Marine Morbilliviruses: Diversity and Interaction with Signaling Lymphocyte Activation Molecules


Abstract
:1. Introduction
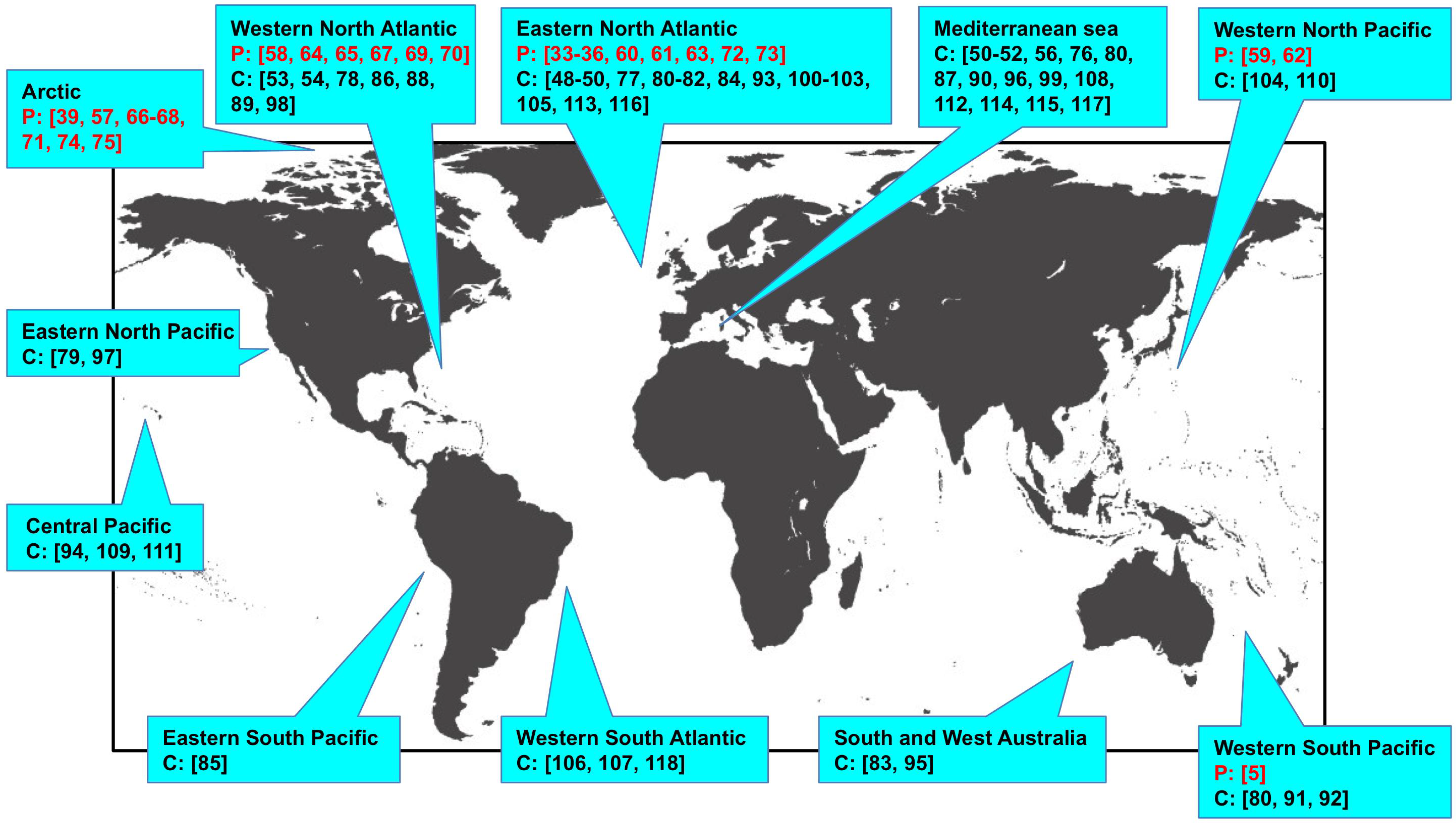
2. Discovery of Marine Morbilliviruses
3. Epidemiological Studies Based on Molecular Biology and Serology
4. Pathology and Pathogenesis
5. Transmission and Maintenance of PDV and CeMV
6. Interaction between SLAM and the Viral H Protein
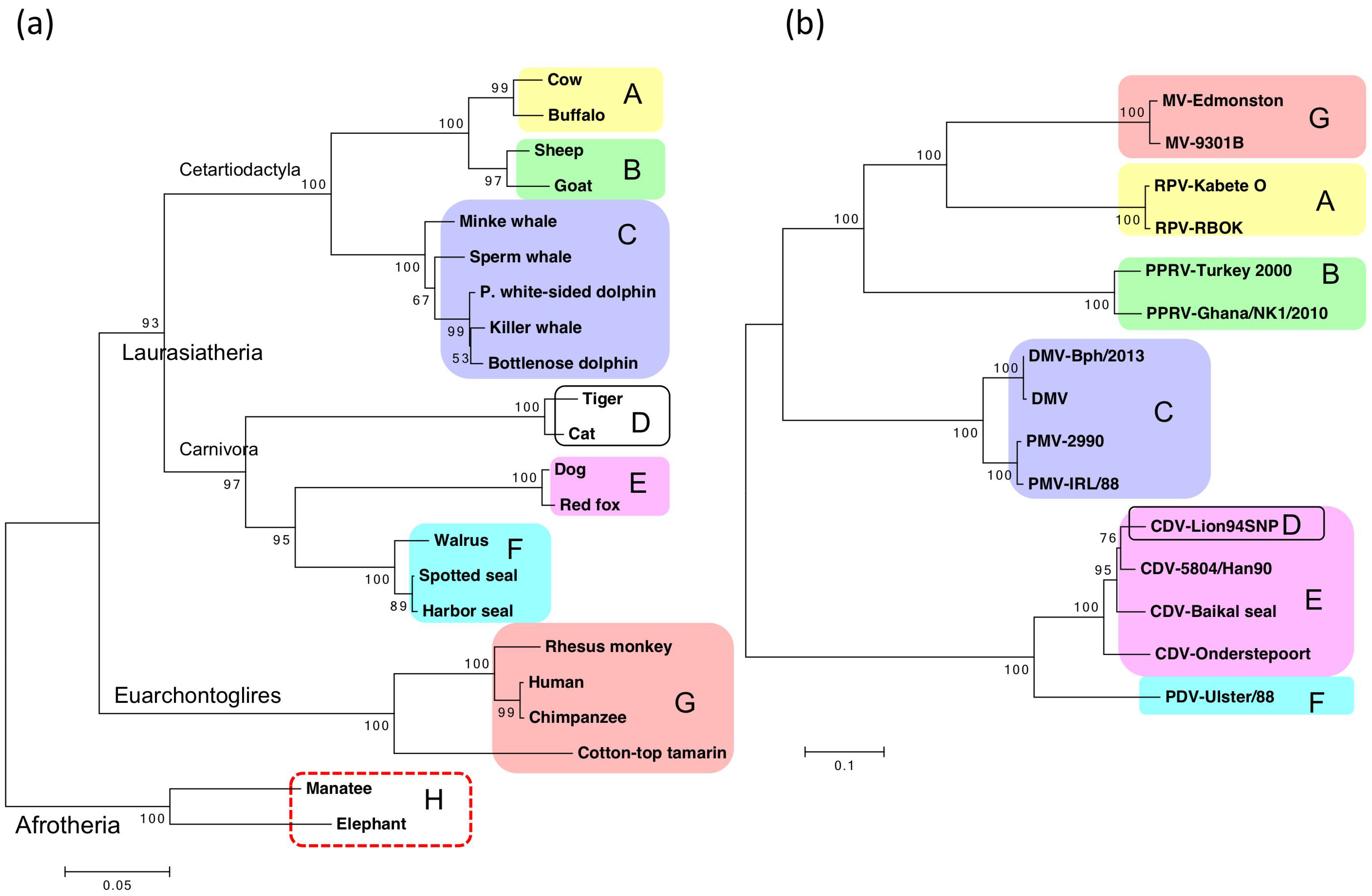
6.1. Phylogenetic Relationship between SLAMs and Viral H Proteins
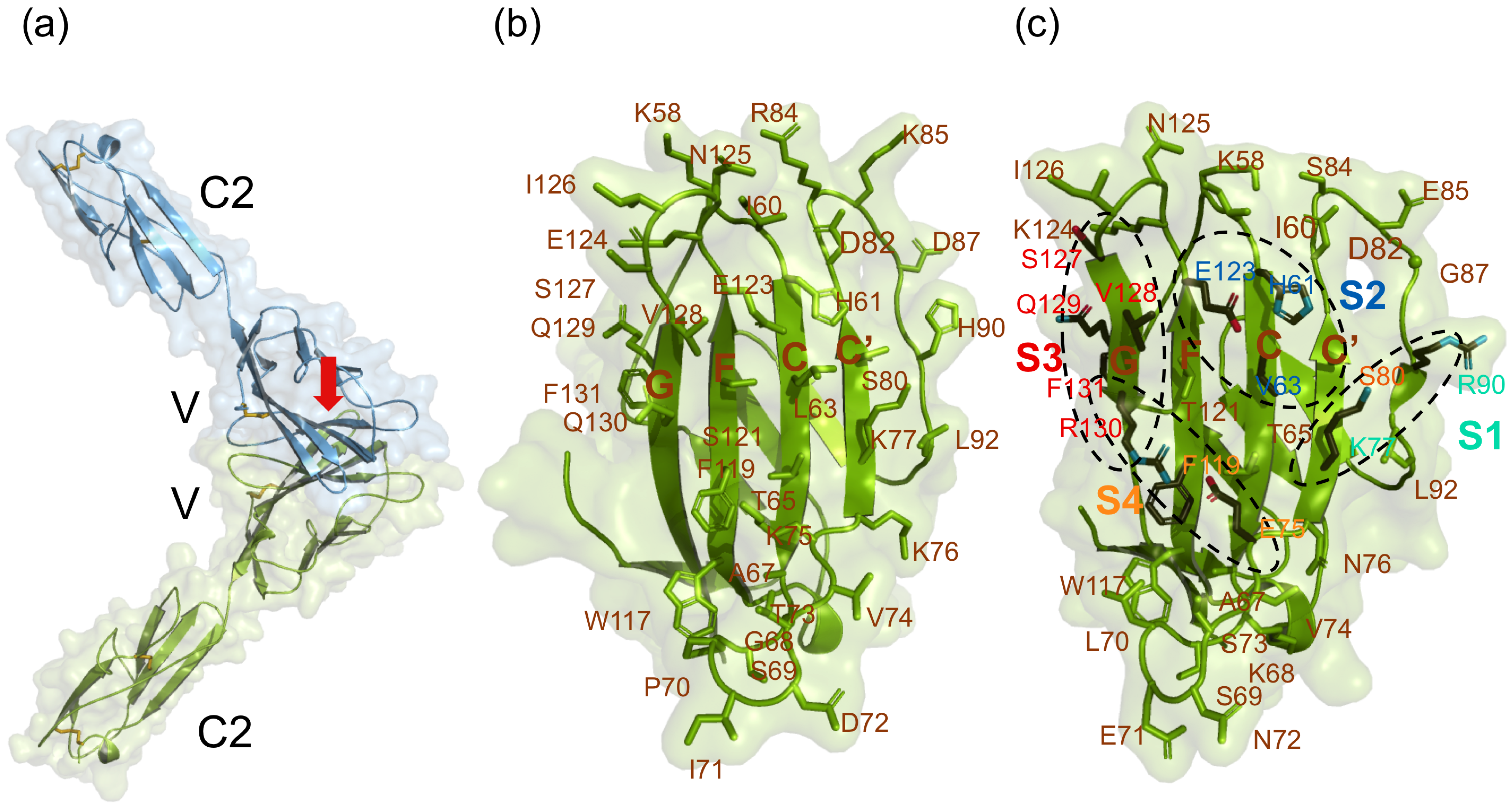
6.2. The SLAM–Viral H Protein Interface
6.3. SLAM Residues Important for Binding to the Morbillivirus H Protein
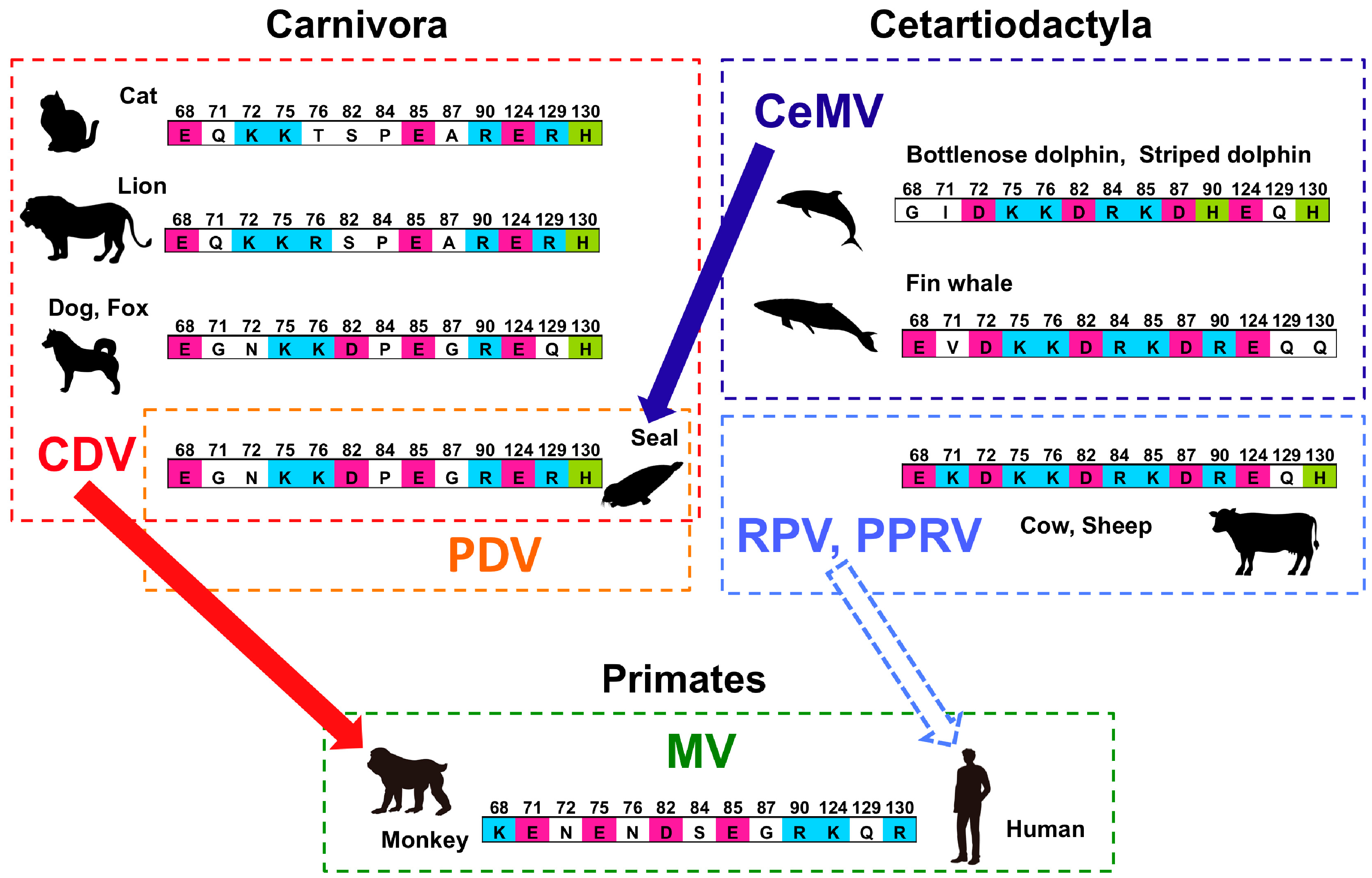
6.4. Sequence Variations in the 35 SLAM Residues Potentially Involved in H Protein Binding among Pinnipeds and Carnivores
6.5. Sequence Variations in the 35 SLAM Residues Potentially Involved in H Protein Binding Among Cetaceans
6.6. Host Switching and Expansion of CeMV Beyond the Order
7. In silico Analysis
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Griffin, D.E. Measles Virus. In Fields’ Virology, 3rd ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: New York, NY, USA, 2001; pp. 1401–1442. [Google Scholar]
- Barrett, T.; Blixenkrone-Møller, M.; di Guardo, G.; Domingo, M.; Duignan, P.; Hall, A.; Mamaev, L.; Osterhaus, A.D.M.E. Morbilliviruses in aquatic mammals: Report on round table discussion. Vet. Microbiol. 1995, 44, 261–265. [Google Scholar] [CrossRef]
- Kennedy, S. Morbillivirus infections in aquatic mammals. J. Comp. Pathol. 1998, 119, 201–225. [Google Scholar] [CrossRef]
- Di Guardo, G.; Marruchella, G.; Agrimi, U.; Kennedy, S. Morbillivirus infections in aquatic mammals: A brief overview. J. Vet. Med. Ser. A Physiol. Pathol. Clin. Med. 2005, 52, 88–93. [Google Scholar] [CrossRef]
- Duignan, P.J.; Van Bressem, M.-F.; Baker, J.; Barbieri, M.; Colegrove, K.M.; De Guise, S.; de Swart, R.; Di Guardo, G.; Dobson, A.; Duprex, W.P. Phocine Distemper Virus: Current knowledge and future directions. Viruses 2014, 6, 5093–5134. [Google Scholar] [CrossRef]
- Van Bressem, M.-F.; Duignan, P.J.; Banyard, A.; Barbieri, M.; Colegrove, K.M.; De Guise, S.; Di Guardo, G.; Dobson, A.; Domingo, M.; Fauquier, D. Cetacean Morbillivirus: Current Knowledge and Future Directions. Viruses 2014, 6, 5145–5181. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Wong, B.H.; Fan, R.Y.; Wong, A.Y.; Zhang, A.J.; Wu, Y.; Choi, G.K.; Li, K.S.; Hui, J.; et al. Feline morbillivirus, a previously undescribed paramyxovirus associated with tubulointerstitial nephritis in domestic cats. Proc. Natl. Acad. Sci. USA 2012, 109, 5435–5440. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, D.A.; Temmam, S.; Lebarbenchon, C.; Lagadec, E.; Chotte, J.; Guillebaud, J.; Ramasindrazana, B.; Heraud, J.M.; de Lamballerie, X.; Goodman, S.M.; et al. Identification of novel paramyxoviruses in insectivorous bats of Southwest Indian Ocean. Virus Res. 2012, 170, 159–163. [Google Scholar] [CrossRef]
- Oldstone, M.B.; Lewicki, H.; Thomas, D.; Tishon, A.; Dales, S.; Patterson, J.; Manchester, M.; Homann, D.; Naniche, D.; Holz, A. Measles virus infection in a transgenic model: Virus-induced immunosuppression and central nervous system disease. Cell 1999, 98, 629–640. [Google Scholar] [CrossRef]
- Schneider-Schaulies, S.; Schneider-Schaulies, J. Measles virus-induced immunosuppression. Curr. Top Microbiol. Immunol. 2009, 330, 243–269. [Google Scholar]
- Griffin, D.E. Measles virus-induced suppression of immune responses. Immunol. Rev. 2010, 236, 176–189. [Google Scholar] [CrossRef] [Green Version]
- Brindley, M.A.; Takeda, M.; Plattet, P.; Plemper, R.K. Triggering the measles membrane fusion machinery. Proc. Natl. Acad. Sci. USA 2012, 109, E3018–E3027. [Google Scholar] [CrossRef] [PubMed]
- Ader-Ebert, N.; Khosravi, M.; Herren, M.; Avila, M.; Alves, L.; Bringolf, F.; Orvell, C.; Langedijk, J.P.; Zurbriggen, A.; Plemper, R.K.; et al. Sequential conformational changes in the morbillivirus attachment protein initiate the membrane fusion process. PLoS Pathog. 2015, 11, e1004880. [Google Scholar] [CrossRef] [PubMed]
- Plattet, P.; Alves, L.; Herren, M.; Aguilar, H.C. M121easles virus fusion protein: structure, function and inhibition. Viruses 2016, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Herren, M.; Shrestha, N.; Wyss, M.; Zurbriggen, A.; Plattet, P. Regulatory role of the morbillivirus attachment protein head -to-stalk linker module in membrane fusion triggering. J. Virol. 2018, 92, e00679-18. [Google Scholar] [CrossRef] [PubMed]
- Tatsuo, H.; Ono, N.; Tanaka, K.; Yanagi, Y. SLAM (CDw150) is a cellular receptor for measles virus. Nature 2000, 406, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Muhlebach, M.D.; Mateo, M.; Sinn, P.L.; Prufer, S.; Uhlig, K.M.; Leonard, V.H.; Navaratnarajah, C.K.; Frenzke, M.; Wong, X.X.; Sawatsky, B.; et al. Adherens junction protein nectin-4 is the epithelial receptor for measles virus. Nature 2011, 480, 530–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noyce, R.S.; Bondre, D.G.; Ha, M.N.; Lin, L.T.; Sisson, G.; Tsao, M.S.; Richardson, C.D. Tumor cell marker PVRL4 (nectin 4) is an epithelial cell receptor for measles virus. PLoS Pathog. 2011, 7, e1002240. [Google Scholar] [CrossRef] [PubMed]
- De Witte, L.; de Vries, R.D.; van der Vlist, M.Y.; Yuksel, S.; Litjens, M.; de Swart, R.L.; Geijtenbeek, T.B. DC-SIGN and CD150 have distinct roles in transmission of measles virus from dendritic cells to T-lymphocytes. PLoS Pathog. 2008, 4, e1000049. [Google Scholar] [CrossRef]
- Leonard, V.H.; Sinn, P.L.; Hodge, G.; Miest, T.; Devaux, P.; Oezguen, N.; Braun, W.; McCray, P.B., Jr.; McChesney, M.B.; Cattaneo, R. Measles virus blind to its epithelial cell receptor remains virulent in rhesus monkeys but cannot cross the airway epithelium and is not shed. J. Clin. Investig. 2008, 118, 2448–2458. [Google Scholar] [CrossRef] [Green Version]
- De Vries, R.D.; Lemon, K.; Ludlow, M.; McQuaid, S.; Yuksel, S.; van Amerongen, G.; Rennick, L.J.; Rima, B.K.; Osterhaus, A.D.; de Swart, R.L.; et al. In vivo tropism of attenuated and pathogenic measles virus expressing green fluorescent protein in macaques. J. Virol. 2010, 84, 4714–4724. [Google Scholar] [CrossRef]
- Leonard, V.H.; Hodge, G.; Reyes-Del Valle, J.; McChesney, M.B.; Cattaneo, R. Measles virus selectively blind to signaling lymphocytic activation molecule (SLAM; CD150) is attenuated and induces strong adaptive immune responses in rhesus monkeys. J. Virol. 2010, 84, 3413–3420. [Google Scholar] [CrossRef] [PubMed]
