Emergence of Fluoxetine-Resistant Variants during Treatment of Human Pancreatic Cell Cultures Persistently Infected with Coxsackievirus B4

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Reagents
2.2. Virus and Persistent Infection
2.3. Antiviral Activity Testing
2.4. Determination of Viral Titer
2.5. Viral Genome Sequencing
2.6. Statistical Analysis
3. Results
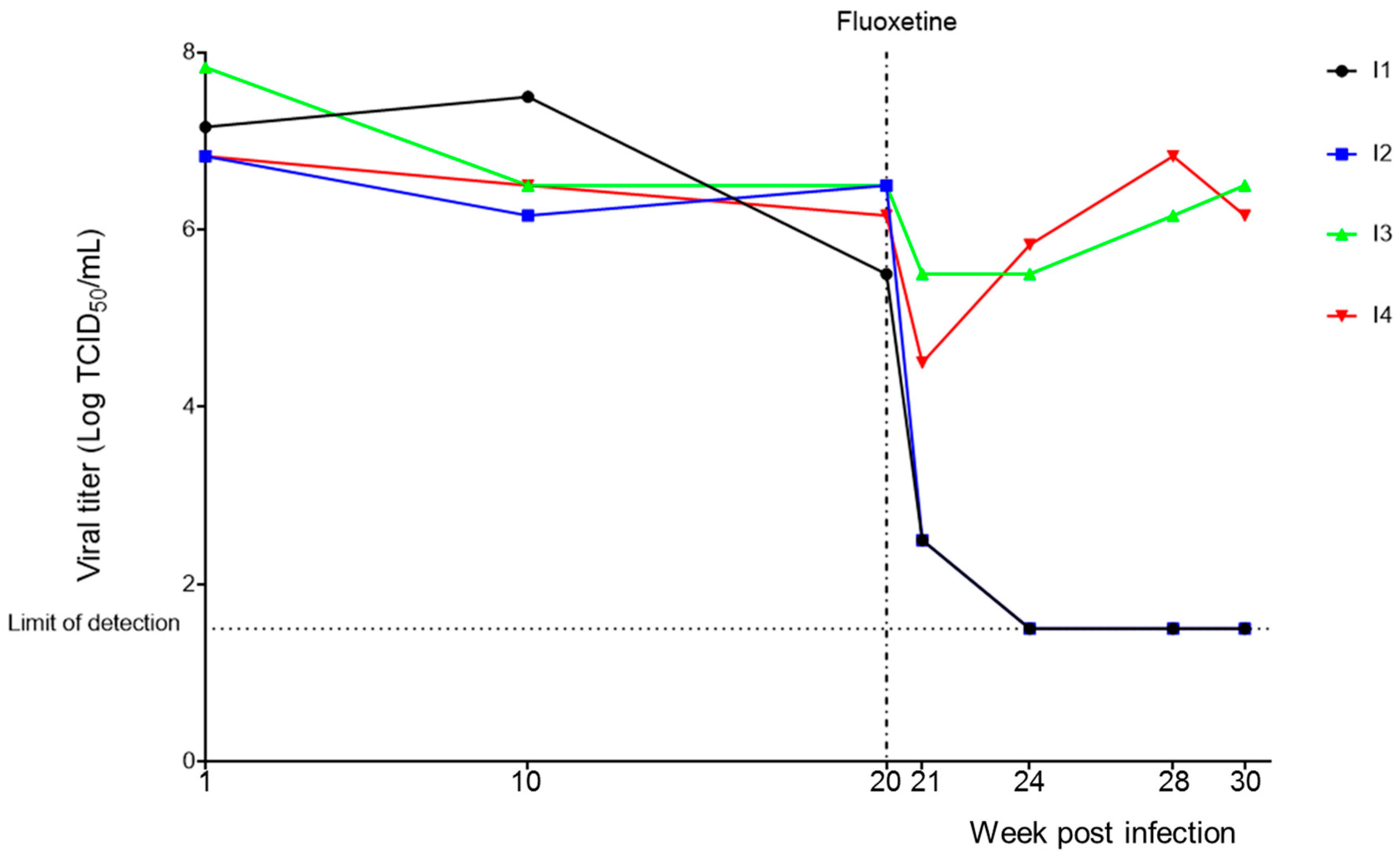
3.1. Persistent CVB4 Infection and Fluoxetine Treatment
3.2. Investigation of Resistance to Fluoxetine Treatment
3.3. Mutations in 2C Protein Associated with Resistance to Fluoxetine
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Knowles, N.; Hovi, T.; Hyypiä, T. Picornaviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 855–880. [Google Scholar]
- Romero, J.R. Pediatric group B coxsackievirus infections. Curr. Top. Microbiol. Immunol. 2008, 323, 223–239. [Google Scholar] [PubMed]
- Tapparel, C.; Siegrist, F.; Petty, T.J.; Kaiser, L. Picornavirus and enterovirus diversity with associated human diseases. Infect. Genet. Evol. 2013, 14, 282–293. [Google Scholar] [CrossRef]
- Chapman, N.M.; Kim, K.S. Persistent coxsackievirus infection: Enterovirus persistence in chronic myocarditis and dilated cardiomyopathy. Curr. Top. Microbiol. Immunol. 2008, 323, 275–292. [Google Scholar] [PubMed]
- Hober, D.; Alidjinou, E.K. Enteroviral pathogenesis of type 1 diabetes: Queries and answers. Curr. Opin. Infect. Dis. 2013, 26, 263–269. [Google Scholar] [CrossRef]
- Hober, D.; Sauter, P. Pathogenesis of type 1 diabetes mellitus: Interplay between enterovirus and host. Nat. Rev. Endocrinol. 2010, 6, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Alidjinou, E.K.; Sané, F.; Engelmann, I.; Geenen, V.; Hober, D. Enterovirus persistence as a mechanism in the pathogenesis of type 1 diabetes. Discov. Med. 2014, 18, 273–282. [Google Scholar]
- De Palma, A.M.; Vliegen, I.; De Clercq, E.; Neyts, J. Selective inhibitors of picornavirus replication. Med. Res. Rev. 2008, 28, 823–884. [Google Scholar] [CrossRef]
- Thibaut, H.J.; Leyssen, P.; Puerstinger, G.; Muigg, A.; Neyts, J.; De Palma, A.M. Towards the design of combination therapy for the treatment of enterovirus infections. Antiviral Res. 2011, 90, 213–217. [Google Scholar] [CrossRef]
- Ulferts, R.; van der Linden, L.; Thibaut, H.J.; Lanke, K.H.W.; Leyssen, P.; Coutard, B.; De Palma, A.M.; Canard, B.; Neyts, J.; van Kuppeveld, F.J.M. Selective serotonin reuptake inhibitor fluoxetine inhibits replication of human enteroviruses B and D by targeting viral protein 2C. Antimicrob. Agents Chemother. 2013, 57, 1952–1956. [Google Scholar] [CrossRef]
- Zuo, J.; Quinn, K.K.; Kye, S.; Cooper, P.; Damoiseaux, R.; Krogstad, P. Fluoxetine is a potent inhibitor of coxsackievirus replication. Antimicrob. Agents Chemother. 2012, 56, 4838–4844. [Google Scholar] [CrossRef]
