Evaluation of Real-Time RT-PCR for Diagnostic Use in Detection of Puumala Virus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Samples
2.2. Construction of Synthetic Standard PUUV RNA
2.3. Primers and Probes
2.4. PUUV-qRT-PCR
2.5. RT-Nested-PCR
2.6. Detection of Immunoglobulins
3. Results
3.1. PUUV-qRT-PCR Is Both Sensitive and Specific
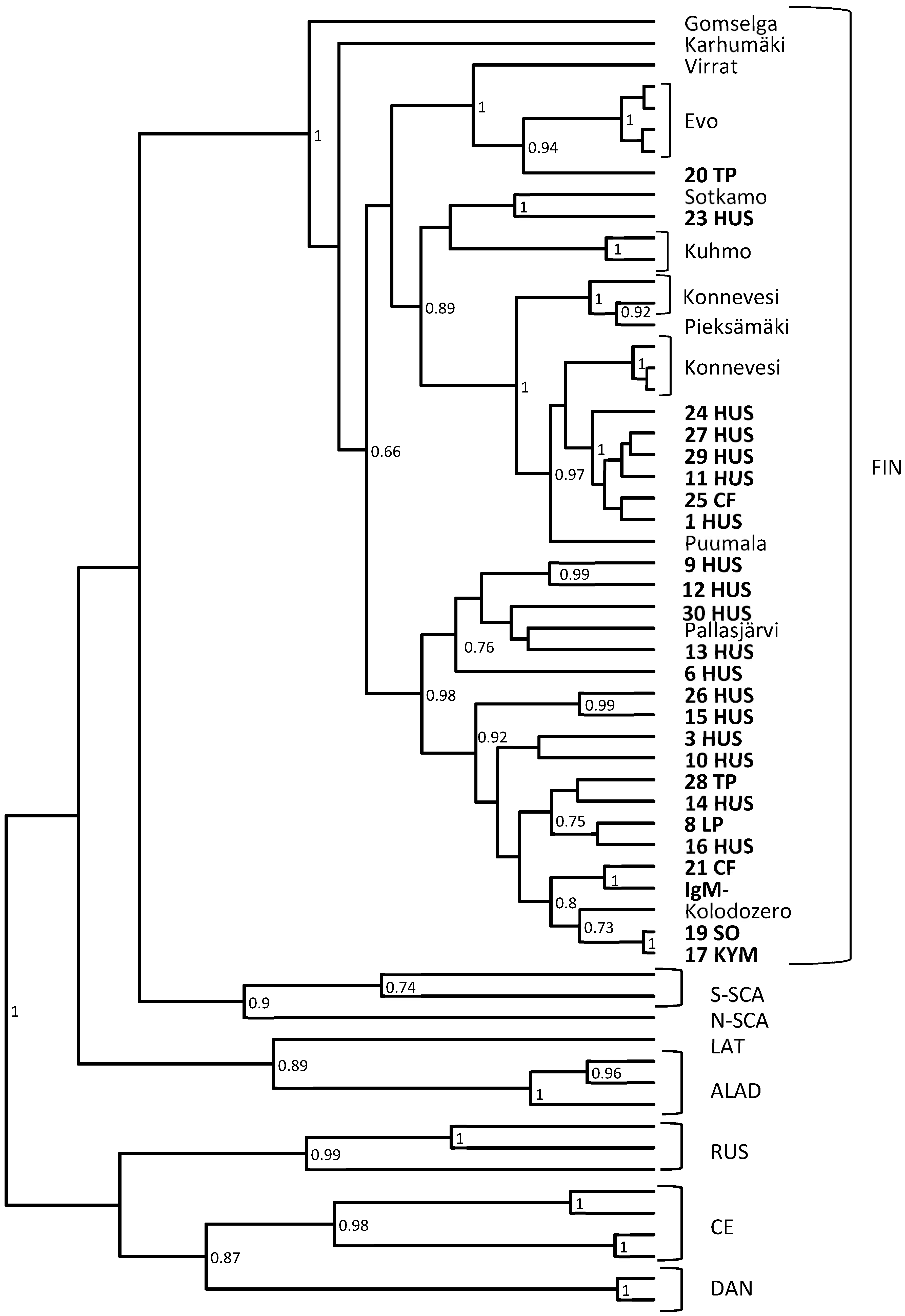
3.2. RT-Nested-PCR and Subsequent Sequencing Shows the High Variability of PUUV Strains Detected
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krüger, D.; Figueiredo, L.T.M.; Song, J.W.; Klempa, B. Hantaviruses—Globally emerging pathogens. J. Clin. Virol. 2015, 64, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Makary, P.; Kanerva, M.; Ollgren, J.; Virtanen, M.J.; Vapalahti, O.; Lyytikäinen, O. Disease burden of Puumala virus infections, 1995–2008. Epidemiol. Infect. 2010, 138, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Sironen, T.; Voutilainen, L.; Hepojoki, S.; Niemimaa, J.; Isoviita, V.M. Hantaviruses in Finnish soricomorphs: Evidence for two distinct hantaviruses carried by Sorex araneus suggesting ancient host–switch. Infect. Genet. Evol. 2014, 27, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Vaheri, A.; Hepojoki, S.; Levanov, L.; Jääskeläinen, A.; Henttonen, H. Serological survey of Seewis virus antibodies in patients suspected for hantavirus infection in Finland; a cross-reaction between Puumala virus antiserum with Seewis virus N protein. J. Gen. Virol. 2015, 96, 1664–1675. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Arai, S.; Hope, A.G.; Song, J.W.; Cook, J.A.; Yanagihara, R. Genetic diversity and phylogeography of Seewis virus in the Eurasian common shrew in Finland and Hungary. Virol. J. 2009, 24, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Lähdevirta, J. Nephropathia epidemica in Finland. A clinical histological and epidemiological study. Ann. Clin. Res. 1971, 3, 1–54. [Google Scholar] [PubMed]
- Mustonen, J.; Brummer-Korvenkontio, M.; Hedman, K.; Pasternack, A.; Pietila, K.; Vaheri, A. Nephropathia epidemica in Finland: A retrospective study of 126 cases. Scand. J. Infect. Dis. 1994, 26, 7–13. [Google Scholar] [CrossRef]
- Vapalahti, O.; Mustonen, J.; Lundkvist, A.; Henttonen, H.; Plyusnin, A.; Vaheri, A. Hantavirus infections in Europe. Lancet. Infect. Dis. 2003, 3, 653–661. [Google Scholar] [CrossRef]