- Ludlow, M.; Rennick, L.J.; Sarlang, S.; Skibinski, G.; McQuaid, S.; Moore, T.; de Swart, R.L.; Duprex, W.P. Wild-type measles virus infection of primary epithelial cells occurs via the basolateral surface without syncytium formation or release of infectious virus. J. Gen. Virol. 2010, 91, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Lemon, K.; de Vries, R.D.; Mesman, A.W.; McQuaid, S.; van Amerongen, G.; Yuksel, S.; Ludlow, M.; Rennickm, L.J.; Kuiken, T.; Rima, B.K.; et al. Early target cells of measles virus after aerozol of non-human primates. PLoS Pathog. 2011, 7, e1001263. [Google Scholar] [CrossRef] [PubMed]
- Sawatsky, B.; Wong, X.X.; Hinkelmann, S.; Cattaneo, R.; von Messling, V. Canine distemper virus epithelial cell infection is required for clinical disease but not for immunosuppression. J. Virol. 2012, 86, 3658–3666. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Tahara, M.; Nagata, N.; Seki, F. Wild-type measles virus is intrinsically dual-tropic. Front Microbiol. 2012, 2, 279. [Google Scholar] [CrossRef] [PubMed]
- Frenzke, M.; Sawatsky, B.; Wong, X.X.; Delpeut, S.; Mateo, M.; Cattaneo, R.; von Messling, V. Nectin-4-dependent measles virus spread to the cynomolgus monkey tracheal epithelium: role of infected immune cells infiltrating the lamina propria. J. Virol. 2013, 87, 2526–2534. [Google Scholar] [CrossRef] [PubMed]
- Sawatsky, B.; Cattaneo, R.; von Messling, V. Canine distemper virus spread and transmission to naive ferrets: selective pressure on signaling lymphocyte activation molecule-dependent entry. J. Virol. 2018, 92, e00669-18. [Google Scholar] [CrossRef] [PubMed]
- Tatsuo, H.; Ono, N.; Yanagi, Y. Morbilliviruses use signaling lymphocyte activation molecules (CD150) as cellular receptors. J. Virol. 2001, 75, 5842–5850. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, K.; Ando, A.; Suzuki, R.; Takishita, K.; Kawato, M.; Katsumata, E.; Ohtsu, D.; Okutsu, K.; Tokutake, K.; Miyahara, H.; et al. Host-virus specificity of morbilliviruses predicted by 6 structural modeling of the marine mammal SLAM, a receptor. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Noyce, R.S.; Delpeut, S.; Richardson, C.D. Dog nectin-4 is an epithelial cell receptor for canine distemper virus that facilitates virus entry and syncytia formation. Virology 2013, 436, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Delpeut, S.; Noyce, R.S.; Richardson, C.D. The tumor-associated marker, PVRL4 (nectin-4), is the epithelial receptor for morbilliviruses. Viruses 2014, 6, 2268–2286. [Google Scholar] [CrossRef] [PubMed]
- Osterhaus, A.D.; Vedder, E.J. Identification of virus causing recent seal deaths. Nature 1988, 335, 20. [Google Scholar] [CrossRef] [PubMed]
- Cosby, S.L.; Mcquaid, S.; Duffy, N.; Lyons, C.; Rima, B.K.; Allan, G.M.; Mccullough, S.J.; 10 Kennedy, S.; Smyth, J.A.; Mcneilly, F.; et al. Characterization of a seal morbillivirus. Nature 1988, 336, 115–116. [Google Scholar] [CrossRef]
- Kennedy, S.; Smyth, J.A.; McCullough, S.J.; Allan, G.M.; McNeilly, F.; McQuaid, S. Confirmation of cause of recent seal deaths. Nature 1988, 335, 404. [Google Scholar] [CrossRef] [PubMed]
- Mahy, B.W.J.; Barrett, T.; Evans, S.; Anderson, E.C.; Bostock, C.J. Characterization of a seal morbillivirus. Nature 1988, 336, 115. [Google Scholar] [CrossRef] [PubMed]
- Goodhart, C.B. Did virus transfer from harp seals to common seals. Nature 1988, 336, 21. [Google Scholar] [CrossRef]
- McGourty, C. Species jump may be responsible for seals’ virus infection. Nature 1988, 335, 3. [Google Scholar] [CrossRef] [PubMed]
- Dietz, R.; Ansen, C.T.; Have, P.; Heide-Jorgensen, M.P. Clue to seal epizootic. Nature 1989, 338, 627. [Google Scholar] [CrossRef]
- Osterhaus, A.D.M.E.; Groen, J.; Uytdehaag, F.G.C.M.; Visser, I.K.G.; Vedder, E.J.; Crowther, J.; Bostock, C.J. Morbillivirus infections in European seals before 1988. Vet. Rec. 1989, 125, 326. [Google Scholar] [CrossRef]
- Grachev, M.A.; Kumarev, V.P.; Mamaev, L.V.; Zorin, V.L.; Baranova, l.V.; Denikina, N.N.; Belikov, S.I.; Petrov, E.A.; Kolesnik, V.S.; Kolesnik, R.S.; et al. Distemper virus in Baikal seals. Nature 1989, 338, 209. [Google Scholar] [CrossRef]
- Osterhaus, A.D.M.E.; Groen, J.; Uytdehaag, F.G.C.M.; Visser, I.K.G.; Bildt, M.W.G.V.D.; Bergman, A.; Klingeborn, B. Distemper virus in Baikal seals. Nature 1989, 338, 209–210. [Google Scholar] [CrossRef]
- Visser, I.K.G.; Kumarev, V.P.; Orvell, C.; Vries, P.D.E.; Broeders, H.W.J.; van de Bildt, M.W.J.; Grosen, J.; Teppema, J.S.; Burger, M.C.; UytdeHaag, F.G.C.M.; et al. Comparison of two morbilliviruses isolated from seals during outbreaks of distemper in northwest Europe and Siberia. Arch. Virol. 1990, 111, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Crowther, J.; Osterhaus, A.D.M.E.; Subbarao, S.M.; Groen, J.; Haas, I.; Mamaev, I.V.; Titenko, A.M.; Visser, I.K.G.; Bostock, C.J. Molecular and serological studies on the recent seal virus epizootics in Europe and Siberia. Sci. Total Environ. 1992, 115, 117–132. [Google Scholar] [CrossRef] [Green Version]
- Mamaev, L.V.; Denikina, N.N.; Belikov, S.I.; Volchkov, V.E.; Visser, I.K.G.; Fleming, M.; Kai, C.; Harder, T.C.; Liess, B.; Osterhaus, A.D.M.E.; et al. Characterisation of morbillivirus isolated from Lake Baikal seals (Phoca sibirica). Vet. Microbiol. 1995, 44, 251–259. [Google Scholar] [CrossRef]
- Forsyth, M.A.; Kennedy, S.; Wilson, S.; Eybatov, T.; Barrett, T. Canine distemper virus in a Caspian seal. Vet. Rec. 1998, 143, 662–664. [Google Scholar] [PubMed]
- Kennedy, S.; Kuiken, T.; Jepson, P.D.; Deaville, R.; Forsyth, M.; Barrett, T.; van de Bildt, M.W.; Osterhaus, A.D.; Eybatov, T.; Duck, C.; et al. Mass die-Off of Caspian seals caused by canine distemper virus. Emerg. Infect. Dis. 2000, 6, 637–639. [Google Scholar] [CrossRef]
- Kennedy, S.; Smyth, J.A.; Cush, P.F.; McCullough, S.J.; Allan, G.M.; McQuaid, S. Viral distemper now found in porpoises. Nature 1988, 336, 21. [Google Scholar] [CrossRef]
- McCullough, S.J.; McNeilly, F.; Allan, G.M.; Kennedy, S.; Smyth, J.A.; Cosby, S.L.; McQuaid, S.; Rima, B.K. Isolation and characterisation of a porpoise morbillivirus. Arch. Virol. 1991, 118, 247–252. [Google Scholar] [CrossRef]
- Visser, I.K.; Van Bressem, M.F.; de Swart, R.L.; van de Bildt, M.W.; Vos, H.W.; van der Heijden, R.W.; Saliki, J.T.; Orvell, C.; Kiching, P.; Kuiken, T.; et al. Characterization of morbilliviruses isolated from dolphins and porpoises in Europe. J. Gen. Virol. 1993, 74, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Domingo, M.; Ferrer, L.; Pumarola, M.; Marco, A.; Plana, J.; Kennedy, S.; McAliskey, M.; Rima, B.K. Morbillivirus in dolphins. Nature 1990, 348, 21. [Google Scholar] [CrossRef]
