Seoul Virus Tropism and Pathology in Naturally Infected Feeder Rats

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Studies
2.3. Detection of SEOV Antibodies
2.4. Detection of Viral RNA
2.5. Pathology & Histology:
2.6. Statistics
3. Results
3.1. Clinical Signs
3.2. Laboratory Findings
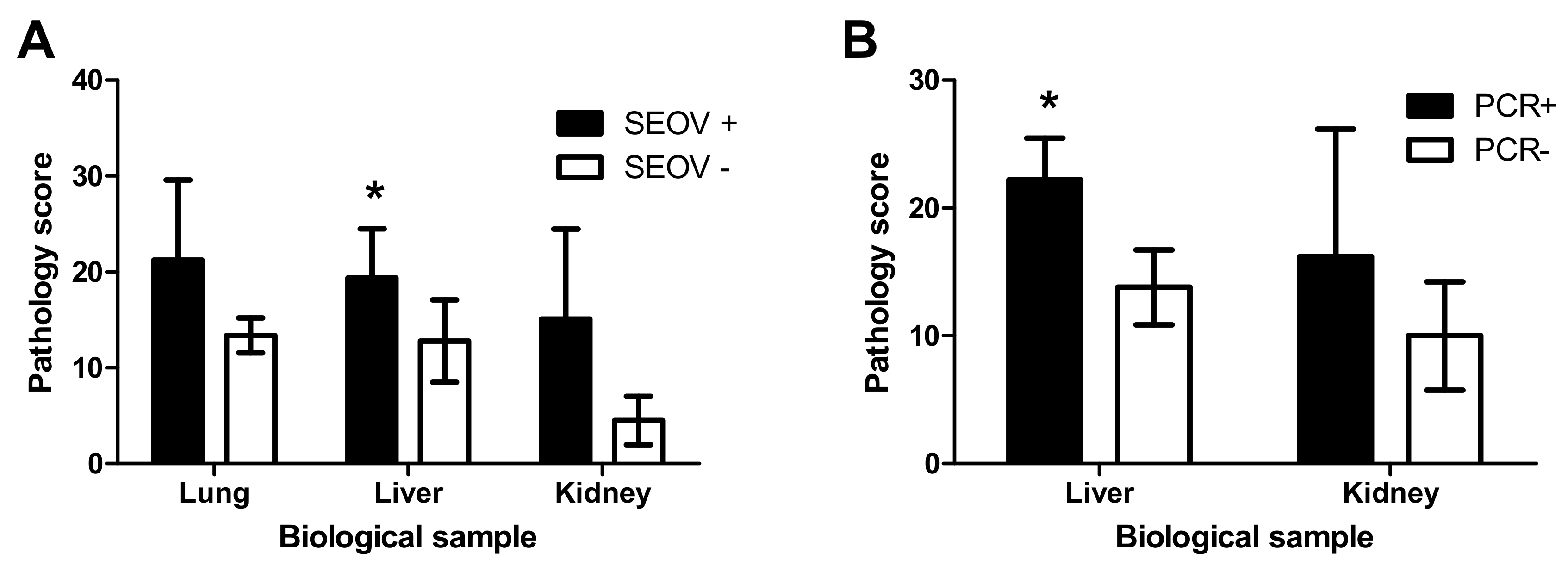
3.3. Pathological Findings Rats
3.3.1. Gross Lesions
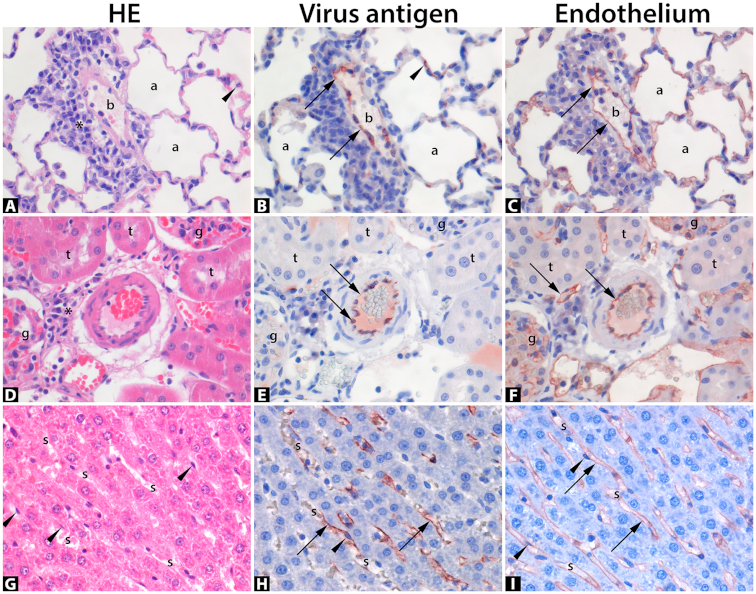
3.3.2. Histopathology in Lungs
3.3.3. Histopathology in Kidneys
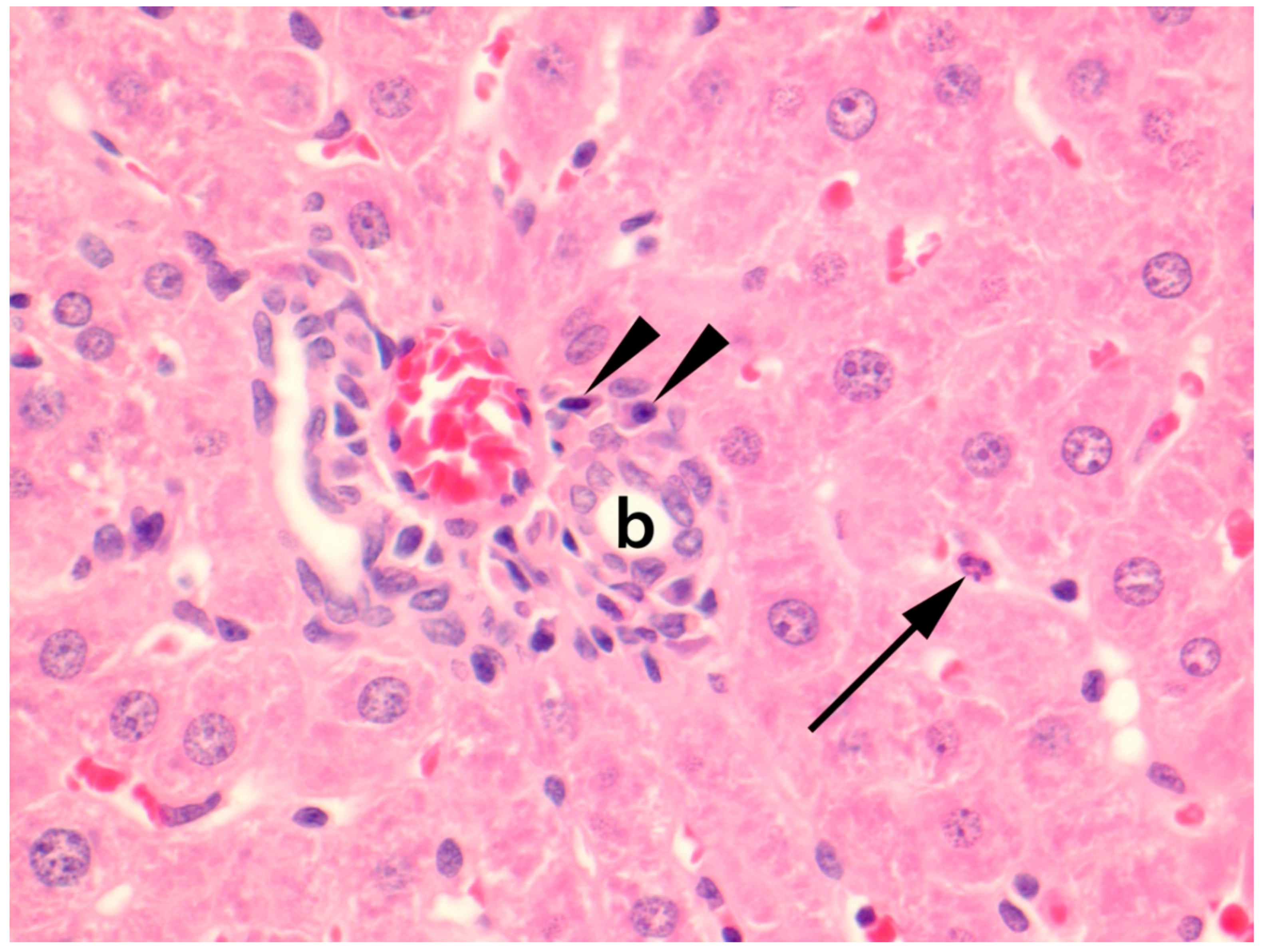
3.3.4. Histopathology in Liver
3.3.5. Other Histopathological Findings
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kruger, D.H.; Figueiredo, L.T.M.; Song, J.W.; Klempa, B.J. Hantaviruses—Globally emerging pathogens. J. Clin. Virol. 2015, 64, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Goeijenbier, M.; Verner-Carlsson, J.; van Gorp, E.C.; Rockx, B.; Koopmans, M.P.; Lundkvist, A.; van der Giessen, J.W.; Reusken, C.B. Seoul hantavirus in brown rats in the Netherlands: Implications for physicians—Epidemiology, clinical aspects, treatment and diagnostics. Neth. J. Med. 2015, 73, 155–160. [Google Scholar] [PubMed]
- Kim, W.K.; No, J.S.; Lee, S.H.; Song, D.H.; Lee, D.; Kim, J.A.; Gu, S.H.; Park, S.; Jeong, S.T.; Kim, H.C.; et al. Multiplex PCR-Based Next-Generation Sequencing and Global Diversity of Seoul Virus in Humans and Rats. Emerg. Infect. Dis. 2018, 24, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Hart, C.A.; Bennett, M. Hantavirus infections: Epidemiology and pathogenesis. Microbes Infect. 1999, 1, 1229–1237. [Google Scholar] [CrossRef]
- Glass, G.E.; Childs, J.; Korch, G.W.; LeDuc, J. Association of intraspecific wounding with hantaviral infection in wild rats (Rattus norvegicus). Epidemiol. Infect. 1988, 101, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Swanink, C.; Reimerink, J.; Gisolf, J.; de Vries, A.; Claassen, M.; Martens, L.; Waegemaekers, T.; Rozendaal, H.; Valkenburgh, S.; Hoornweg, T.; et al. Autochthonous Human Case of Seoul Virus Infection, the Netherlands. Emerg. Infect. Dis. 2018, 24, 2158–2163. [Google Scholar] [CrossRef] [PubMed]
- Kerins, J.L.; Koske, S.E.; Kazmierczak, J.; Austin, C.; Gowdy, K.; Dibernardo, A.; Group, C.S.V.I.; Group, C.S.V.I.; Morbidity, S.V.W.G.J.; Report, M.W. Outbreak of Seoul virus among rats and rat owners—United States and Canada, 2017. Morb. Mortal. Wkly. Rep. 2018, 67, 131. [Google Scholar] [CrossRef] [PubMed]
