Evaluation of Diagnostic Performance of Three Indirect Enzyme-Linked Immunosorbent Assays for the Detection of IgG Antibodies to Ebola Virus in Human Sera

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens and Ethics Clearance
2.2. ELISA Serum Controls and Internal Quality Control (IQC)
2.3. Enzyme-Linked Immunosorbent Assays
2.3.1. Production of Ebola Virus Whole Antigen
2.3.2. Prodution of Recombinant Ebola Virus Nucleoprotein
2.3.3. Recombinant Ebola Virus Glycoprotein
2.3.4. Indirect Enzyme-Linked Immunosorbent Assay Procedures
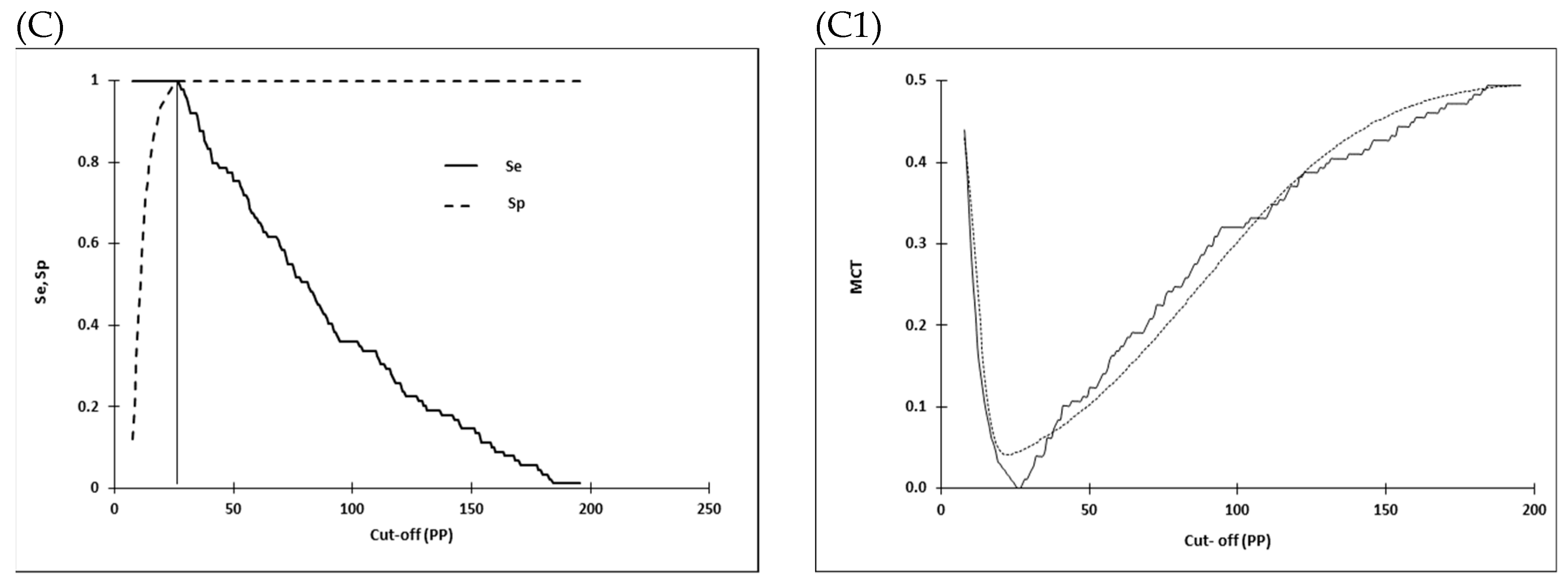
2.4. Selection of Cut-off Values and Statistical Analysis
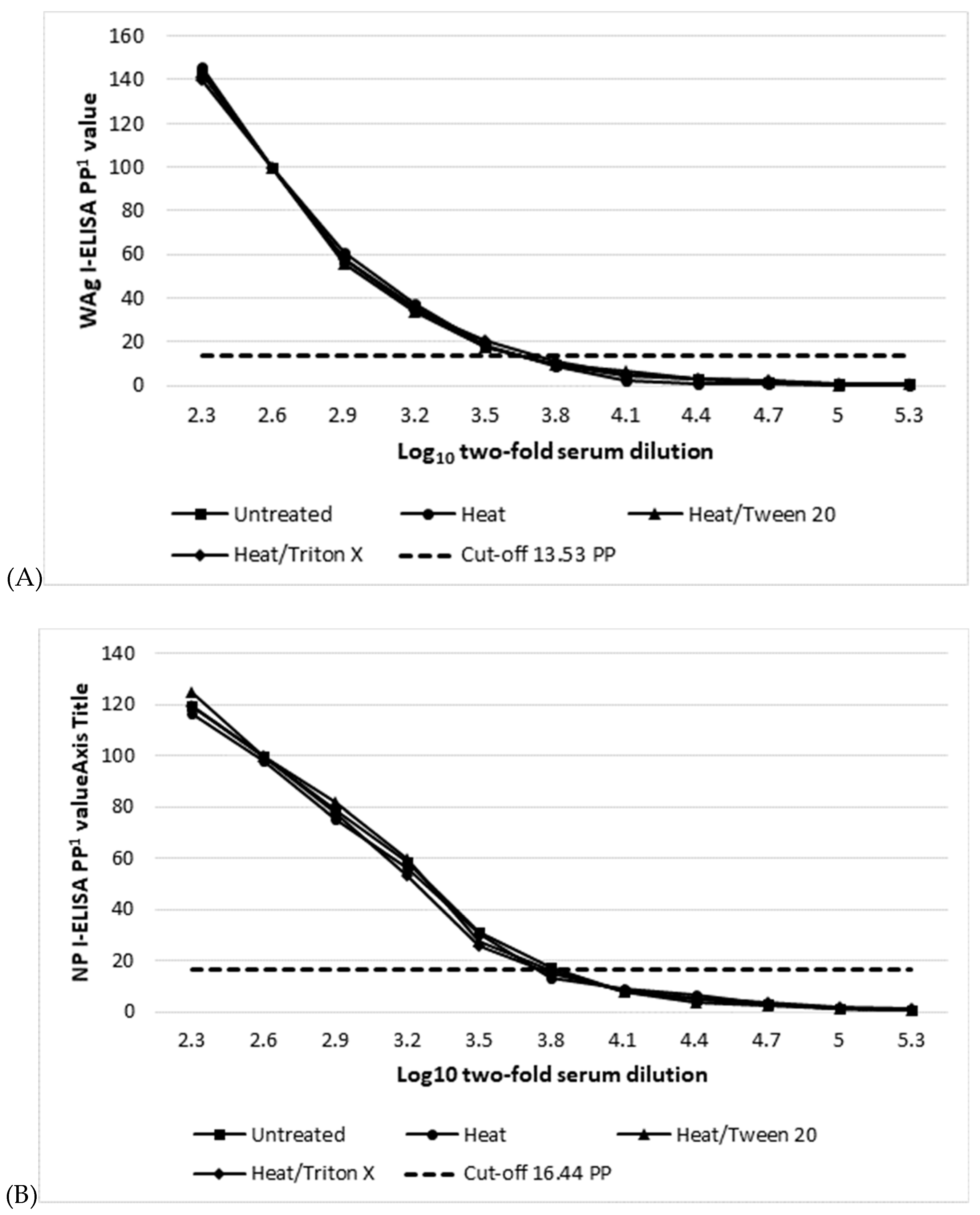
2.5. Serum Inactivation
3. Results
3.1. IQC and Repeatability
3.2. Distribution of I-ELISA PP Values in EBOV IgG Negative Sera and the Selection of Cut-off Values