- Kallio-Kokko, H.; Leveelahti, R.; Brummer-Korvenkontio, M.; Lundkvist, A.; Vaheri, A.; Vapalahti, O. Human immune response to Puumala virus glycoproteins and nucleocapsid protein expressed in mammalian cells. J. Med. Virol. 2001, 65, 605–613. [Google Scholar] [CrossRef]
- Mäkelä, S.; Ala-Houhala, I.; Mustonen, J.; Koivisto, A.M.; Kouri, T.; Turjanmaa, V. Renal function and blood pressure five years after Puumala virus-induced nephropathy. Kidney Int. 2000, 58, 1711–1718. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, M.H.; Makela, S.M.; Ala-Houhala, I.O.; Huhtala, H.S.; Koobi, T.; Vaheri, A.I. Ten-year prognosis of Puumala hantavirus-induced acute interstitial nephritis. Kidney Int. 2006, 69, 2043–2048. [Google Scholar] [CrossRef] [Green Version]
- Mäkelä, S.; Jaatinen, P.; Miettinen, M.; Salmi, J.; Ala-Houhala, I.; Huhtala, H.; Hurme, M.; Pörsti, I.; Vaheri, A.; Mustonen, J. Hormonal deficiencies during and after Puumala hantavirus infection. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Rasmuson, J.; Andersson, C.; Norrman, E.; Haney, M.; Evander, M.; Ahlm, C. Time to revise the paradigm of hantavirus syndromes? Hantavirus pulmonary syndrome caused by European hantavirus. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 685–690. [Google Scholar] [CrossRef] [Green Version]
- Sironen, T.; Sane, J.; Lokki, M.L.; Meri, S.; Andersson, L.C.; Hautala, T.; Kauma, H.; Vuorinen, S.; Rasmuson, J.; Evander, M.; et al. Fatal Puumala hantavirus disease: Involvement of complement activation and vascular leakage in the pathobiology. Open Forum Infect. Dis. 2017, 4, 229. [Google Scholar] [CrossRef]
- Pettersson, L.; Thunberg, T.; Rocklov, J.; Klingström, J.; Evander, M.; Ahlm, C. Viral load and humoral immune response in association with disease severity in Puumala hantavirus-infected patients-implications for treatment. Clin. Microbiol. Infect. 2014, 20, 235–241. [Google Scholar] [CrossRef]
- Kallio-Kokko, H.; Vapalahti, O.; Lundkvist, Å.; Vaheri, A. Evaluation of Puumala virus IgG and IgM enzyme immunoassays based on recombinant baculovirus-expressed nucleocapsid protein for early nephropathia epidemica diagnosis. Clin. Diagn. Virol. 1998, 10, 83–90. [Google Scholar] [CrossRef]