- Van Bressem, M.-F.; Visser, I.K.G.; van de Bildt, M.W.; Teppema, J.S.; Raga, J.A.; Osterhaus, A.D.M.E. Morbillivirus infection in Mediterranean striped dolphins (Stenella coeruleoalba). Vet. Rec. 1991, 129, 471–472. [Google Scholar] [CrossRef] [PubMed]
- Lipscomb, T.P.; Schulman, F.Y.; Moffett, D.; Kennedy, S. Morbilliviral disease in Atlantic bottlenose dolphins (Tursiops truncatus) from the 1987–1988 epizootic. J. Wildl. Dis. 1994, 30, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Tsai, M.; Krafft, A.E.; Lichy, J.H.; Reid, A.H.; Schulman, F.Y.; Lipscomb, T.P. Two morbilliviruses implicated in bottlenose dolphin epizootics. Emerg. Infect. Dis. 1996, 2, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Curran, M.D.; O’Loan, D.; Rima, B.K.; Kennedy, S. Nucleotide sequence analysis of phocine distemper virus reveals its distinctness from canine distemper virus. Vet. Rec. 1990, 127, 430–431. [Google Scholar] [PubMed]
- Blixenkrone-Moller, M.; Bolt, G.; Gottschalk, E.; Kentner, M. Comparative analysis of the gene encoding the nucleocapsid protein of dolphin morbilliviruses. J. Gen. Virol. 1994, 75, 2829–2834. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.R.; Visser, I.K.G.; Broeders, H.W.; van de Bildt, M.W.; Bowen, W.D.; Osterhaus, A.D. Antibodies to phocine distemper virus Canadian seals. Vet. Rec. 1992, 130, 514–516. [Google Scholar] [CrossRef] [PubMed]
- Duignan, P.J.; Saliki, J.T.; St Aubin, D.J.; Early, G.; Sadove, S.; House, J.A.; Kovacs, K.; Geraci, J.R. Epizootiology of morbillivirus infection in North American harbor seals (Phoca vitulina) and gray seals (Halichoerus grypus). J. Wildl. Dis. 1995, 31, 491–501. [Google Scholar] [CrossRef]
- Ohashi, K.; Kai, C. Morbillivirus Infections in wildlife of Japan. J. Vet. Med. 2000, 53, 834–838. (In Japanese) [Google Scholar]
- Jensen, T.; Bildt, M.v.d.; Dietz, H.H.; Andersen, T.H.; Hammer, A.S.; Kuiken, T.; Osterhaus, A. Another phocine distemper outbreak in Europe. Science 2002, 297, 209. [Google Scholar] [CrossRef]
- Muller, G.; Wohlsein, P.; Beineke, A.; Haas, L.; Greiser-Wilke, I.; Siebert, U.; Fonfara, S.; 9 Harder, T.; Stede, M.; Gruber, A.D.; et al. Phocine distemper in German seals, 2002. Emerg. Infect. Dis. 2004, 10, 723–725. [Google Scholar] [CrossRef]
- Fujii, K.; Sato, H.; Kakumoto, C.; Kobayashi, M.; Saito, S.; Kariya, T.; Watanabe, Y.; Sakoda, Y.; Kai, C.; Kida, H.; et al. Seroepidemiological survey of morbillivirus infection in Kuril harbor seals (Phoca vitulina stejnegeri) of Hokkaido, Japan. Jpn. J. Vet. Res. 2006, 54, 109–117. [Google Scholar] [PubMed]
- Rijks, J.M.; Read, F.L.; van de Bildt, M.W.; van Bolhuis, H.G.; Martina, B.E.; Wagenaar, J.A.; 1 van der Meulen, K.; Osterhaus, A.D.; Kuiken, T. Quantitative analysis of the 2002 phocine distemper epidemic in the Netherlands. Vet. Pathol. 2008, 45, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Earle, J.A.P.; Melia, M.M.; Doherty, N.V.; Nielsen, O.; Cosby, S.L. Phocine distemper virus in seals, east coast, United States, 2006. Emerg. Infect. Dis. 2011, 17, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Siembieda, J.L.; Hall, A.J.; Gulland, F.M.D.; Rowles, T.; Garron, M.; Matassa, K.; Rotstein, D.S.; Gonzalez, S.; Northeast Region Marine Mammal Stranding Network; Johnson, C.K. Epidemiology of a phocine distemper virus outbreak along North Atlantic coast of the United States. Aqua. Mam. 2017, 43, 254–263. [Google Scholar] [CrossRef]
- Stuen, S.; Have, P.; Osterhaus, A.D.; Arnemo, J.M.; Moustgaard, A. Serological investigation of virus infections in harp seals (Phoca groenlandica) and hooded seals (Cystophora cristata). Vet. Rec. 1994, 134, 502–503. [Google Scholar] [CrossRef]
- Duignan, P.J.; Nielsen, O.; House, C.; Kovacs, K.M.; Duffy, N.; Early, G.; Sadove, S.; St Aubin, D.J.; Rima, B.K.; Geraci, J.R. Epizootiology of morbillivirus infection in harp, hooded, and ringed seals from the Canadian Arctic and western Atlantic. J. Wildl. Dis. 1997, 33, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Markussen, N.H.; Have, P. Phocine distemper virus infection in harp seals (Phoca groenlandica). Mar. Mamm. Sc. 1992, 8, 19–26. [Google Scholar] [CrossRef]
- Daoust, P.Y.; Haines, D.M.; Thorsen, J.; Duignan, P.J.; Geraci, J.R. Phocine distemper in a harp seal (Phoca groenlandica) from the Gulf of St. Lawrence, Canada. J. Wildl. Dis. 1993, 29, 114–117. [Google Scholar] [CrossRef]
- Lipscomb, T.P.; Mense, M.G.; Habecker, P.L.; Taubenberger, J.K.; Schoelkopf, R. Morbilliviral dermatitis in seals. Vet. Pathol. 2001, 38, 724–726. [Google Scholar] [CrossRef]
- Henderson, G.; Trudgett, A.; Lyons, C.; Ronald, K. Demonstration of antibodies in archival sera from Canadian seals reactive with a European isolate of phocine distemper virus. Sci. Total Environ. 1992, 115, 93–98. [Google Scholar] [CrossRef]
- Kennedy, S.A. Review of the 1988 European seal morbillivirus epizootic. Vet. Rec. 1990, 127, 563–567. [Google Scholar] [PubMed]
- Hughes, D.E.; Carter, S.D.; Robinson, I.; Clarke, D.D.; Clarke, C.J. Anti-canine distemper virus antibodies in common and grey seals. Vet. Rec. 1992, 130, 449–450. [Google Scholar] [CrossRef] [PubMed]
- Duignan, P.J.; Saliki, J.T.; St Aubin, D.J.; House, J.A.; Geraci, J.R. Neutralizing antibodies to phocine distemper virus in Atlantic walruses (Odobenus rosmarus rosmarus) from Arctic Canada. J. Wildl. Dis. 1994, 30, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, O.; Stewart, R.E.; Measures, L.; Duignan, P.; House, C. A morbillivirus antibody survey of Atlantic walrus, narwhal and beluga in Canada. J. Wildl. Dis. 2000, 36, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Van Bressem, M.-F.; Visser, I.K.G.; de Swart, R.L.; Örvell, C.; Stanzani, L.; Androukaki, E.; Siakavara, K.; Osterhaus, A.D.M.E. Dolphin morbillivirus in different parts of the Mediterranean Sea. Arch. Virol. 1993, 129, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Van Bressem, M.-F.; Jepson, P.; Barrett, T. Further insight on the epidemiology of cetacean morbillivirus in the northeastern Atlantic. Mar. Mamm. Sci. 1998, 14, 605–613. [Google Scholar] [CrossRef]
- Duignan, P.J.; House, C.; Geraci, J.R.; Duffy, N.; Rima, B.K.; Walsh, M.T.; Early, G.; St Aubin, D.J.; Sadove, S.; Koopman, H.; et al. Morbillivirus infection in cetaceans of the western Atlantic. Vet. Microbiol. 1995, 44, 241–249. [Google Scholar] [CrossRef]
- Reidarson, T.H.; McBain, J.; House, C.; King, D.P.; Stott, J.L.; Krafft, A.; Taubenberger, J.K.; Heyning, J.; Lipscomb, T.P. Morbillivirus infection in stranded common dolphins from the Pacific Ocean. J. Wildl. Dis. 1998, 34, 771–776. [Google Scholar] [CrossRef]