- Jameson, L.; Logue, C.; Atkinson, B.; Baker, N.; Galbraith, S.; Carroll, M.; Brooks, T.; Hewson, R. The continued emergence of hantaviruses: Isolation of a Seoul virus implicated in human disease, United Kingdom, October 2012. Eurosurveill 2013, 18, 20344. [Google Scholar]
- Mace, G.; Feyeux, C.; Mollard, N.; Chantegret, C.; Audia, S.; Rebibou, J.M.; Spagnolo, G.; Bour, J.B.; Denoyel, G.A.; Sagot, P.; et al. Severe Seoul hantavirus infection in a pregnant woman, France, October 2012. Eurosurveill 2013, 18, 20464. [Google Scholar]
- Verner-Carlsson, J.; Lohmus, M.; Sundstrom, K.; Strand, T.M.; Verkerk, M.; Reusken, C.; Yoshimatsu, K.; Arikawa, J.; van de Goot, F.; Lundkvist, A. First evidence of Seoul hantavirus in the wild rat population in the Netherlands. Infect. Ecol. Epidemiol. 2015, 5, 27215. [Google Scholar] [CrossRef] [PubMed]
- Maas, M.; De Vries, A.; Reusken, C.; Buijs, J.; Goris, M.; Hartskeerl, R.; Ahmed, A.; Van Tulden, P.; Swart, A.; Pijnacker, R.; et al. Prevalence of Leptospira spp. and Seoul hantavirus in brown rats (Rattus norvegicus) in four regions in the Netherlands, 2011–2015. Infect. Ecol. Epidemiol. 2018, 8, 1490135. [Google Scholar] [CrossRef] [PubMed]
- Friesema, I.H.M.; Bakker, J.; Maas, M.; Goris, M.G.A.; van der Giessen, J.W.B.; Rockx, B.H.G. Seroprevalence of hantaviruses and Leptospira in muskrat and coypu trappers in the Netherlands, 2016. Infect. Ecol. Epidemiol. 2018, 8, 1474707. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Prescott, J. Hantavirus immunology of rodent reservoirs: Current status and future directions. Viruses 2014, 6, 1317–1335. [Google Scholar] [CrossRef]
- Easterbrook, J.D.; Klein, S.L. Immunological mechanisms mediating hantavirus persistence in rodent reservoirs. PLoS Pathog. 2008, 4, e1000172. [Google Scholar] [CrossRef] [PubMed]
- Ermonval, M.; Baychelier, F.; Tordo, N. What Do We Know about How Hantaviruses Interact with Their Different Hosts? Viruses 2016, 8, 223. [Google Scholar] [CrossRef]
- Schonrich, G.; Rang, A.; Lutteke, N.; Raftery, M.J.; Charbonnel, N.; Ulrich, R.G. Hantavirus-induced immunity in rodent reservoirs and humans. Immunol. Rev. 2008, 225, 163–189. [Google Scholar] [CrossRef]