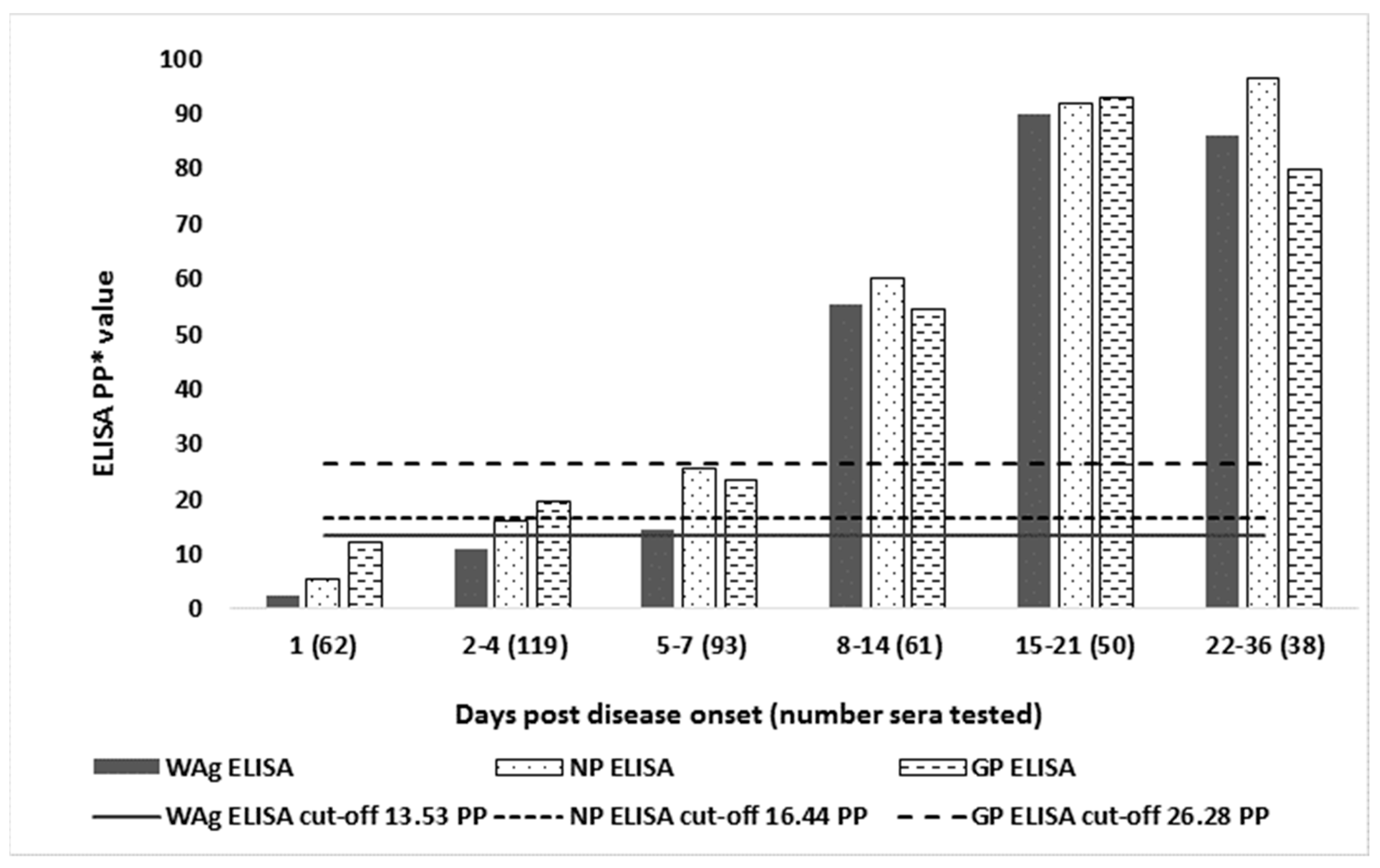
3.3. Comparison of D-Sp and D-Se
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kuhn, J.H.; Adachi, T.; Adhikari, N.K.J.; Arribas, J.R.; Bah, I.E.; Bausch, D.G.; Bhadelia, N.; Borchert, M.; Broch Brantsaeter, A.; Brett-Major, D.M.; et al. New filovirus disease classification and nomenclature. Nat. Rev. Microbiol. 2019, 17, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Muyembe-Tamfum, J.J.; Mulangu, S.; Masumu, J.; Kayembe, J.M.; Kemp, A.; Paweska, J.T. Ebola virus outbreaks in Africa: Past and present. Onderstepoort J. Vet. Res. 2012, 79, 451. [Google Scholar] [CrossRef] [PubMed]
- Baize, S.; Pannetier, D.; Oestereich, L.; Rieger, T.; Koivogui, L.; Magassouba, N.; Soropogui, B.; Sow, M.S.; Keita, S.; de Clerck, H.; et al. Emergence of Zaire Ebola virus disease in Guinea. N. Engl. J. Med. 2014, 371, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Briand, S.; Bertherat, E.; Cox, P.; Formenty, P.; Kieny, M.P.; Myhre, J.K.; Roth, C.; Shindo, N.; Dye, C. The international Ebola emergency. N. Engl. J. Med. 2014, 371, 1180–1183. [Google Scholar] [CrossRef] [PubMed]
- WHO Ebola Response Team. Ebola virus disease in West Africa—the first 9 months of the epidemic and forward projections. N. Engl. J. Med. 2014, 371, 1481–1495. [Google Scholar] [CrossRef]
- World Health Organization—Ebola virus Disease—Democratic Republic of the Congo. Available online: https://www.who.int/csr/don/07-february-2019-ebola-drc/en/ (accessed on 11 February 2019).
- Médecins Sans Frontières. DRC Ebola outbreak crisis update—April 2019. Available online: https://www.msf.org/drc-2018-ebola-outbreak-crisis-update (accessed on 18 April 2018).
