, J.L.; Dias, A.C.F.; Coutinho, F.; Rayra, J.; da Silva, R.A.; et al. Evidence of natural Zika virus infection in neotropical non-human primates in Brazil. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health (Ministério da Saúde). Boletim Epidemiológico 2. 2018; Volume 49. Available online: https://portalarquivos2.saude.gov.br/images/pdf/2018/janeiro/23/Boletim-2018-001-Dengue.pdf (accessed on 23 January 2018).
- Ministry of Health (Ministério da Saúde). Boletim Epidemiológico 22. 2019; Volume 50. Available online: https://portalarquivos2.saude.gov.br/images/pdf/2019/setembro/11/BE-arbovirose-22.pdf (accessed on 11 September 2019).
- Instituto Brasileiro de Geografia e Estatística. 2019. Available online: https://www.ibge.gov.br/cidades-e-estados/mt/varzea-grande.html (accessed on 1 October 2019).
- Lane, J. Neotropical Culicidae; Editora Universidade de São Paulo: São Paulo, Brazil, 1953; Volume 2, p. 1660. [Google Scholar]
- Forattini, O.P. Culicidologia médica: Identificação, Biologia, Epidemiologia; Editora da Universidade de São Paulo: São Paulo, Brazil, 2002; p. 860. [Google Scholar]
- Moureau, G.; Temmam, S.; Gonzalez, J.P.; Charrel, R.N.; Grard, G.; de Lamballerie, X. A real-time RT-PCR method for the universal detection and identification of flaviviruses. Vector Borne Zoonotic Dis. 2007, 7, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Pauvolid-Corrêa, A.; Campos, Z.; Juliano, R.; Velez, J.; Nogueira, R.M.R.; Komar, N. Serological evidence of widespread circulation of West Nile virus and other flaviviruses in Pantanal equines, Brazil. PLoS Negl. Trop. Dis. 2014, 8, e2706. [Google Scholar]
- Pauvolid-Corrêa, A.; Solberg, O.; Couto-Lima, D.; Kenney, J.; Serra-Freire, N.; Brault, A.; Nogueira, R.; Langevin, S.; Komar, N. Nhumirim virus, a novel flavivirus isolated from mosquitoes from the Pantanal, Brazil. Arch. Virol. 2015, 160, 21–27. [Google Scholar] [CrossRef]
- Beaty, B.; Calisher, C.H.; Shope, R.E. Arboviruses. In Viral, Rickettsial, and Chlamydial Infections; Lennette, E.H., Schmidt, N.J., Eds.; American Public Health Association: Washington, DC, USA, 1995; pp. 189–212. [Google Scholar]
- Pauvolid-Corrêa, A.; Campos, Z.; Soares, R.; Nogueira, R.M.R.; Komar, N. Neutralizing antibodies for orthobunyaviruses in Pantanal, Brazil. PLoS Negl. Trop. Dis. 2017, 11, e0006014. [Google Scholar]
- Mills, J.N.; Yates, T.L.; Childs, J.E.; Parmenter, R.E.; Ksiazek, T.G.; Rollin, P.E.; Peters, C.J. Guidelines for Working With Rodents Potentially Infected With Hantavirus. J. Mammal. 1995, 76, 716–722. [Google Scholar] [CrossRef]
- Sikes, R.S.; Gannon, W.L. Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 2011, 92, 235–253. [Google Scholar] [CrossRef]