- Rockx, B.; Brining, D.; Kramer, J.; Callison, J.; Ebihara, H.; Mansfield, K.; Feldmann, H. Clinical outcome of henipavirus infection in hamsters is determined by the route and dose of infection. J. Virol. 2011, 85, 7658–7671. [Google Scholar] [CrossRef] [PubMed]
- Reperant, L.A.; van de Bildt, M.W.; van Amerongen, G.; Leijten, L.M.; Watson, S.; Palser, A.; Kellam, P.; Eissens, A.C.; Frijlink, H.W.; Osterhaus, A.D.; et al. Marked endotheliotropism of highly pathogenic avian influenza virus H5N1 following intestinal inoculation in cats. J. Virol. 2012, 86, 1158–1165. [Google Scholar] [CrossRef]
- Hue, K.D.; Tuan, T.V.; Thi, H.T.; Bich, C.T.; Anh, H.H.; Wills, B.A.; Simmons, C.P. Validation of an internally controlled one-step real-time multiplex RT-PCR assay for the detection and quantitation of dengue virus RNA in plasma. J. Virol. Methods 2011, 177, 168–173. [Google Scholar] [CrossRef] [Green Version]
- Kucinskaite-Kodze, I.; Petraityte-Burneikiene, R.; Zvirbliene, A.; Hjelle, B.; Medina, R.A.; Gedvilaite, A.; Razanskiene, A.; Schmidt-Chanasit, J.; Mertens, M.; Padula, P.; et al. Characterization of monoclonal antibodies against hantavirus nucleocapsid protein and their use for immunohistochemistry on rodent and human samples. Arch. Virol. 2011, 156, 443–456. [Google Scholar] [CrossRef]
- Haagmans, B.L.; Kuiken, T.; Martina, B.E.; Fouchier, R.A.; Rimmelzwaan, G.F.; van Amerongen, G.; van Riel, D.; de Jong, T.; Itamura, S.; Chan, K.H.; et al. Pegylated interferon-alpha protects type 1 pneumocytes against SARS coronavirus infection in macaques. Nat. Med. 2004, 10, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Van den Brand, J.M.; Stittelaar, K.J.; van Amerongen, G.; Rimmelzwaan, G.F.; Simon, J.; de Wit, E.; Munster, V.; Bestebroer, T.; Fouchier, R.A.; Kuiken, T.; et al. Severity of pneumonia due to new H1N1 influenza virus in ferrets is intermediate between that due to seasonal H1N1 virus and highly pathogenic avian influenza H5N1 virus. J. Infect. Dis. 2010, 201, 993–999. [Google Scholar] [CrossRef] [PubMed]
- McElhinney, L.M.; Marston, D.A.; Pounder, K.C.; Goharriz, H.; Wise, E.L.; Verner-Carlsson, J.; Jennings, D.; Johnson, N.; Civello, A.; Nunez, A.; et al. High prevalence of Seoul hantavirus in a breeding colony of pet rats. Epidemiol. Infect. 2017, 145, 3115–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.L.; Bird, B.H.; Glass, G.E. Sex differences in immune responses and viral shedding following Seoul virus infection in Norway rats. Am. J. Trop. Med. Hyg. 2001, 65, 57–63. [Google Scholar] [CrossRef] [PubMed]