- Lundkvist, Å.; Hukic, M.; Horling, J.; Gilljam, M.; Nichol, S.; Niklasson, B. Puumala and Dobrava viruses cause hemorrhagic fever with renal syndrome in Bosnia-Herzegovina: Evidence of highly cross-neutralizing antibody responses in early patient sera. J. Med. Virol. 1997, 53, 51–59. [Google Scholar] [CrossRef]
- Plyusnin, A.; Hörling, J.; Kanerva, M.; Mustonen, J.; Cheng, Y.; Partanen, J. Puumala hantavirus genome in patients with nephropathia epidemica: Correlation of PCR positivity with HLA haplotype and link to viral sequences in local rodents. J. Clin. Microbiol. 1997, 35, 1090–1096. [Google Scholar]
- Vapalahti, O.; Lundkvist, A.; Kallio-Kokko, H.; Paukku, K.; Julkunen, I.; Lankinen, H. Antigenic properties and diagnostic potential of puumala virus nucleocapsid protein expressed in insect cells. J. Clin. Microbiol. 1996, 34, 119–125. [Google Scholar] [Green Version]
- Evander, M.; Eriksson, I.; Pettersson, L.; Juto, P.; Ahlm, C.; Olsson, G.E. Puumala hantavirus viremia diagnosed by real-time reverse transcriptase PCR using samples from patients with hemorrhagic fever and renal syndrome. J. Clin. Microbiol. 2007, 45, 2491–2497. [Google Scholar] [CrossRef]
- Lagerqvist, N.; Hagstrom, A.; Lundahl, M.; Nilsson, E.; Juremalm, M.; Larsson, I. Molecular diagnosis of Puumala virus-caused hemorrhagic fever with renal syndrome. J. Clin. Microbiol. 2016, 54, 1335–1339. [Google Scholar] [CrossRef]
- Voutilainen, L.; Sironen, T.; Tonteri, E.; Back, A.T.; Razzauti, M.; Karlsson, M. Life-long shedding of Puumala hantavirus in wild bank voles (Myodes glareolus). J. Gen. Virol. 2015, 96, 1238–1247. [Google Scholar] [CrossRef]
- Saksida, A.; Duh, D.; Korva, M.; Avsic-Zupanc, T. Dobrava virus RNA load in patients who have hemorrhagic fever with renal syndrome. J. Infect. Dis. 2008, 197, 681–685. [Google Scholar] [CrossRef]
- Korva, M.; Saksida, A.; Kejzar, N.; Schmaljohn, C.; Avsic-Zupanc, T. Viral load and immune response dynamics in patients with haemorrhagic fever with renal syndrome. Clin. Microbiol. Infect. 2013, 19, 358–366. [Google Scholar] [CrossRef]
- Bunz, H.; Weyrich, P.; Peter, A.; Baumann, D.; Tschritter, O.; Guthoff, M.; Beck., R.; Jahn., G.; Artunc., F.; Häring, H.U.; et al. Urinary neutrophil gelatinase-associated lipocalin (NGAL) and proteinuria predict severity of acute kidney injury in Puumala virus infection. BMC Infect. Dis. 2015, 15, 464. [Google Scholar] [CrossRef]