- Sang, R.; Gichogo, A.; Gachoya, J.; Dunster, M.D.; Ofula, V.; Hunt, A.R.; Crabtree, M.B.; Miller, B.R.; Dunster, L.M. Isolation of a new flavivirus related to Cell fusing agent virus (CFAV) from field-collected flood-water Aedes mosquitoes sampled from a dambo in central Kenya. Arch. Virol. 2003, 148, 1085. [Google Scholar] [CrossRef]
- L’vov, D.K.; Al’khovskiĭ, S.V.; Shchelkanov, M.; Deriabin, P.G.; Shchetinin, A.M.; Samokhvalov, E.I.; Aristova, V.A.; Gitel’man, A.K.; Botikov, A.G. Genetic characterization of the Geran virus (GERV, Bunyaviridae, Nairovirus, Qalyub group) isolated from the ticks Ornithodoros verrucosus Olenev, Zasukhin and Fenyuk, 1934 (Argasidae) collected in the burrow of Meriones erythrourus Grey, 1842 in Azerbaijan. Vopr. Virusol. 2014, 59, 13–18. [Google Scholar]
- Ferreira, C.M.M.; Fischer, E.; Pulcherio-Leite, A. Fauna de morcegos em remanescentes urbanos de Cerrado em Campo Grande, Mato Grosso do Sul. Biota Neotrop. 2010, 10, 155–160. [Google Scholar] [CrossRef]
- Machado, N.G.; Rocha, L.A.; Silva, N.D.; Silva, D.F.N.; Florêncio, F.P.; Fernandes, T.B.S. Biodiversity in urban green space: A case study in the neotropics. Nativa 2017, 5, 320–329. [Google Scholar] [CrossRef]
- Ministry of Health (Ministério da Saúde). Boletim Epidemiológico 4. 2019; Volume 50. Available online: https://portalarquivos2.saude.gov.br/images/pdf/2019/janeiro/28/2019-002.pdf (accessed on 28 January 2019).
- Barrett, A.D.T.; Monath, T.P. Epidemiology and ecology of yellow fever virus. Adv. Virus. Res. 2003, 61, 291–315. [Google Scholar] [PubMed]
- Calisher, C.H.; Karabatsos, N.; Dalrymple, J.M.; Shope, R.E.; Porterfield, J.S.; Westaway, E.G.; Brandt, W.E. Antigenic relationships between flaviviruses as determined by cross neutralization tests with polyclonal antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.W.; Kosoy, O.; Hunsperger, E.; Beltran, M.; Delorey, M.; Guirakhoo, F.; Monath, T. Evaluation of chimeric Japanese encephalitis and Dengue viruses for use in diagnostic plaque reduction neutralization tests. Clin. Vaccine Immunol. 2009, 16, 1052–1059. [Google Scholar] [CrossRef] [PubMed]
- Iwashita, H.; Higa, Y.; Futami, K.; Lutiali, P.A.; Njenga, S.M.; Nabeshima, T.; Minakawa, N. Mosquito arbovirus survey in selected areas of Kenya: Detection of insect-specific virus. Trop. Med. Health 2018, 46, 19. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | MT | MS | Total | MT | MS | ||
|---|---|---|---|---|---|---|---|
| Vertebrate Species | N | N (%) | N (%) | Mosquito Species | N | N (%) | N (%) |
| Gallus gallus domesticus | 226 | 154 (68) | 72 (32) | Culex spp. | 21,207 | 19,215 (91) | 1992 (9) |
| Canis lupus familiaris | 190 | 79 (42) | 111 (58) | Aedes (Stegomyia) aegypti | 479 | 430 (90) | 49 (10) |