- De Vries, A.; Vennema, H.; Bekker, D.L.; Maas, M.; Adema, J.; Opsteegh, M.; van der Giessen, J.W.; Reusken, C.B. Characterization of Puumala hantavirus in bank voles from two regions in the Netherlands where human cases occurred. J. Gen. Virol. 2016, 97, 1500–1510. [Google Scholar] [CrossRef] [PubMed]
- Maas, M.; de Vries, A.; van Roon, A.; Takumi, K.; van der Giessen, J.; Rockx, B. High Prevalence of Tula Hantavirus in Common Voles in The Netherlands. Vector Borne Zoonotic Dis. 2017, 17, 200–205. [Google Scholar] [CrossRef]
- Easterbrook, J.D.; Klein, S.L. Seoul virus enhances regulatory and reduces proinflammatory responses in male Norway rats. J. Med. Virol. 2008, 80, 1308–1318. [Google Scholar] [CrossRef] [Green Version]
- Wong, T.W.; Chan, Y.C.; Joo, Y.G.; Lee, H.W.; Lee, P.W.; Yanagihara, R. Hantavirus infections in humans and commensal rodents in Singapore. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 248–251. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ahn, C.; Han, J.S.; Kim, S.; Lee, J.S.; Lee, P.W. Hemorrhagic fever with renal syndrome caused by the Seoul virus. Nephron 1995, 71, 419–427. [Google Scholar] [CrossRef]
- Nielsen, C.F.; Sethi, V.; Petroll, A.E.; Kazmierczak, J.; Erickson, B.R.; Nichol, S.T.; Rollin, P.E.; Davis, J.P. Seoul virus infection in a Wisconsin patient with recent travel to China, March 2009: First documented case in the Midwestern United States. Am. J. Trop. Med. Hyg. 2010, 83, 1266–1268. [Google Scholar] [CrossRef]
- Gnemmi, V.; Verine, J.; Vrigneaud, L.; Glowacki, F.; Ratsimbazafy, A.; Copin, M.C.; Dewilde, A.; Buob, D. Microvascular inflammation and acute tubular necrosis are major histologic features of hantavirus nephropathy. Hum. Pathol. 2015, 46, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Strandin, T.; Makela, S.; Mustonen, J.; Vaheri, A. Neutrophil Activation in Acute Hemorrhagic Fever With Renal Syndrome Is Mediated by Hantavirus-Infected Microvascular Endothelial Cells. Front. Immunol. 2018, 9, 2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundstrom, J.B.; McMullan, L.K.; Spiropoulou, C.F.; Hooper, W.C.; Ansari, A.A.; Peters, C.J.; Rollin, P.E. Hantavirus infection induces the expression of RANTES and IP-10 without causing increased permeability in human lung microvascular endothelial cells. J. Virol. 2001, 75, 6070–6085. [Google Scholar] [CrossRef] [PubMed]