- Van Bressem, M.-F.; van Waerebeek, K.; Jepson, P.D.; Raga, J.A.; Duignan, P.J.; Nielsen, O.; di Beneditto, A.P.; Siciliano, S.; Ramos, R.; Kant, W.; et al. An insight into the epidemiology of dolphin morbillivirus worldwide. Vet. Microb. 2001, 81, 287–304. [Google Scholar] [CrossRef]
- Sierra, E.; Sánchez, S.; Saliki, J.T.; Blas-Machado, U.; Arbelo, M.; Zucca, D.; Fernández, A. Retrospective study of etiologic agents associated with nonsuppurative meningoencephalitis in stranded cetaceans in the Canary Islands. J. Clin. Microbiol. 2014, 52, 2390–2397. [Google Scholar] [CrossRef]
- Bento, M.; Eira, C.; Vingada, J.; Marçalo, A.; Ferreira, M.; Fernandez, A.; Tavares, L.; Duarte, A. New insight into dolphin morbillivirus phylogeny and epidemiology in the northeast Atlantic: opportunistic study in cetaceans stranded along the Portuguese and Galician coasts. BMC Vet. Res. 2016, 12, 176. [Google Scholar] [CrossRef] [PubMed]
- Kemper, C.; Tomo, I.; Bingham, J.; Bastianello, S.; Wang, J.; Gibbs, S.; Woolford, L.; Dickason, C.; Kelly, D. Morbillivirus-associated unusual mortality event in South Australian bottlenose dolphins is largest reported for the Southern Hemisphere. Royal Socie. open Sci. 2016, 3, 160838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Delgado, J.; Sierra, E.; Vela, A.I.; Arbelo1, M.; Zucca, D.; Groch, K.R.; Fernandez, A. Coinfection by Streptcoccus phocae and cetacean morbillivirus in a short-beaked common dolphin Delphinus delphis. Dis. Aquat. Organ. 2017, 11, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Van Bressem, M.-F.; van Waerebeek, K.; Fleming, M.; Barrett, T. Serological evidence of morbillivirus infection in small cetaceans from the Southeast Pacific. Vet. Microb. 1998, 59, 89–98. [Google Scholar] [CrossRef]
- Duignan, P.J.; House, C.; Ode11, D.K.; Wells, R.S.; Hansen, W.; Walsh, M.T.; Aubin, D.J., St.; Rima, B.K.; Geraci, J.R. Morbillivirus in bottlenose dolphins: Evidence for recurrent epizootics in the western Atlantic and Gulf of Mexico. Mar. Mamm. Sci. 1996, 12, 499–515. [Google Scholar] [CrossRef]
- Tsur, L.; Goffman, O.; Yakobsen, B.; Moffet, D.; Kennedy, S. Morbillivirus infection in a bottlenose dolphin (Tursiops truncatus) from the Mediterranean Sea. Eur. J. Vet. Pathol. 1997, 2, 83–85. [Google Scholar]
- Taubenberger, J.K.; Tsai, M.M.; Atkin, T.J.; Fanning, T.G.; Krafft, A.E.; Moeller, R.B.; Kodsi, S.E.; Mense, M.G.; Lipscomb, T.P. Molecular genetic evidence of a novel morbillivirus in a long-finned pilot whale (Globicephala melas). Emerg. Infect. Dis. 2000, 6, 42–45. [Google Scholar] [CrossRef]
- Bossart, G.D.; Reif, J.S.; Schaefer, A.M.; Goldstein, J.; Fair, P.A.; Saliki, J.T. Morbillivirus infection in free-ranging Atlantic bottlenose dolphins (Tursiops truncatus) from the Southeastern United States: Seroepidemiologic and pathologic evidence of subclinical infection. Vet. Microbiol. 2010, 143, 160–166. [Google Scholar] [CrossRef]
- Keck, N.; Kwiatek, O.; Dhermain, F.; Dupraz, F.; Boulet, H.; Danes, C.; Laprie, C.; Perrin, A.; Godenir, J.; Micout, L.; et al. Resurgence of Morbillivirus infection in Mediterranean dolphins off the French coast. Vet. Rec. 2010, 166, 654–655. [Google Scholar] [CrossRef] [Green Version]
- Stone, B.M.; Blyde, D.J.; Saliki, J.T.; Blas-Machado, U.; Bingham, J.; Hyatt, A.; Wang, J.; Payne, J.; Crameri, S. Fatal cetacean morbillivirus infection in an Australian offshore bottlenose dolphin (Tursiops truncatus). Aust. Vet. J. 2011, 89, 452–457. [Google Scholar] [CrossRef]
- Stone, B.M.; Blyde, D.J.; Saliki, J.T.; Morton, J.M. Morbillivirus infection in live stranded, injured, trapped, and captive cetaceans in southeastern Queensland and northern New South Wales, Australia. J. Wildl. Dis. 2012, 48, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Sierra, E.; Zucca, D.; Arbelo, M.; García-Álvarez, N.; Andrada, M.; Déniz, S.; Fernández, A. Fatal systemic morbillivirus infection in bottlenose dolphin, Canary Islands, Spain. Emerg. Infect. Dis. 2014, 20, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.M.; West, K.L.; Levine, G.; Sanchez, S.; Jensen, B.A. Initial characterization of novel beaked whale morbillivirus in Hawaiian cetaceans. Dis. Aqua. Org. 2016, 117, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Stephens, N.; Duignan, P.J.; Wang, J.; Bingham, J.; Finn, H.; Bejder, L.; Patterson, A.P.; Holyoake, C. Cetacean morbillivirus in coastal Indo-Pacific bottlenose dolphins, Western Australia. Emerg. Infect. Dis. 2014, 20, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Raga, J.A.; Banyard, A.; Domingo, M.; Corteyn, M.; Van Bressem, M.-F.; Fernández, M.; Aznar, F.J.; Barrett, T. Dolphin morbillivirus epizootic resurgence, Mediterranean Sea. Emerg. Infect. Dis. 2008, 14, 471–473. [Google Scholar] [CrossRef] [PubMed]
- Serrano, L.; Simeone, C.A.; Colegrove, K.M.; Duignan, P.J.; Goldstein, T.; Gulland, F.M.D. Cetacean morbillivirus in Odontocetes stranded along the central California coast, USA, 2000-2015. J. Wildl. Dis. 2017, 53, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Duignan, P.J.; House, C.; Geraci, J.R.; Early, G.; Copland, H.G.; Walsh, M.T.; Bossart, G.D.; Cray, C.; Sadove, S.; Aubin, D.J., St.; et al. Morbillivirus infection in two species of pilot whales (Globicephala sp.) from the western Atlantic. Mar. Mamm. Sci. 1995, 11, 150–162. [Google Scholar] [CrossRef]
- Fernández, A.; Esperón, F.; Herraéz, P.; Espinosa de los Monteros, A.; Clavel, C.; Bernabé, A.; Sanchez-Vizcaino, M.; Verborgh, P.; DeStephanis, R.; Toledano, F.; et al. Morbillivirus and pilot whale deaths, Mediterranean Sea. Emerg. Infect. Dis. 2008, 14, 792–794. [Google Scholar] [CrossRef]
- Sierra, E.; Fernández, A.; Suárez-Santana, C.; Xuriach, A.; Zucca, D.; de Quirós, Y.; García-Álvarez, N.; Fuente, J.; Sacchini, S.; Andrada, M.; et al. Morbillivirus and pilot whale deaths, Canary Islands, Spain, 2015. Emerg. Infect. Dis. 2016, 22, 740–742. [Google Scholar] [CrossRef]
- Bellière, E.N.; Esperón, F.; Fernández, A.; Arbelo, M.; Muñoz, M.J.; Sánchez-Vizcaíno, J.M. Phylogenetic analysis of a new Cetacean morbillivirus from a short-finned pilot whale stranded in the Canary Islands. Res. Vet. Sci. 2011, 90, 324–328. [Google Scholar] [CrossRef]
- Wohlsein, P.; Puff, C.; Kreutzer, M.; Siebert, U.; Baumgärtner, W. Distemper in a dolphin. Emerg. Infect. Dis. 2007, 13, 1959–1961. [Google Scholar] [CrossRef] [PubMed]