- Alidjinou, E.K.; Sané, F.; Bertin, A.; Caloone, D.; Hober, D. Persistent infection of human pancreatic cells with Coxsackievirus B4 is cured by fluoxetine. Antiviral Res. 2015, 116, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Alidjinou, E.K.; Engelmann, I.; Bossu, J.; Villenet, C.; Figeac, M.; Romond, M.-B.; Sané, F.; Hober, D. Persistence of Coxsackievirus B4 in pancreatic ductal-like cells results in cellular and viral changes. Virulence 2017, 8, 1229–1244. [Google Scholar] [CrossRef] [PubMed]
- Sane, F.; Caloone, D.; Gmyr, V.; Engelmann, I.; Belaich, S.; Kerr-Conte, J.; Pattou, F.; Desailloud, R.; Hober, D. Coxsackievirus B4 can infect human pancreas ductal cells and persist in ductal-like cell cultures which results in inhibition of Pdx1 expression and disturbed formation of islet-like cell aggregates. Cell Mol. Life Sci. 2013, 70, 4169–4180. [Google Scholar] [CrossRef] [PubMed]
- Ulferts, R.; de Boer, S.M.; van der Linden, L.; Bauer, L.; Lyoo, H.R.; Maté, M.J.; Lichière, J.; Canard, B.; Lelieveld, D.; Omta, W.; et al. Screening of a Library of FDA-Approved Drugs Identifies Several Enterovirus Replication Inhibitors That Target Viral Protein 2C. Antimicrob. Agents Chemother. 2016, 60, 2627–2638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfister, T.; Jones, K.W.; Wimmer, E. A cysteine-rich motif in poliovirus protein 2C(ATPase) is involved in RNA replication and binds zinc in vitro. J. Virol. 2000, 74, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Singleton, M.R.; Dillingham, M.S.; Wigley, D.B. Structure and mechanism of helicases and nucleic acid translocases. Annu. Rev. Biochem. 2007, 76, 23–50. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Wang, P.; Wang, G.-C.; Yang, J.; Sun, X.; Wu, W.; Qiu, Y.; Shu, T.; Zhao, X.; Yin, L.; et al. Human Enterovirus Nonstructural Protein 2CATPase Functions as Both an RNA Helicase and ATP-Independent RNA Chaperone. PLoS Pathog. 2015, 11, e1005067. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.C. The not-so-infinite malleability of RNA viruses: Viral and cellular determinants of RNA virus mutation rates. PLoS Pathog. 2017, 13, e1006254. [Google Scholar] [CrossRef]
- De Palma, A.M.; Heggermont, W.; Lanke, K.; Coutard, B.; Bergmann, M.; Monforte, A.-M.; Canard, B.; De Clercq, E.; Chimirri, A.; Pürstinger, G.; et al. The thiazolobenzimidazole TBZE-029 inhibits enterovirus replication by targeting a short region immediately downstream from motif C in the nonstructural protein 2C. J. Virol. 2008, 82, 4720–4730. [Google Scholar] [CrossRef]
- Guan, H.; Tian, J.; Qin, B.; Wojdyla, J.A.; Wang, B.; Zhao, Z.; Wang, M.; Cui, S. Crystal structure of 2C helicase from enterovirus 71. Sci. Adv. 2017, 3, e1602573. [Google Scholar] [CrossRef]
- LLerena, A.; Dorado, P.; Berecz, R.; González, A.; Jesús Norberto, M.; de la Rubia, A.; Cáceres, M. Determination of fluoxetine and norfluoxetine in human plasma by high-performance liquid chromatography with ultraviolet detection in psychiatric patients. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2003, 783, 25–31. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward/Reverse | Primer Sequence (5′–3′) | Nucleotide Position |