| Equus ferus caballus | 189 | 90 (48) | 99 (52) | Aedes (Stegomyia) albopictus | 343 | 311 (91) | 32 (9) |
| Bos indicus/taurus | 177 | 64 (36) | 113 (64) | Culex (Culex) quinquefasciatus | 222 | 222 (100) | 0 (0) |
| Ovis aries | 170 | 53 (31) | 117 (69) | Psorophora (Grabhamia) dimidiata | 197 | 83 (42) | 114 (58) |
| Felis silvestris catus | 102 | 52 (51) | 50 (49) | Culex (Culex) nigripalpis | 175 | 175 (100) | 0 (0) |
| Rhinella diptycha | 92 | 57 (62) | 35 (38) | Wyeomyia spp. | 167 | 167 (100) | 0 (0) |
| Nasua nasua | 86 | 4 (5) | 82 (95) | Aedes (Ochlerotatus) scapularis | 148 | 55 (37) | 93 (63) |
| Didelphis albiventris | 72 | 56 (78) | 16 (22) | Anopheles spp. | 143 | 77 (54) | 66 (46) |
| Anser anser domesticus | 63 | 63 (100) | 0 (0) | Psorophora (Janthinosoma) albigenu | 139 | 139 (100) | 0 (0) |
| Phrynops geoffroanus | 49 | 4 (8) | 45 (92) | Psorophora (Janthinosoma) ferox | 114 | 114 (100) | 0 (0) |
| Mico melanurus | 48 | 48 (100) | 0 (0) | Psorophora spp. | 69 | 39 (56,5) | 30 (43,5) |
| Crotalus durissus | 48 | 0 (0) | 48 (100) | Aedes spp. | 65 | 26 (40) | 39 (60) |
| Artibeus lituratus | 38 | 19 (50) | 19 (50) | Anopheles (Nyssorhynchus) rangeli | 64 | 64 (100) | 0 (0) |
| Artibeus planirostris | 38 | 5 (13) | 33 (87) | Deinocerites spp. | 64 | 64 (100) | 0 (0) |
| Sus scrofa domesticus | 36 | 36 (100) | 0 (0) | Limatus spp. | 58 | 58 (100) | 0 (0) |
| Carollia perspicillata | 36 | 21 (58) | 15 (42) | Anopheles (Stethomyia) kompi | 55 | 0 (0) | 55 (100) |
| Bothrops moojeni | 36 | 0 (0) | 36 (100) | Haemagogus (Haemagogus) janthinomys | 50 | 0 (0) | 50 (100) |
| Caiman yacare | 34 | 34 (100) | 0 (0) | Mansonia spp. | 49 | 49 (100) | 0 (0) |
| Bothrops alternatus | 24 | 0 (0) | 24 (100) | Psorophora (Psorophora) cilipes | 37 | 0 (0) | 37 (100) |
| Hydrochoerus hydrochaeris | 23 | 1 (4) | 22 (96) | Psorophora (Grabhamia) cingulata | 35 | 0 (0) | 35 (100) |
| Phasianus colchicus | 17 | 17 (100) | 0 (0) | Uranotaenia spp. | 30 | 30 (100) | 0 (0) |
| Anopheles (Nyssorhynchus) benarrochi | 29 | 0 (0) | 29 (100) |
| Class | Order | Species | ID | State | ZIKV | YFV | DENV-2 | WNV | Monotypic |
|---|---|---|---|---|---|---|---|---|---|
| Mammalia | Primates | Ateles marginatus | AU0002 | MT | 80 | <10 | <10 | <10 | YES |
| Mammalia | Perissodactyla | Equus ferus caballus | AU0194 | MT | 40 | <10 | <10 | <10 | YES |
| Mammalia | Artiodactyla | Ovis aries | AU0199 | MT | 40 | <10 | <10 | <10 | YES |
| Mammalia | Artiodactyla | Bos indicus/taurus | AU0219 | MT | 20 | <10 | <10 | <10 | YES |
| Mammalia | Artiodactyla | Bos indicus/taurus | AU0274 | MT | 20 | <10 | <10 | <10 | YES |
| Mammalia | Carnivora | Canis lupus familiaris | AU0063 | MT | 20 | <10 | <10 | <10 | YES |