- Van Elk, C.E.; van de Bildt, M.W.; Jauniaux, T.; Hiemstra, S.; van Run, P.R.; Foster, G.; Meerbeek, J.; Osterhaus, A.D.M.E.; Kuiken, T. Is Dolphin Morbillivirus Virulent for White-Beaked Dolphins (Lagenorhynchus albirostris)? Vet. Pathol. 2014, 51, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Muranaka, M.; Horii, Y.; Murakami, N.; Yamaguchi, R.; Tateyama, S. Non-purulent meningoencephalomyelitis of a Pacific striped dolphin (Lagenorhynchus obliquidens). The first evidence of morbillivirus infection in a dolphin at the Pacific Ocean around Japan. J. Vet. Med. Sci. 1999, 61, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Sierra, E.; Fernandez, A.; Zucca, D.; Camara1, N.; Felipe-Jimenez, I.; Suarez-Santana, C.; de Quiros, Y.B.; Diaz-Delgado, J.; Arbelo, M. Morbillivirus infection in Risso’s dolphin Grampus griseus: a phylogenetic and pathological study of cases from the Canary Islands. study of cases from the Canary Islands. Dis. Aquat. Organ. 2018, 129, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Groch, K.R.; Colosio, A.C.; Marcondes, M.C.; Zucca, D.; Díaz-Delgado, J.; Niemeyer, C.; Marigo, J.; Brandão, P.E.; Fernández, A.; Catão-Dias, J.L. Novel cetacean morbillivirus in Guiana dolphin, Brazil. Emerg. Infect. Dis. 2014, 20, 511–513. [Google Scholar] [CrossRef] [PubMed]
- Groch, K.R.; Santos-Neto, E.B.; Díaz-Delgado, J.; Ikeda, J.M.P.; Carvalho, R.R.; Oliveira, R.B.; Guari, E.B.; Bisi, T.L.; Azevedo, A.F.; Lailson-Brito, J.; et al. Guiana dolphin unusual mortality event and link to cetacean morbillivirus, Brazil. Emerg. Infect. Dis. 2018, 24, 1349–1354. [Google Scholar] [CrossRef]
- Centelleghe, C.; Giorgia Beffagna, G.; Palmisano, G.; Franzo, G.; Casalone, C.; Pautasso, A.; Giorda, F.; Di Nocera, F.; Iaccarino, D.; Santoro, M.; et al. Dolphin Morbillivirus in a Cuvier’s Beaked Whale (Ziphius cavirostris), Italy. Front. Microbiol. 2017, 8, 111. [Google Scholar] [CrossRef]
- West, K.L.; Sanchez, S.; Rotstein, D.; Robertson, K.M.; Dennison, S.; Levine, G.; Davis, N.; Schofield, D.; Potter, C.W.; Jensen, B. A Longman’s beaked whale (Indopacetus pacificus) strands in Maui, Hawaii, with first case of morbillivirus in the central Pacific. Mar. Mamm. Sci. 2013, 29, 767–776. [Google Scholar] [CrossRef]
- Yang, W.C.; Pang, V.F.; Jeng, C.R.; Chou, L.S.; Chueh, L.L. Morbilliviral infection in a pygmy sperm whale (Kogia breviceps) from Taiwanese waters. Vet. Microbiol. 2006, 116, 69–76. [Google Scholar] [CrossRef]
- West, K.L.; Levine, G.; Jacob, J.; Jensen, B.; Sanchez, S.; Colegrove, K.; Rotstein, D. Coinfection and vertical transmission of Brucella and morbillivirus in a neonatal sperm whale (Physeter macrocephalus) in Hawaii, USA. J. Wildl. Dis. 2015, 51, 227–232. [Google Scholar] [CrossRef]
- Mazzariol, S.; Centelleghe, C.; Di Provvido, A.; Di Renzo, L.; Cardeti, G.; Cersini, A.; Fichi, G.; Petrella, A.; Di Francesco, C.E.; Mignone, W.; et al. Dolphin morbillivirus associated with a mass stranding of sperm whales, Italy. Emerg. Infect. Dis. 2017, 23, 144–146. [Google Scholar] [CrossRef]
- Jauniaux, T.; Charlier, G.; Desmecht, M.; Haelters, J.; Jacques, T.; Losson, B.; van Gompel, J.; Tavernier, J.; Coignoul, F. Pathological findings in two fin whales (Balaenoptera physalus) with evidence of morbillivirus infection. J. Comp. Pathol. 2000, 123, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Mazzariol, S.; Marcer, F.; Mignone, W.; Serracca, L.; Goria, M.; Marsili, L.; di Guardo, G.; Casalone, C. Dolphin Morbillivirus and Toxoplasma gondii coinfection in a Mediterranean fin whale (Balaenoptera physalus). BMC Vet. Res. 2012, 8, e20. [Google Scholar] [CrossRef]
- Mazzariol, S.; Centelleghe, C.; Beffagna, G.; Povinelli, M.; Terracciano, G.; Cocumelli, C.; Pintore, A.; Denurra, D.; Casalone, C.; Pautasso, A.; et al. Mediterranean fin whales (Balaenoptera physalus) threatened by dolphin morbillivirus. Emerg. Infect. Dis. 2016, 22, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.K.; Grilo, M.L.; Wohlsein, P.; Andersen-Ranberg, E.U.; Hansen, M.S.; Carl, C.; Kinze, C.C.; Hjulsager, C.K.; Olsen, M.T.; Lehnert, K.; et al. Dolphin Morbillivirus in a Fin Whale (Balaenoptera physalus) in Denmark, 2016. J. Wildl. Dis. 2017, 53, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Di Guardo, G.; Agrimi, U.; Morelli, L.; Cardeti, G.; Terracciano, G.; Kennedy, S. Post mortem investigations on cetaceans found stranded on the coasts of Italy between 1990 and 1993. Vet. Rec. 1995, 136, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Groch, K.R.; Kolesnikovas, C.K.M.; de Castilho, P.V.; Moreira, L.M.P.; Barros, C.R.M.B.; Morais, C.R.; Renault-Braga, E.P.; Sierra, E.; Fernandez, A.; Catao-Dias, J.L.; et al. Cetacean morbillivirus in Southern right whales, Brazil. Transbound. Emerg. Dis. 2019, 66, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.K.; Kruppa, J.; Habierski, A.; van de Bildt, M.; Mazzariol, S.; Di Guardo, G.; Siebert, U.; Kuiken, T.; Jung, K.; Osterhaus, A.; et al. Evolutionary evidence for multi-host transmission of cetacean morbillivirus. Emerg. Microb. Infect. 2018, 7, 201. [Google Scholar] [CrossRef]
- Harder, T.; Willhaus, T.; Frey, H.R.; Liess, B. Morbillivirus infections of seals during the 1988 epidemic in the Bay of Heligoland: III. Transmission studies of culture-propagated phocine distemper virus in harbor seals (Phoca vitulina) and a grey seal (Halichoerus grypus): clinical, virological and serological results. Zentralbl. Veterinarmed B. 1990, 37, 641–650. [Google Scholar]
- Ohno, S.; Ono, N.; Seki, F.; Takeda, M.; Kura, S.; Tsuzuki, T.; Yanagi, Y. Measles virus infection of SLAM (CD150) knockin mice reproduces tropism and immunosuppression in human infection. J Virol. 2007, 81, 1650–1659. [Google Scholar] [CrossRef]
- Von Messling, V.; Svitek, N.; Cattaneo, R. Receptor (SLAM [CD150]) recognition and the V protein sustain swift lymphocyte-based invasion of mucosal tissue and lymphatic organs by a morbillivirus. J. Virol. 2006, 80, 6084–6092. [Google Scholar] [CrossRef] [PubMed]
- Melia, M.M.; Earle, J.P.; Abdullah, H.; Reaney, K.; Tangy, F.; Cosby, S.L. Use of SLAM and PVRL4 and identification of pro-HB-EGF as cell entry receptors for wild type phocine distemper virus. PLoS ONE 2014, 9, e106281. [Google Scholar] [CrossRef] [PubMed]
- Di Guardo, G.; Mazzariol, S. Cetacean morbillivirus-associated pathology: knowns and unknowns. Front. Microbiol. 2016, 7, 112. [Google Scholar] [CrossRef] [PubMed]
- Domingo, M.; Vilafranca, M.; Visa, J.; Prats, N.; Trudgett, A.; Visser, I. Evidence for chronic morbillivirus infection in the Mediterranean striped dolphin (Stenella coeruleoalba). Vet. Microbiol. 1995, 44, 229–239. [Google Scholar] [CrossRef]