- Epstein, H. 11 313 unnecessary Ebola-related deaths: Building citizen trust in health systems. Brenthurst Foundation Special Report 1/2016. Sheaf Publishing: Benoni, South Africa. Available online: www.thebrenthurstfoundation.org/Brenthurst-special-report-201601.p (accessed on 20 May 2019).
- World Bank Group. The economic impact of the 2014 Ebola epidemic: Short- and medium-term estimates for West Africa. Available online: https://openknowlege.worldbank.org/handle/10986/20592. (accessed on 20 May 2019).
- Evans, D.K.; Goldstein, M.; Popova, A. Health-care worker mortality and the legacy of the Ebola epidemic. Lancet Glob. Health 2015, 3, e439–e440. [Google Scholar] [CrossRef] [Green Version]
- Gostin, L.O.; Fridman, A.A. A retrospective and prospective analysis of the West African Ebola virus disease epidemic: Robust national health systems at the foundation and an empowered WHO at the apex. Lancet 2015, 385, 1902–1909. [Google Scholar] [CrossRef]
- Shoman, H.; Karafillakis, E.; Rawaf, S. The link between the West African Ebola outbreak and health systems in Guinea, Liberia and Sierra Leone: A systematic review. Glob. Health 2017, 13, 1. [Google Scholar] [CrossRef]
- Vinck, P.; Pham, P.N.; Bindu, K.K.; Bedford, J.; Nilles, E. Institutional trust and misinformation in the response to the 2018-19 Ebola outbreak in North Kivu, DR Congo; a population-based survey. Lancet Infect. Dis. 2019, 19, 529–536. [Google Scholar] [CrossRef]
- Gostin, L.; Phelan, A.; Coutinho, A.G.; Eccleston-Turner, M.; Erondu, N.; Filani, O.; Inglesby, T.; Katz, R.; Maleche, A.; Nuzzo, B.; et al. Ebola in the Democratic Republic of the Congo; time to sound a global alert? Lancet Infect. Dis. 2019, 393, 617–620. [Google Scholar] [CrossRef]
- Jansen van Vuren, P.; Grobbelaar, A.; Storm, N.; Conteh, O.; Konneh, K.; Kamara, A.; Sanne, I.; Paweska, J.T. Comparative evaluation of the diagnostic performance of the prototype Cepheid GeneXpert Ebola assay. J. Clin. Microbiol. 2016, 54, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Raftery, P.; Condell, O.; Wasunna, C.; Kpaka, J.; Zwizwai, R.; Nuha, M.; Fallah, M.; Freemen, M.; Harris, V.; Miller, M.; et al. Establishing Ebola virus disease (EVD) diagnostics using GeneXpert technology at a mobile laboratory in Liberia: Impact on outbreak response, case management and laboratory systems strengthening. PLoS Negl. Trop. Dis. 2018, 12, e0006135. [Google Scholar] [CrossRef] [PubMed]
- Phan, J.C.; Pettitt, J.; George, J.S.; Fakoli, L.S., III; Taweh, F.M.; Bateman, S.L.; Bennett, R.S.; Norris, S.L.; Spinnler, D.A.; Pimentel, G.; et al. Lateral flow immunoassays for Ebola virus disease detection in Liberia. J. Infect. Dis. 2016, 214, S222–S228. [Google Scholar] [CrossRef] [PubMed]
- Wonderly, B.; Jones, S.; Gatton, M.L.; Barber, J.; Killip, M.; Hudson, C.; Carter, L.; Sipson, A.J.H.; Urassa, W.; Chua, A.; et al. Comparative performance of four rapid Ebola antigen-detection lateral flow immunoassays during the 2014-2016 Ebola epidemic in West Africa. PLoS ONE 2019, 7, e0212113. [Google Scholar] [CrossRef] [PubMed]