{kind=link}
| Patient No | Sex | Age | Hospital District | IgM | PUUV-qRT-PCR | RT-Nested-PCR |
|---|---|---|---|---|---|---|
| 1 | F | 39 | HUS | pos | pos | pos |
| 2 | M | 25 | HUS | pos | pos | NEG |
| 3 | F | 38 | HUS | pos | pos | pos |
| 4 | M | 55 | HUS | pos | NEG | NEG |
| 5 | F | 45 | HUS | pos | NEG | NEG |
| 6 | M | 65 | HUS | pos | pos | pos |
| 7 | F | 30 | HUS | pos | pos | NEG |
| 8 | F | 61 | LP | pos | pos | pos |
| 9 | F | 26 | HUS | pos | pos | pos |
| 10 | M | 57 | HUS | pos | pos | pos |
| 11 | F | 47 | HUS | pos | pos | pos |
| 12 | F | 64 | HUS | pos | pos | pos |
| 13 | M | 29 | KYM | pos | pos | pos |
| 14 | F | 61 | HUS | pos | pos | pos |
| 15 | F | 32 | HUS | pos | pos | pos |
| 16 | M | 46 | HUS | pos | pos | pos |
| 17 | F | 65 | KYM | pos | pos | pos |
| 18 | M | 66 | HUS | pos | pos | NEG |
| 19 | M | 71 | SO | pos | pos | pos |
| 20 | M | 77 | TP | pos | pos | pos |
| 21 | M | 48 | CF | pos | pos | pos |
| 22 | F | 28 | HUS | pos | pos | pos |
| 23 | M | 50 | HUS | pos | pos | pos |
| 24 | M | 39 | HUS | pos | pos | pos |
| 25 | M | 51 | CF | pos | pos | pos |
| 26 | F | 66 | HUS | pos | pos | pos |
| 27 | F | 51 | HUS | pos | pos | pos |
| 28 | M | 43 | TP | pos | pos | pos |
| 29 | M | 54 | HUS | pos | pos | pos |
| 30 | M | 57 | HUS | pos | pos | pos |
| No of positives: | 30 | 28 | 25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niskanen, S.; Jääskeläinen, A.; Vapalahti, O.; Sironen, T. Evaluation of Real-Time RT-PCR for Diagnostic Use in Detection of Puumala Virus. Viruses 2019, 11, 661. https://doi.org/10.3390/v11070661
Niskanen S, Jääskeläinen A, Vapalahti O, Sironen T. Evaluation of Real-Time RT-PCR for Diagnostic Use in Detection of Puumala Virus. Viruses. 2019; 11(7):661. https://doi.org/10.3390/v11070661
Chicago/Turabian StyleNiskanen, Silja, Anne Jääskeläinen, Olli Vapalahti, and Tarja Sironen. 2019. "Evaluation of Real-Time RT-PCR for Diagnostic Use in Detection of Puumala Virus" Viruses 11, no. 7: 661. https://doi.org/10.3390/v11070661
APA StyleNiskanen, S., Jääskeläinen, A., Vapalahti, O., & Sironen, T. (2019). Evaluation of Real-Time RT-PCR for Diagnostic Use in Detection of Puumala Virus. Viruses, 11(7), 661. https://doi.org/10.3390/v11070661