| Mammalia | Perissodactyla | Equus ferus caballus | AU0137 | MT | 20 | <10 | <10 | <10 | YES |
| Mammalia | Perissodactyla | Equus ferus caballus | AU0188 | MT | 20 | <10 | <10 | <10 | YES |
| Mammalia | Perissodactyla | Equus ferus caballus | AU0192 | MT | 20 | <10 | <10 | <10 | YES |
| Mammalia | Artiodactyla | Ovis aries | AU0205 | MT | 20 | <10 | <10 | <10 | YES |
| Aves | Anseriformes | Anser anser domesticus | AU0439 | MT | 20 | <10 | <10 | <10 | YES |
| Aves | Anseriformes | Anser anser domesticus | AU0440 | MT | 20 | <10 | <10 | <10 | YES |
| Aves | Anseriformes | Anser anser domesticus | AU0445 | MT | 20 | <10 | <10 | <10 | YES |
| Aves | Anseriformes | Anser anser domesticus | AU0447 | MT | 20 | <10 | <10 | <10 | YES |
| Aves | Galliformes | Gallus gallus domesticus | AU0008 | MT | 20 | <10 | <10 | <10 | YES |
| Aves | Galliformes | Gallus gallus domesticus | AU0029 | MT | 20 | <10 | <10 | <10 | YES |
| Aves | Galliformes | Gallus gallus domesticus | AU0090 | MT | 20 | <10 | <10 | <10 | YES |
| Mammalia | Artiodactyla | Bos indicus/taurus | AG0329 | MS | 40 | <10 | <10 | <10 | YES |
| Mammalia | Artiodactyla | Bos indicus/taurus | AG0340 | MS | 20 | <10 | <10 | <10 | YES |
| Mammalia | Artiodactyla | Bos indicus/taurus | AG0348 | MS | 20 | <10 | <10 | <10 | YES |
| Mammalia | Carnivora | Canis lupus familiaris | AG0007 | MS | 20 | <10 | <10 | <10 | YES |
| Mammalia | Carnivora | Canis lupus familiaris | AG0019 | MS | 20 | <10 | <10 | <10 | YES |
| Aves | Galliformes | Gallus gallus domesticus | AG0379 | MS | 20 | <10 | <10 | <10 | YES |
| Mammalia | Artiodactyla | Bos indicus/taurus | AG0346 | MS | 20 | ≥10 | ≥10 | <10 | NO |
| Mammalia | Primates | Sapajus cay | AG0297 | MS | 160 | <10 | ≥10 | <10 | NO |
| Mammalia | Perissodactyla | Equus ferus caballus | AU0366 | MT | 40 | ≥10 | ≥10 | ≥10 | NO |
| Mammalia | Rodentia | Dasyprocta azarae | AU0500 | MT | 80 | <10 | ≥10 | <10 | NO |
| Mammalia | Perissodactyla | Equus ferus caballus | AU0184 | MT | 80 | <10 | <10 | ≥10 | NO |
| Mammalia | Carnivora | Cerdocyon thous | AU0197 | MT | 160 | <10 | ≥10 | <10 | NO |
| Mammalia | Carnivora | Canis lupus familiaris | AU0257 | MT | 1280 | <10 | <10 | ≥10 | NO |
| Mammalia | Chiroptera | Artibeus planirostris | AEU042 | MT | 2560 | <10 | ≥10 | ≥10 | NO |
| Mammalia | Primates | Ateles marginatus | AU0001 | MT | ≥320 | <10 | ≥10 | <10 | NO |
| Aves | Anseriformes | Anser anser domesticus | AU0438 | MT | 20 | <10 | ≥10 | <10 | NO |
| Aves | Galliformes | Gallus gallus domesticus | AU0375 | MT | 80 | <10 | <10 | ≥10 | NO |
| Species | Total | MT | MS | ||||
|---|---|---|---|---|---|---|---|
| Scientific Name | Common Name | N | M (%) | N | M (%) | N | M (%) |
| Anser anser domesticus | Domestic graylag goose | 37 | 4 (10.8) | 37 | 4 (10.8) | 0 | _ |