- MacNeil, A.; Reed, Z.D.; Rollin, P.E. Serologic cross-reactivity of human IgM and IgG antibodies to five species of Ebola virus. PLoS Negl. Trop. Dis. 2011, 5, e1175. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, Y.; Wu, W.; Li, A.; Yang, X.; Zhang, S.; Li, C.; Su, Q.; Cai, S.; Sun, D.; et al. Serological investigation of laboratory-confirmed and suspected Ebola virus disease patients during the late phase of the Ebola outbreak in Sierra Leone. Virol. Sin. 2018, 33, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Cross, R.W.; Ksiazek, T.G. Production of antigens for ELISA. In Ebolaviruses. Methods in Molecular Biology; Hoenen, T., Groseth, A., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1628, pp. 353–362. [Google Scholar]
- Niikura, M.; Ikegami, T.; Saijo, M.; Kurata, T.; Kurane, I.; Morikawa, S. Analysis of linear B-cell epitopes of the nucleoprotein of Ebola virus that distinguish Ebola virus subtypes. Clin. Diagn. Lab. Immunol. 2003, 10, 83–87. [Google Scholar] [CrossRef]
- Sanchez, A.; Kiley, M.P.; Holloway, B.P.; Auperin, D.D. Sequence analysis of the Ebola virus genome: Organization, genetic elements, and comparison with the genome of Marburg virus. Virus Res. 1993, 29, 215–240. [Google Scholar] [CrossRef]
- Saijo, M.; Niikura, M.; Morikawa, S.; Ksiazek, T.G.; Meyer, R.F.; Peters, C.J.; Kurane, I. Enzyme-linked immunosorbent assays for detection of antibodies to Ebola and Marburg viruses using recombinant nucleoproteins. J. Clin. Microbiol. 2001, 39, 1–7. [Google Scholar] [CrossRef]
- Natesan, M.; Jensen, S.M.; Keasey, S.L.; Kamata, T.; Kuehne, I.E.; Stonier, S.W.; Lutwama, J.J.; Lobel, L.; Dye, J.M.; Ulrich, R.G. Human survivors of disease outbreaks caused by Ebola or Marburg virus exhibit cross-reactive and long-lived antibody responses. Clin. Vaccine Immunol. 2016, 23, 717–724. [Google Scholar] [CrossRef]
- Nakayama, E.; Yokoyama, A.; Miyamoto, H.; Igarashi, M.; Kishida, N.; Matsuno, K.; Marzi, A.; Feldmann, H.; Ito, K.; Saijo, M.; et al. Enzyme-linked immunosorbent assay for detection of filovirus species-specific antibodies. Clin. Vaccine Immunol. 2010, 17, 1723–1728. [Google Scholar] [CrossRef] [PubMed]
- Takada, A.; Ebihara, H.; Jones, S.; Feldmann, H.; Kawaoka, Y. Protective efficacy of neutralizing antibodies against Ebola virus infection. Vaccine 2007, 25, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Prehaud, C.; Hellebrand, E.; Coudrier, D.; Volchkov, V.E.; Volchkova, V.A.; Feldmann, H.; Le Guenno, B.; Bouloy, M. Recombinant Ebola virus nucleoprotein and glycoprotein (Gabon 94 strain) provide new tools for the detection of human infections. J. Gen. Virol. 1998, 79, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