|---|---|---|---|
| EXT-1 | Forward | CTCAAGCGGAAAGTGTCCCA | 3988–4007 |
| INT-1 | Reverse | TTTCCCATCAGGGTTCTGGC | 4593–4574 |
| INT-2 | Forward | GATTGGGCGTTCACTTGCAG | 4461–4480 |
| EXT-2 | Reverse | ACTGCCTCACTATCCACCGA | 5126–5107 |
| 2C Protein Sequences | Amino-Acid Positions | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 133 | 188 | 216 | 227 | 229 | 255 | 296 | ||||
| CVB4 E2 reference published strain (NCBI, accession: AF311939.1) | A | S | P | I | A | S | R | |||
| Laboratory CVB4 E2 stock strain | A | S | P | I | A | S | R | |||
| Sequences of isolates from CVB4 E2 persistently infected Panc-1 cells | Baseline samples (CVB4 E2 persistent infection, 20 weeks) | I1 | A | S | P | I | A | N | R | |
| I2 | A | S | S | I | A | S | R | |||
| I3 | A | S | P | I | A | S | G | |||
| I4 | A | S | P | I | A | S | G | |||
| 4 weeks post treatment | Fluoxetine | I1 | ND | ND | ND | ND | ND | ND | ND | |
| I2 | ND | ND | ND | ND | ND | ND | ND | |||
| I3 | A | S | P | I/V | A | S | G | |||
| I4 | A | S | P | I | A | S | G | |||
| DMSO | I1 | A | S | P | I | A | N | R | ||
| I2 | A | S | S | I | A | S | R | |||
| I3 | A | S | P | I | A | S | G | |||
| I4 | A | S | P | I | A | S | G | |||
| 10 weeks posttreatment | Fluoxetine | I1 | ND | ND | ND | ND | ND | ND | ND | |
| I2 | ND | ND | ND | ND | ND | ND | ND | |||
| I3 | T | S/A | P | V | A/V | S | G | |||
| I4 | A/T | S | P | V | A | S | G | |||
| DMSO | I1 | A | S | P | I | A | N | R | ||
| I2 | A | S | S | I | A | S | R | |||
| I3 | A | S | P | I | A | S | G | |||
| I4 | A | S | P | I | A | S | G | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alidjinou, E.K.; Bertin, A.; Sane, F.; Caloone, D.; Engelmann, I.; Hober, D. Emergence of Fluoxetine-Resistant Variants during Treatment of Human Pancreatic Cell Cultures Persistently Infected with Coxsackievirus B4. Viruses 2019, 11, 486. https://doi.org/10.3390/v11060486
Alidjinou EK, Bertin A, Sane F, Caloone D, Engelmann I, Hober D. Emergence of Fluoxetine-Resistant Variants during Treatment of Human Pancreatic Cell Cultures Persistently Infected with Coxsackievirus B4. Viruses. 2019; 11(6):486. https://doi.org/10.3390/v11060486
Chicago/Turabian StyleAlidjinou, Enagnon Kazali, Antoine Bertin, Famara Sane, Delphine Caloone, Ilka Engelmann, and Didier Hober. 2019. "Emergence of Fluoxetine-Resistant Variants during Treatment of Human Pancreatic Cell Cultures Persistently Infected with Coxsackievirus B4" Viruses 11, no. 6: 486. https://doi.org/10.3390/v11060486
APA StyleAlidjinou, E. K., Bertin, A., Sane, F., Caloone, D., Engelmann, I., & Hober, D. (2019). Emergence of Fluoxetine-Resistant Variants during Treatment of Human Pancreatic Cell Cultures Persistently Infected with Coxsackievirus B4. Viruses, 11(6), 486. https://doi.org/10.3390/v11060486