- Di Guardo, G.; Counmelli, C.; Scholl, F.; Difrancesco, C.E.; Spernanza, R.; Pennelli, M.; Eleni, C. Morbilliviral encephalitis in a striped dolphin Stenella coeruleoalba calf from Italy. Dis. Aquat. Organ. 2011, 95, 247–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, S.; Alba, A.; Ganges, L.; Vidal, E.; Raga, J.A.; Alegre, F.; González, B.; Medina, P.; Zorrilla, I.; Martínez, J. Post-epizootic chronic dolphin morbillivirus infection in Mediterranean striped dolphins Stenella coeruleoalba. Dis. Aquat. Organ. 2011, 96, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.K. Subacute sclerosing panencephalitis. J. Neurol. 2008, 255, 1861–1871. [Google Scholar] [CrossRef]
- Oldstone, M.B. Modeling subacute sclerosing panencephalitis in a transgenic mouse system: uncoding pathogenesis of disease and illuminating components of immune control. Curr. Top Microbiol. Immunol. 2009, 330, 31–54. [Google Scholar]
- Wendorf, K.A.; Winter, K.; Zipprich, J.; Schechter, R.; Hacker, J.K.; Preas, C.; Cherry, J.D.; Glaser, C.; Harriman, K. Subacute sclerosing panencephalitis: the complication that might be more common than previously estimated. Clin. Infect. Dis. 2017, 65, 226–232. [Google Scholar] [CrossRef]
- Domingo, M. Immunocytochemical studies of morbillivirus infection in striped dolphins. In Molecular and cell biology of marine mammals; Pfeiffer, C.J., Ed.; Krieger Publishing Company: Malabar, FL, USA, 2002; pp. 173–177. [Google Scholar]
- Kweder, H.; Ainouze, M.; Cosby, S.L.; Muller, C.P.; Levy, C.; Verhoeyen, E.; Cosset, F.L.; Manet, E.; Buckland, R. Mutations in the H, F, or M proteins can facilitate resistance of measles virus to neutralizing human anti-MV Sera. Adv. Virol. 2014. [CrossRef]
- Kweder, H.; Ainouze, M.; Brunel, J.; Gerlier, D.; Manet, E.; Buckland, R. Measles Virus: Identification in the M protein primary sequence of a potential molecular marker for subacute sclerosing panencephalitis. Adv. Virol. 2015. [CrossRef] [PubMed]
- Von Messling, V.; Milosevic, D.; Cattaneo, R. Tropism illuminated: lymphocyte-based pathways blazed by lethal morbillivirus through the host immune system. Proc. Natl. Acad. Sci. USA 2004, 101, 14216–14221. [Google Scholar] [CrossRef] [PubMed]
- De Swart, R.L.; Ludlow, M.; de Witte, L.; Yanagi, Y.; van Amerongen, G.; McQuaid, S.; Yuksel, S.; Geijtenbeek, T.B.; Duprex, W.P.; Osterhaus, A.D. Predominant infection of CD150+ lymphocytes and dendritic cells during measles virus infection of macaques. PLoS Pathog. 2007, 3, e178. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Tahara, M.; Hashiguchi, T.; Sato, T.A.; Jinnouchi, F.; Ueki, S.; Ohno, S.; Yanagi, Y. A human lung carcinoma cell line supports efficient measles virus growth and syncytium formation via a SLAM- and CD46-independent mechanism. J. Virol. 2007, 81, 12091–12096. [Google Scholar] [CrossRef] [PubMed]
- Tahara, M.; Takeda, M.; Shirogane, Y.; Hashiguchi, T.; Ohno, S.; Yanagi, Y. Measles virus infects both polarized epithelial and immune cells by using distinctive receptor-binding sites on its hemagglutinin. J. Virol. 2008, 82, 4630–4637. [Google Scholar] [CrossRef] [PubMed]
- Shirogane, Y.; Takeda, M.; Tahara, M.; Ikegame, S.; Nakamura, T.; Yanagi, Y. Epithelial-mesenchymal transition abolishes the susceptibility of polarized epithelial cell lines to measles virus. J. Biol. Chem. 2010, 285, 20882–20890. [Google Scholar] [CrossRef]
- Black, F. Epidemiology of Paramyxoviridae. In The Paramyxoviruses; Kingsburry, D.W., Ed.; Plenum Press: New York, NY, USA, 1991; pp. 509–536. [Google Scholar]
- Murphy, W.J.; Eizirik, E.; O’Brien, S.J.; Madsen, O.; Scally, M.; Douady, C.J.; et al. Resolution of the early placental mammal radiation using Bayesian phylogenetics. Science 2001, 294, 2348–2351. [Google Scholar] [CrossRef]
- Kriegs, J.O.; Churakov, G.; Kiefmann, M.; Jordan, U.; Brosius, J.; Schmitz, J. Retroposed elements as archives for the evolutionary history of placental mammals. PLoS Biol. 2006, 4, 537–544. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Veillette, A.; Cruz-Munoz, M.; Zhong, M. SLAM family receptors and SAP-related adaptors: matters arising. Trends Immunol. 2006, 27, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Mavaddat, N.; Mason, D.W.; Aikinson, P.D.; Evans, E.J.; Gilbert, R.J.; Stuart, D.I.; Fennelly, J.A.; Barclay, A.N.; Davis, S.J.; Brown, M.H. Signaling lymphocytic activation molecule (CDw150) is homophilic but self-associates with very low affinity. J Biol Chem 2000, 275, 28100–28109. [Google Scholar] [CrossRef] [PubMed]
- Ono, N.; Tatsuo, H.; Tanaka, K.; Minagawa, H.; Yanagi, Y. V domain of human SLAM (CDw150) is essential for its function as a measles virus receptor. J. Virol. 2001, 75, 1594–1600. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, T.; Kajikawa, M.; Maita, N.; Takeda, M.; Kuroki, K.; Sasaki, K.; Kohda, D.; Yanagi, Y.; Maenaka, K. Crystal structure of measles virus hemagglutinin provides insight into effective vaccines. Proc. Natl. Acad. Sci. USA 2007, 104, 19535–19540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashiguchi, T.; Ose, T.; Kubota, M.; Maita, N.; Kamishikiryo, J.; Maenaka, K.; Yanagi, Y. Structure of the measles virus hemagglutinin bound to its cellular receptor SLAM. Nat. Struct. Mol. Biol. 2011, 18, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Kobune, F.; Sakata, H.; Sugiura, A. Marmoset lymphoblastoid cells as a sensitive host for isolation of measles virus. J. Virol. 1990, 64, 700–705. [Google Scholar] [Green Version]
- Shimizu, Y.; Ohishi, K.; Suzuki, R.; Tajima, Y.; Yamada, T.; Kakizoe, Y.; Bando, T.; Fujise, Y.; Taru, H.; Murayama, T.; et al. Amino acid sequence variations of signaling lymphocyte activation molecule and mortality caused by morbillivirus infection in cetaceans. Microbiol. Immunol. 2013, 57, 624–632. [Google Scholar] [PubMed]
- Ohishi, K.; Suzuki, R.; Maruyama, T. Host-Virus Specificity of the Morbillivirus Receptor, SLAM, in Marine Mammals: Risk Assessment of Infection Based on Three-Dimensional Models. In New Approaches to the Study of Marine Mammals; Romero, A., Keith, E.O., Eds.; InTech: Janeza Trdine, Rijeka, Croatia, 2014; pp. 123–204. [Google Scholar]
- Bieringer, M.; Han, J.W.; Kendl, S.; Khosravi, M.; Plattet, P.; Schneider-Schaulies, J. Experimental adaptation of wild-type canine distemper virus (CDV) to the human entry receptor CD150. PLoS ONE 2013, 8, e57488. [Google Scholar] [CrossRef]
- Khosravi, M.; Bringolf, F.; Rothlisberger, S.; Bieringer, M.; Schneider-Schaulies, J.; Zubriggen, A.; Origgi, F.; Plattet, P. Canine distemper virus fusion activation: critical role of residue E123 of CD150/SLAM. J. Virol. 2016, 90, 1622–1637. [Google Scholar] [CrossRef]
- Blixenkrone-Moller, M.V.; Svansson, V.; Have, P.; Botner, A.; Nielsen, J. Infection studies in mink with seal-derived morbillivirus. Arch. Virol. 1989, 106, 165–170. [Google Scholar] [CrossRef]