- Groen, J.; van den Hoogen, B.G.; Burghoorn-Maas, C.P.; Fooks, A.R.; Burton, J.; Clegg, C.J.; Zeller, H.; Osterhaus, A.D. Serological reactivity of baculovirus-expressed Ebola virus VP35 and nucleoproteins. Microbes Infect. 2003, 5, 379–385. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, Y.; Yang, M.; Zhang, Z.; Song, D.; Yuan, Z. Nucleoprotein-based indirect enzyme-linked immunosorbent assay (indirect ELISA) for detecting antibodies specific to Ebola virus and Marburg virus. Virol. Sin. 2014, 29, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.; Shulenin, S.; Grolla, A.; Audet, J.; He, S.; Kobinger, G.; Unfer, R.C.; Warfield, K.L.; Aman, M.J.; Holtsberg, F.W. Quantitative serology assays for determination of antibody responses to Ebola virus glycoprotein and matrix protein in nonhuman primates and humans. Antiviral Res. 2016, 126, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef]
- Jarvis, D.L.; Kawar, Z.S.; Hollister, J.R. Engineering N-glycosylation pathways in the baculovirus-insect cell system. Curr. Opin. Biotechnol. 1998, 9, 528–533. [Google Scholar] [CrossRef]
- Glynn, J.R.; Bower, H.; Johnson, S.; Houlihan, C.F.; Montesano, C.; Scott, J.T.; Semple, M.G.; Bangura, M.S.; Kamara, A.J.; Kamara, O.; et al. Asymptomatic infection and unrecognised Ebola virus disease in Ebola-affected households in Sierra Leone: A cross-sectional study using a new non-invasive assay for antibodies to Ebola virus. Lancet Infect. Dis. 2017, 17, 645–653. [Google Scholar] [CrossRef]
- Shurtleff, A.C.; Biggins, J.E.; Keeney, A.E.; Zumbrun, E.E.; Bloomfield, H.A.; Kuehne, A.; Audet, J.L.; Alfson, K.J.; Griffiths, A.; Olinger, G.G.; et al. Standardization of the filovirus plaque assay for use in preclinical studies. Viruses 2012, 4, 3511–3530. [Google Scholar] [CrossRef]
- Paweska, J.T.; Jansen van Vuren, P.; Meier, G.H.; Le Roux, C.; Conteh, O.S.; Kemp, A.; Fourie, C.; Naidoo, P.; Naicker, S.; Ohaebosim, P.; et al. South African Ebola diagnostic response in Sierra Leone: A modular high biosafety field laboratory. PLoS Negl. Trop. Dis. 2017, 11, e0005665. [Google Scholar] [CrossRef] [PubMed]
- Panning, M.; Laue, T.; Olschlager, S.; Eickmann, M.; Becker, S.; Raith, S.; Georges Courbot, M.C.; Nilsson, M.; Gopal, R.; Lundkvist, A.; et al. Diagnostic reverse-transcription polymerase chain reaction kit for filoviruses based on the strain collections of all European biosafety level 4 laboratories. J. Infect. Dis. 2007, 196, S199–S204. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Ebola hemorrhagic fever—South Africa. Wkly. Epidemiol. Rec. 1996, 71, 359. [Google Scholar]
- Richards, G.A.; Murphy, S.; Jobson, R.; Mer, M.; Zinman, C.; Taylor, R.; Swanepoel, R.; Duse, A.; Sharp, G.; de la Rey, I.C.J.; et al. Unexpected Ebola virus in a tertiary setting: Clinical and epidemiological aspects. Crit. Care Med. 2000, 28, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Ksiazek, T.G.; West, C.P.; Rollin, P.E.; Jahrling, P.B.; Peters, C.J. ELISA for the detection of antibodies to Ebola viruses. J. Infect. Dis. 1999, 179, S192–S198. [Google Scholar] [CrossRef]