- Goeijenbier, M.; Meijers, J.C.; Anfasa, F.; Roose, J.M.; van de Weg, C.A.; Bakhtiari, K.; Henttonen, H.; Vaheri, A.; Osterhaus, A.D.; van Gorp, E.C.; et al. Effect of Puumala hantavirus infection on human umbilical vein endothelial cell hemostatic function: Platelet interactions, increased tissue factor expression and fibrinolysis regulator release. Front. Microbiol. 2015, 6, 220. [Google Scholar] [CrossRef] [PubMed]
- Gavrilovskaya, I.N.; Gorbunova, E.E.; Mackow, N.A.; Mackow, E.R. Hantaviruses direct endothelial cell permeability by sensitizing cells to the vascular permeability factor VEGF, while angiopoietin 1 and sphingosine 1-phosphate inhibit hantavirus-directed permeability. J. Virol. 2008, 82, 5797–5806. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| number tested | lung (pos) | kidney (pos) | liver (pos) | blood (pos) | saliva swab (pos) | rectal swab (pos) | urine (pos) | ||
|---|---|---|---|---|---|---|---|---|---|
| seropositive | adult | 19 | 19 | 15 | 14 | 14 | 12 | 8 | 3 |
| juvenile | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| seronegative | juvenile | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maas, M.; van Heteren, M.; de Vries, A.; Kuiken, T.; Hoornweg, T.; Veldhuis Kroeze, E.; Rockx, B. Seoul Virus Tropism and Pathology in Naturally Infected Feeder Rats. Viruses 2019, 11, 531. https://doi.org/10.3390/v11060531
Maas M, van Heteren M, de Vries A, Kuiken T, Hoornweg T, Veldhuis Kroeze E, Rockx B. Seoul Virus Tropism and Pathology in Naturally Infected Feeder Rats. Viruses. 2019; 11(6):531. https://doi.org/10.3390/v11060531
Chicago/Turabian StyleMaas, Miriam, Melanie van Heteren, Ankje de Vries, Thijs Kuiken, Tabitha Hoornweg, Edwin Veldhuis Kroeze, and Barry Rockx. 2019. "Seoul Virus Tropism and Pathology in Naturally Infected Feeder Rats" Viruses 11, no. 6: 531. https://doi.org/10.3390/v11060531
APA StyleMaas, M., van Heteren, M., de Vries, A., Kuiken, T., Hoornweg, T., Veldhuis Kroeze, E., & Rockx, B. (2019). Seoul Virus Tropism and Pathology in Naturally Infected Feeder Rats. Viruses, 11(6), 531. https://doi.org/10.3390/v11060531