- Goldstein, T.; Mazet, J.A.; Gill, V.A.; Doroff, A.M.; Burek, K.A.; Hammond, J.A. Phocine distemper virus in northern sea otters in the Pacific ocean, Alaska, USA. Emerg. Infect. Dis. 2009, 15, 925–927. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Rima, B.K. Molecular Biology of Morbillivirus Diseases of Marine Mammals. In Molecular and Cell Biology of Marine Mammals; Pfeiffer, C.J., Ed.; Krieger Publishing Company: Malabar, FL, USA, 2002; pp. 161–172. [Google Scholar]
- Beineke, A.; Baumgärtner, W.; Wohlsein, P. Cross-species transmission of canine distemper virus-an update. One Health 2015, 1, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gutierrez, M.; Ruiz-Saenz, J. Diversity of susceptible hosts in canine distemper hosts in canine distemper virus infection: a systematic review and data synthesis. BMC Vet. Res. 2016, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, M.H.; Yasue, K.; Ohshima, H.; Abe, H.; Kato, T.; Kishiro, T.; Goto, M.; Munechika, I.; Okada, N. Molecular evidence from retroposons that whales form a clade within even-toothed ungulates. Nature 1997, 388, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, M.; Matsuno, F.; Hamilton, H.; Brownell, R.L., Jr.; Cao, Y.; Ding, W.; Zuoyan, Z.; Shedlock, A.M.; Fordyce, R.E.; Hasegawa, M.; et al. Retroposon analysis of major cetacean lineage: The monophyly of toothed whales and the paraphyly of river dolphins. Proc Natl Acad Sci USA 2001, 98, 7384–7389. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, M.; Hamilton, H.; Makino, H.; Sasaki, T.; Takahshi, K.; Goto, M.; Kanda, N.; Pastene, L.A.; Okada, N. Baleen whale phylogeny and a past extensive radiation event revealed by SINE insertion analysis. Mol Biol Evol 2005, 23, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, T.; Kohno, N.; Hasegawa, M. The monophyletic origin of sea lions and fur seals (Carnivora; Otariidae) in the South hemisphere. Gene 2009, 441, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Appel, M.J.G.; Yates, R.A.; Foly, G.L.; Bernstein, J.J.; Santinelli, S.; Spelman, L.H.; Miller, L.D.; Arp, L.H.; Anderson, M.; Barr, M. Canine distemper epizootic in lions, tigers, and leopards in North America. J. Vet. Diag. Invest. 1994, 6, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Roelke-Parker, M.E.; Munson, L.; Packer, C.; Kock, R.; Cleaveland, S.M.; Carpenter, M.; O’Brian, S.J. A canine distemper virus epidemic in Serengeti lions (Panthera leo). Nature 1996, 79, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, K.; Suzuki, R.; Maeda, T.; Tsuda, M.; Abe, E.; Yoshida, T.; Endo, Y.; Okamura, M.; Nagamine, T.; Yamamoto, H.; et al. Recent host range expansion of canine distemper virus and variation in its receptor, signaling lymphocyte activation molecule, in carnivores. J. Wildl. Dis. 2014, 50, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y.; Suzuki, J.; Noguchi, K.; Terada, Y.; Shimoda, H.; Mizuno, T.; Maeda, K. Function of feline signaling lymphocyte activation molecule as a receptor of canine distemper virus. J. Vet. Med. Sci. 2013, 75, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Nikolin, V.M.; Osterrieder, K.; von Messling, V.; Hofer, H.; Anderson, D.; Dubovi, E.; Brunner, E.; East, M. Antagonistic pleiotropy and fitness trade-offs reveal specialist and generalist traits in strains of canine distemper virus. PLOS ONE 2012, 7, e50955. [Google Scholar] [CrossRef] [PubMed]
- Nikolin, V.M.; Olarte-Castillo, X.A.; Osterrieder, N.; Hofer, H.; Dubovi, E.; Mazzoni, C.J.; Brunner, E.; Goller, K.V.; Fyumagwa, R.D.; Moehlman, P.D.; et al. Canine distemper virus in the Serengeti ecosystem: molecular adaptation of different carnivore species. Mol. Ecol. 2017, 7, 2111–2130. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Visser, I.K.; Mamaev, L.; Goatley, L.; van Bressem, M.F.; Osterhaust, A.D. Dolphin and porpoise morbilliviruses are genetically distinct from phocine distemper virus. Virology 1993, 193, 1010–1012. [Google Scholar] [CrossRef] [PubMed]
- Mazzariol, S.; Peletto, S.; Mondin, A.; Centelleghe, C.; di Guardo, G.; di Francesco, C.E.; Casalone, C.; Acutis, P.L. Dolphin morbillivirus infection in a captive harbor seal (Phoca vitulina). J. Clin. Microbiol. 2013, 51, 708–711. [Google Scholar] [CrossRef]
- Osterhaus, A.; Groen, J.; Niesters, H.; van de Bildt, M.; Martina, B.; Vedder, L.; Vos, J.; van Egmond, H. Abou-Sidi, B.; Barham, M.E. Morbillivirus in monk seal mass mortality. Nature 1997, 388, 838–839. [Google Scholar] [CrossRef] [PubMed]
- Van de Bildt, M.W.; Martina, B.E.; Sidi, B.A.; Osterhaus, A.D.M.E. Morbillivirus infection in a bottlenose dolphin and a Mediterranean monk seal from the Atlantic coast of West Africa. Vet. Rec. 2001, 148, 210–211. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, O.; Smith, G.; Weingartl, H.; Lair, S.; Measures, L. Use of a SLAM transfected Vero cell line to isolate and characterize marine mammal morbilliviruses using an experimental ferret model. J. Wildl. Dis. 2008, 44, 600–611. [Google Scholar] [CrossRef]
- Peletto, S.; Caruso, C.; Cerutti, F.; Modesto, P.; Biolatti, C.; Pautasso, A.; Grattarola, C.; Giorda, F.; Mazzariol, S.; Mignone, W.; et al. Efficient isolation on Vero.dogSLAMtag cells and full genome characterization of dolphin morbillivirus (DMV) by next generation sequencing. Sci. Rep. 2018, 8, 860. [Google Scholar] [CrossRef]
- Iwasaki, M.; Yanagi, Y. Expression of the Sendai (murine parainfluenza) virus C protein alleviates restriction of measles virus growth in mouse cells. Proc. Natl. Acad. Sci. USA 2011, 108, 15384–15389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stejskalova, K.; Bayerova1, Z.; Futas, J.; Hrazdilova, K.; Klumplerova, M.; Oppelt, J.; Splichalova, P.; Di Guardo, G.; Mazzariol, S.; Di Francesco, C.E. Candidate gene molecular markers as tools for analyzing genetic susceptibility to morbillivirus infection in stranded cetaceans. HLA 2017, 90, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, Y.; Ochikubo, F.; Matsubara, Y.; Tsuruoka, H.; Ishii, M.; Shirota, K.; Nomura, Y.; Sugiyama, M.; Yamanouchi, K. Natural infection with canine distemper virus in a Japanese monkey (Macaca fuscata). Vet. Microbiol. 1989, 20, 193–205. [Google Scholar] [CrossRef]
- Sun, Z.; Li, A.; Ye, H.; Shi, Y.; Hu, Z.; Zeng, L. Natural infection with canine distemper virus in hand-feeding Rhesus monkeys in China. Vet. Microbiol. 2010, 141, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Nagata, N.; Ami, Y.; Seki, F.; Suzaki, Y.; Iwata-Yoshikawa, N.; Suzuki, T.; Fukushi, S.; Mizutani, T.; Yoshikawa, T.; et al. Lethal canine distemper virus outbreak in cynomolgus monkeys in Japan in 2008. J. Virol. 2013, 87, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Yoshikawa, T.; Seki, F.; Fukushi, S.; Tahara, M.; Nagata, N.; Ami, Y.; Mizutani, T.; Kurane, I.; Yamaguchi, R.; et al. Canine distemper virus associated with a lethal outbreak in monkeys can readily adapt to use human receptors. J. Virol. 2013, 87, 7170–7175. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.; Kelly, J.T.; Graham, S.C.; Birch, J.; Gonalves-Carneiro, D.; Mitchell, T.; Thompson, R.N.; Lythgoe, K.A.; Logan, N.; Hosie, M.J.; et al. Structure-guided identification of a nonhuman morbillivirus with zoonotic potential. J. Virol. 2018, 92, e01248-18. [Google Scholar] [CrossRef] [PubMed]