- Paweska, J.T.; Burt, F.J.; Swanepoel, R. Validation of IgG-sandwich and IgM-capture ELISA for the detection of antibody to Rift Valley fever virus in humans. J. Virol. Methods 2005, 124, 173–181. [Google Scholar] [CrossRef]
- Crowther, R. Elisa Theory and Practice, in Methods in Molecular Biology; Crowther, R., Ed.; Humana Press Inc.: Tatowa, NJ, USA, 1995. [Google Scholar]
- Greiner, M. Two-graph receiver operating characteristics (TG-ROC)—A Microsoft-Excel template for the selection of cut-off values in diagnostic tests. J. Immunol. Methods 1995, 185, 123–132. [Google Scholar] [CrossRef]
- Greiner, M. Two-graph receiver operating characteristics (TG-ROC)—A Microsoft-Excel template for the selection of cut-off values that minimize overall misclassification costs. J. Immunol. Methods 1996, 191, 93–94. [Google Scholar] [CrossRef]
- Greiner, M.; Gardner, I.A. Application of diagnostic tests in veterinary epidemiological studies. Prev. Vet. Med. 2000, 45, 43–59. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef]
- Mitchell, S.W.; McCormick, J.B. Physicochemical inactivation of Lassa, Ebola and Marburg viruses and effect on clinical laboratory analyses. J. Clin. Microbiol. 1984, 20, 486–489. [Google Scholar] [PubMed]
- Haddock, E.; Feldmann, F.; Feldmann, H. Effective chemical inactivation of Ebola virus. Emerg. Infect. Dis. 2016, 22, 1292–1294. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, R.H. Validation of serological assays for diagnosis infectious diseases. Rev. Sci. Tech. Off. Int. Epiz. 1998, 17, 469–486. [Google Scholar] [CrossRef]
- Jacobson, R.H. Principles of validation of diagnostic assays for infectious diseases. In OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 5th ed.; Cambridge University Press: Cambridge, UK, 2004; Volume 1, pp. 21–29. [Google Scholar]
- Vizard, A.L.; Anderson, G.A.; Gasser, R.B. Determination of the optimum cut-off value of a diagnostic test. Prev. Vet. Med. 1990, 10, 137–143. [Google Scholar] [CrossRef]
- Ksiazek, T.G.; Rollin, P.E.; Williams, A.J.; Bressler, D.S.; Martin, M.L.; Swanepoel, R.; Burt, F.J.; Leman, P.A.; Khan, A.S.; Rowe, A.K.; et al. Clinical virology of Ebola hemorrhagic fever (EHF): Virus, virus antigen, and IgG and IgM antibody findings among EHF patients in Kiwit, Democratic Republic of the Congo, 1995. J. Infect. Dis. 1999, 179, S177–S187. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.F.; Nilsson, E.; van Rooij, E.M.A.; Lelenta, M.; Jeggo, M.H. Standarization and validation of enzyme-linked immunosorbent assay techniques for the detection of antibody in infectious disease diagnosis. Rev. Sci. Tech. Off. Int. Epiz. 1993, 12, 435–450. [Google Scholar] [CrossRef]
- Krähling, V.; Becker, D.; Rohde, C.; Eickmann, M.; Eroglu, Y.; Herwig, A.; Kerber, R.; Kowalski, K.; Vergara-Alert, J.; Becker, S.; et al. Development of an antibody capture ELISA using inactivated Ebola Zaire Makona virus. Med. Microbiol. Immunol. 2016, 205, 173–183. [Google Scholar] [CrossRef]