| Bos indicus/taurus | Cattle | 171 | 5 (2.9) | 59 | 2 (3.4) | 112 | 3 (2.7) |
| Gallus gallus domesticus | Chicken | 145 | 4 (2.8) | 102 | 3 (2.9) | 43 | 1 (2.3) |
| Equus ferus caballus | Horse | 187 | 4 (2.1) | 88 | 4 (4.5) | 99 | 0 |
| Canis lupus familiaris | Dog | 159 | 3 (1.9) | 48 | 1 (2.1) | 111 | 2 (1.8) |
| Ovis aries | Sheep | 164 | 2 (1.2) | 49 | 2 (4.1) | 115 | 4 (3.5) |
| Nasua nasua | South American coati | 82 | 0 | 2 | 0 | 80 | 0 |
| Felis catus | Cat | 66 | 0 | 23 | 0 | 43 | 0 |
| Phrynops geoffroanus | Geoffroy’s side-necked turtle | 41 | 0 | 0 | _ | 41 | 0 |
| Didelphis albiventris | White-eared opossum | 38 | 0 | 27 | 0 | 11 | 0 |
| Artibeus lituratus | Great fruit-eating bat | 32 | 0 | 16 | 0 | 16 | 0 |
| Artibeus planirostris | Flat-faced fruit-eating bat | 32 | 0 | 5 | 0 | 27 | 0 |
| Crotalus durissus | South American rattlesnake | 32 | 0 | 0 | _ | 32 | 0 |
| Sus scrofa domesticus | Pig | 32 | 0 | 32 | 0 | 0 | _ |
| Mico melanurus | Black-tailed marmoset | 31 | 0 | 31 | 0 | 0 | _ |
| Rhinella diptycha | Cope’s toad | 26 | 0 | 13 | 0 | 13 | 0 |
| Carollia perspicillata | Seba’s short-tailed bat | 23 | 0 | 11 | 0 | 12 | 0 |
| Bothrops moojeni | Moojen’s lancehed | 22 | 0 | 0 | _ | 22 | 0 |
| Caiman yacare | Yacare caiman | 22 | 0 | 22 | 0 | 0 | _ |
| Hydrochoerus hydrochaeris | Capybara | 21 | 0 | 0 | _ | 21 | 0 |
| Bothrops alternatus | Urutu lancehead | 15 | 0 | 0 | _ | 15 | 0 |
| Sapajus cay | Hooded capuchin | 12 | 0 | 0 | _ | 12 | 0 |
| All other species * | General | 108 | 0 | 74 | 0 | 34 | 0 |
| Pool ID | State | Species | n | Coverage/Identity | Virus Sequences | Genbank# |
|---|---|---|---|---|---|---|
| P0048 | MS | Sabethes sp. | 1 | 34%/95.7% | Mosquito flavivirus | HQ676625.1 |
| P0049 | MS | Sabethes sp. | 1 | 84%/92.2% | Mosquito flavivirus | HQ676625.1 |
| P0057 | MS | Aedes (Stegomyia) aegypti | 2 | 50%/87.4% | Mosquito flavivirus | HQ676625.1 |
| P0217 | MS | Aedes (Stegomyia) aegypti | 4 | 78%/84.5% | Mosquito flavivirus | HQ676625.1 |
| PU0026 | MT | Aedes sp. | 3 | 77%/95.0% | Mosquito flavivirus | HQ676624.1 |
| PU0064 | MT | Culex sp. | 12 | 46%/80.0% | Kamiti river virus | AY149904.1 |
| PU0196 | MT | Culex sp. | 25 | 87%/92.0% | Mosquito flavivirus | HQ676625.1 |
| PU0259 | MT | Limatus sp. | 23 | 77%/93.0% | Mosquito flavivirus | HQ676625.1 |
| PU0261 | MT | Limatus sp. | 25 | 86%/90.0% | Mosquito flavivirus | HQ676625.1 |
| PU0330 | MT | Wyeomyia sp. | 25 | 87%/93.9% | Mosquito flavivirus | HQ676625.1 |
| PU0806 | MT | Aedes (Stegomyia) aegypti | 18 | 54%/78.3% | Kamiti river virus | DQ335465.1 |
| PU0807 | MT | Aedes (Stegomyia) aegypti | 25 | 77%/89.2% | Mosquito flavivirus | HQ676625.1 |