- Santiago, C.; Celma, M.L.; Stehle, T.; Casasnovas, J.M. Structure of the measles virus hemagglutinin bound to the CD46 receptor. Nat. Struct. Mol. Biol. 2010, 17, 124–129. [Google Scholar] [CrossRef]
- Zhang, X.; Lu, G.; Qi, J.; Li, Y.; He, Y.; Xu, X.; Shi, J.; Zhang, C.W.; Yan, J.; Gao, G.F. Structure of measles virus hemagglutinin bound to its epithelial receptor nectin-4. Nat. Struct. Mol. Biol. 2013, 20, 67–72. [Google Scholar] [CrossRef]
- Xu, F.; Tanaka, S.; Watanabe, H.; Shiman, Y.; Iwasawa, M.; Ohishi, K.; Maruyama, T. Computational analysis of the interaction energies between amino acid residues of the measles virus hemagglutinin and its receptors. Viruses 2018, 10, 236. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | References |
|---|---|---|
| Phocidae | Harbor seal (Phoca vitulina) | [33] *, [34,35,36,57,58,59], [60,61] *, [62], [63,64,65] * |
| Ringed seal (Phoca hispida) | [39,66,67] | |
| Spotted seal (Phoca larga) | [59] | |
| Harp seal (Phoca groenlandicus) | [39], [65] *, [66,67,68,69,70] | |
| Grey seal (Halichoerus grypus) | [57,58,65,71,72,73] | |
| Hooded seal (Cystophora cristata) | [66,67,70] | |
| Odobenidae | Walrus (Odobenus rosmarus) | [74,75] |
| Otariidae | Steller sea lion (Eumetopias jubatus) | [59] |
| Hooker’s sea lion (Phocartos hookeri) | [5] | |
| New Zealand fur seal (Arctocephalus forsteri) | [5] |
| Family | Species | References |
|---|---|---|
| Odontoceti | ||
| Delphinidae | Common dolphin (Delphinus delphis) | [76,77,78], [79] *, [80], [81,82,83] *, [84] |
| Long-beaked common dolphin (Delphinus capensis) | [85] | |
| Bottlenose dolphin (Tursiops truncatus) | [53], [54] *, [80], [83] *, [85,86,87], [88] *, [89], [90,91,92,93,94] * | |
| Indo-Ocean bottlenose dolphin (Tursiops aduncus) | [83,92,95] * | |
| Striped dolphin (Stenella coeruleoalba) | [51,52] *, [76,78,80], [81,82,90,94,96,97] * | |
| Atlantic spotted dolphin (Stenella frontalis) | [78] | |
| Long-finned pilot whale (Globicephala melas) | [80], [88,90] *, [98], [99,100] * | |
| Short-finned pilot whale (Globicephala macrorhynchus) | [81,98], [100,101] * | |
| White-beaked dolphin (Lagenorhynchus albirostris | [50,77], [102,103] * | |
| Atlantic white-sided dolphin (Lagenorhynchus acutus) | [78] | |
| Pacific white-sided dolphin (Lagenorhynchus obliquidens) | [104] | |
| Dusky dolphin (Lagenorhynchus obscurus) | [85] | |
| Rough-toothed dolphin (Steno bredanensis) | [94] * | |
| Spotted dolphin (Stenella attenuata) | [94] * | |
| Spinner dolphin (Stenella longirostris) | [94] * | |
| Fraser’s dolphin (Lagenodelphis hosei) | [78,80,92] | |
| Risso’s dolphin (Grampus griseus) | [80], [94,105] * | |
| False killer whale (Pseudorca crassidens) | [78] | |
| Melon-headed whale (Peponocephala electra) | [92] | |
| Pygmy killer whale (Feresa attenuata) | [78] | |
| Guiana dolphin (Sotalia guianensis) | [106,107] * | |
| Phocoenidae | Harbor porpoise (Phocoena phocoena) | [48,49,50] *, [77,78,80] |
| Ziphiidae | Cuvier’s beaked whale (Ziphius cavirostris) | [94,108] * |
| Longman’s beaked whale (Indopacetus pacificus) | [94,109] * | |
| Blainville’s beaked whale (Mesoplodon densirostris) | [94] * | |
| Kogiidae | Pygmy sperm whale (Kogia breviceps) | [78], [94,110] * |
| Physeteridae | Sperm whale (Physeter macrocephalus) | [94,111,112] * |
| Mysticeti | ||
| Balaenopteridae | Fin whale (Balaenoptera physalus) | [56] *, [113], [114,115,116] * |
| Common minke whale (Balaenoptera acutorostrata) | [117] | |
| Bryde’s whale (Balaenoptera edeni) | [92] | |
| Humpback whale (Megaptera novaeangliae) | [94] * | |
| Balaenidae | Southern right whale (Eubalaena australis) | [118] * |
| (a) | ||||||||||||||||||||
| Residue 1 | 58 | 60 | 61 | 63 | 65 | 67 | 68 | 69 | 70 | 71 | 72 | 73 | 74 | 75 | 76 | 77 | 80 | 82 | 84 | Virus |
| β-strand 2 | C | C | C | C | C’ | C’ | C’ | |||||||||||||
| Site 3 | 2 | 2 | 4 | 1 | ||||||||||||||||
| Seal | K | I | H | L | T | A | E | S | P | G | N | S | V | K | K | K | S | D | P | PDV |
| Dog | K | I | H | L | T | A | E | S | P | G | N | S | I | K | K | K | S | D | P | CDV |
| Dolphin | K | I | H | L | T | A | G | S | P | I | D | T | V | K | K | K | S | D | R | CeMV |
| Cow | K | I | H | L | T | A | E | S | P | K | D | T | V | K | K | K | S | D | R | RPV |
| Sheep | K | I | H | L | T | A | E | S | P | R | D | T | V | K | K | K | S | D | R | PPRV |
| Humans | K | I | H | V | T | A | K | S | L | E | N | S | V | E | N | K | S | D | S | MV |
| (b) | ||||||||||||||||||||
| Residue 1 | 85 | 87 | 90 | 92 | 117 | 119 | 121 | 123 | 124 | 125 | 126 | 127 | 128 | 129 | 130 | 131 | Virus | |||
| β-strand 2 | F | F | F | F | G | G | G | G | G | |||||||||||
| Site 3 | 1 | 4 | 2 | 3 | 3 | 3 | 3&4 | 3 | ||||||||||||
| Seal | E | G | R | L | W | F | T | E | E | N | F | S | V | R | H | F | PDV | |||
| Dog | E | G | R | L | W | F | T | E | E | N | F | S | V | Q | H | F | CDV | |||
| Dolphin | K | D | H | L | W | F | S | E | E | N | I | S | V | Q | H | F | CeMV | |||
| Cow | K | D | R | L | W | F | S | E | E | N | V | S | V | Q | H | F | RPV | |||
| Sheep | K | G | H | L | W | F | S | E | E | N | V | S | V | Q | H | F | PPRV | |||
| Humans | E | G | R | L | W | L | T | E | K | N | V | S | V | Q | R | F | MV | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohishi, K.; Maruyama, T.; Seki, F.; Takeda, M. Marine Morbilliviruses: Diversity and Interaction with Signaling Lymphocyte Activation Molecules. Viruses 2019, 11, 606. https://doi.org/10.3390/v11070606
Ohishi K, Maruyama T, Seki F, Takeda M. Marine Morbilliviruses: Diversity and Interaction with Signaling Lymphocyte Activation Molecules. Viruses. 2019; 11(7):606. https://doi.org/10.3390/v11070606
Chicago/Turabian StyleOhishi, Kazue, Tadashi Maruyama, Fumio Seki, and Makoto Takeda. 2019. "Marine Morbilliviruses: Diversity and Interaction with Signaling Lymphocyte Activation Molecules" Viruses 11, no. 7: 606. https://doi.org/10.3390/v11070606
APA StyleOhishi, K., Maruyama, T., Seki, F., & Takeda, M. (2019). Marine Morbilliviruses: Diversity and Interaction with Signaling Lymphocyte Activation Molecules. Viruses, 11(7), 606. https://doi.org/10.3390/v11070606