| PU0808 | MT | Aedes (Stegomyia) aegypti | 25 | 78%/91.4% | Mosquito flavivirus | HQ676625.1 |
| PU0852 | MT | Aedes sp. | 2 | 78%/86.8% | Mosquito flavivirus | HQ676625.1 |
| PU0853 | MT | Aedes sp. | 2 | 80%/85.9% | Mosquito flavivirus | HQ676625.1 |
| PU0874 | MT | Psorophora (Janthinosoma) albigenu | 25 | 37%/92.6% | Geran virus | KP792714.1 |
| PU1001 | MT | Aedes (Stegomyia) aegypti | 1 | 94%/94.7% | Mosquito flavivirus | HQ676625.1 |
| PU1007 | MT | Aedes (Stegomyia) aegypti | 2 | 80%/89.2% | Mosquito flavivirus | HQ676625.1 |
| PU1031 | MT | Aedes (Stegomyia) aegypti | 2 | 53%/80.8% | Kamiti river virus | DQ335465.1 |
| PU1032 | MT | Aedes (Stegomyia) aegypti | 15 | 82%/87.6% | Mosquito flavivirus | HQ676625.1 |
| PU1098 | MT | Aedes (Stegomyia) aegypti | 7 | 82%/87.6% | Mosquito flavivirus | HQ676625.1 |
| PU1124 | MT | Aedes (Stegomyia) aegypti | 6 | 86%/94.7% | Mosquito flavivirus | HQ676625.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pauvolid-Corrêa, A.; Gonçalves Dias, H.; Marina Siqueira Maia, L.; Porfírio, G.; Oliveira Morgado, T.; Sabino-Santos, G.; Helena Santa Rita, P.; Teixeira Gomes Barreto, W.; Carvalho de Macedo, G.; Marinho Torres, J.; et al. Zika Virus Surveillance at the Human–Animal Interface in West-Central Brazil, 2017–2018. Viruses 2019, 11, 1164. https://doi.org/10.3390/v11121164
Pauvolid-Corrêa A, Gonçalves Dias H, Marina Siqueira Maia L, Porfírio G, Oliveira Morgado T, Sabino-Santos G, Helena Santa Rita P, Teixeira Gomes Barreto W, Carvalho de Macedo G, Marinho Torres J, et al. Zika Virus Surveillance at the Human–Animal Interface in West-Central Brazil, 2017–2018. Viruses. 2019; 11(12):1164. https://doi.org/10.3390/v11121164
Chicago/Turabian StylePauvolid-Corrêa, Alex, Helver Gonçalves Dias, Laura Marina Siqueira Maia, Grasiela Porfírio, Thais Oliveira Morgado, Gilberto Sabino-Santos, Paula Helena Santa Rita, Wanessa Teixeira Gomes Barreto, Gabriel Carvalho de Macedo, Jaire Marinho Torres, and et al. 2019. "Zika Virus Surveillance at the Human–Animal Interface in West-Central Brazil, 2017–2018" Viruses 11, no. 12: 1164. https://doi.org/10.3390/v11121164
APA StylePauvolid-Corrêa, A., Gonçalves Dias, H., Marina Siqueira Maia, L., Porfírio, G., Oliveira Morgado, T., Sabino-Santos, G., Helena Santa Rita, P., Teixeira Gomes Barreto, W., Carvalho de Macedo, G., Marinho Torres, J., Arruda Gimenes Nantes, W., Martins Santos, F., Oliveira de Assis, W., Castro Rucco, A., Mamoru dos Santos Yui, R., Bosco Vilela Campos, J., Rodrigues Leandro e Silva, R., da Silva Ferreira, R., Aparecido da Silva Neves, N., ... Komar, N. (2019). Zika Virus Surveillance at the Human–Animal Interface in West-Central Brazil, 2017–2018. Viruses, 11(12), 1164. https://doi.org/10.3390/v11121164