- Sobarzo, A.; Perelman, E.; Groseth, A.; Dolnik, O.; Becker, S.; Lutwama, J.J.; Dye, J.M.; Yavelsky, V.; Lobel, L.; Marks, R.S. Profiling the native specific human humoral immune response to Sudan ebolavirus strain Gulu by chemiluminescence enzyme-linked immunosorbent assay. Clin. Vaccine Immunol. 2012, 19, 1844–1852. [Google Scholar] [CrossRef]
- Lopez, L.; Venteo, A.; Garcia, M.; Camuñas, A.; Ranz, A.; Garcia, J.; Sarraseca, J.; Anaya, C.; Rueda, P. Antigen-capture blocking enzyme-linked immunosorbent assay based on a baculovirus recombinant antigen to differentiate transmissable gastroenteritis virus from porcine respiratory coronavirus antibodies. J. Vet. Diagn. Investig. 2009, 21, 598–608. [Google Scholar] [CrossRef]
- Rowe, A.K.; Bertolli, J.; Khan, A.S.; Mukunu, R.; Muyembe-Tamfum, J.J.; Bressler, D.; Williams, A.J.; Peters, C.J.; Rodriguez, L.; Feldmann, H.; et al. Clinical, virologic and immunologic follow-up of convalescent Ebola hemorrhagic fever patientsand their household contacts, Kikwit, Democratic Republic of the Congo. J. Infect. Dis. 1999, 179, S28–S35. [Google Scholar] [CrossRef]
- Wauquier, N.; Becquart, P.; Gasquet, C.; Leroy, E.M. Immunoglobulin G in Ebola outbreak survivors, Gabon. Emerg. Infect. Dis. 2009, 15, 1136–1137. [Google Scholar] [CrossRef] [PubMed]
- Rudge, T.L., Jr.; Sankovich, K.A.; Niemuth, N.A.; Anderson, M.S.; Badorrek, C.S.; Skomrock, N.D.; Cirimotich, C.M.; Carol, L.; Sabourin, C.L. Development, qualification, and validation of the Filovirus Animal Nonclinical Group anti-Ebola virus glycoprotein immunoglobulin G enzyme-linked immunosorbent assay for human serum samples. PLoS ONE 2019, 14, e0215457. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C++ 1 | OD 2 IQC 3 | Mean OD ± SD 4 | Mean CV 5 ± SD (%) | |||
|---|---|---|---|---|---|---|
| LCL 6 | UCL 7 | Intra-Plate Variation | Inter-Plate Variation | Intra-Plate Variation | Inter-Plate Variation | |
| WAg | 0.81 | 1.29 | 1.00 ± 0.09 | 1.00 ± 0.08 | 4.26 ± 2.67 | 4.41 ± 1.85 |
| NP | 1.14 | 1.79 | 1.45 ± 0.18 | 1.50 ± 0.10 | 3.54 ± 1.99 | 3.11 ± 1.05 |
| GP | 0.9 | 1.39 | 1.19 ± 0.09 | 1.2 ± 0.04 | 4.76 ± 2.16 | 4.8 ± 1.56 |
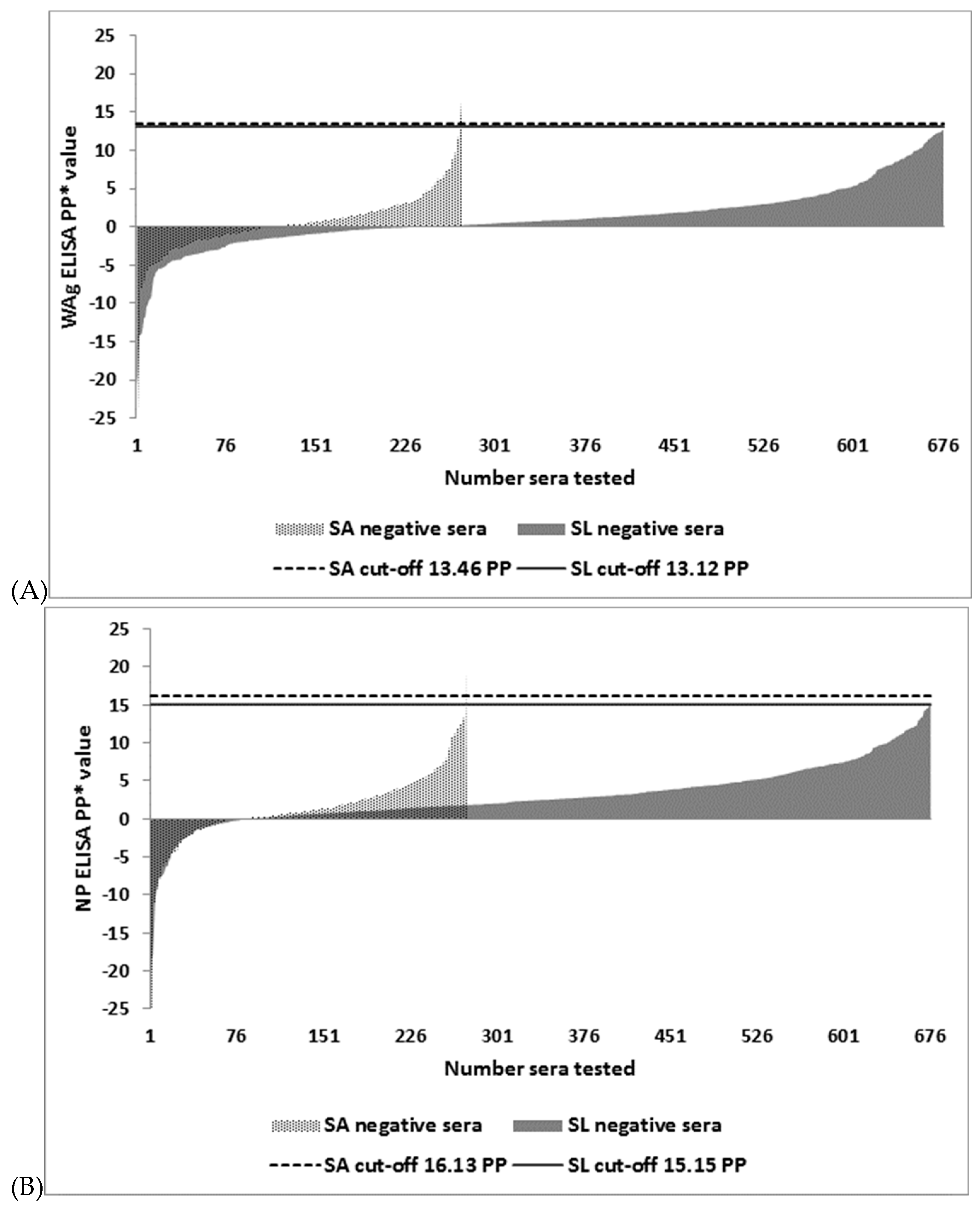
| Test | Cut-off PP 1 | SA Negative Serum Panel (n = 273) | SL Negative Serum Panel (n = 676) | ||
|---|---|---|---|---|---|
| FP 2/TN 3 | DSp 4 (95% CI 5) | FP/TN | DSp (95% CI) | ||
| WAg I-ELISA | 13.46 6 | 2/271 | 99.3 (97.4–99.9) | 0/676 | 100 (99.5–100) |
| WAg I-ELISA | 13.12 7 | 2/271 | 99.3 (97.4–99.9) | 0/676 | 100 (99.5–100) |
| WAg I-ELISA | 13.53 8 | 2/271 | 99.3 (97.4–99.9) | 0/676 | 100 (99.5–100) |
| NP I-ELISA | 16.13 6 | 1/272 | 99.6 (98–100) | 0/676 | 100 (99.5–100) |
| NP I-ELISA | 15.15 7 | 1/272 | 99.6 (98–100) | 2/674 | 99.7 (98.9–100) |
| NP I-ELISA | 16.44 8 | 1/272 | 99.6 (98–100) | 0/676 | 100 (99.5–100) |
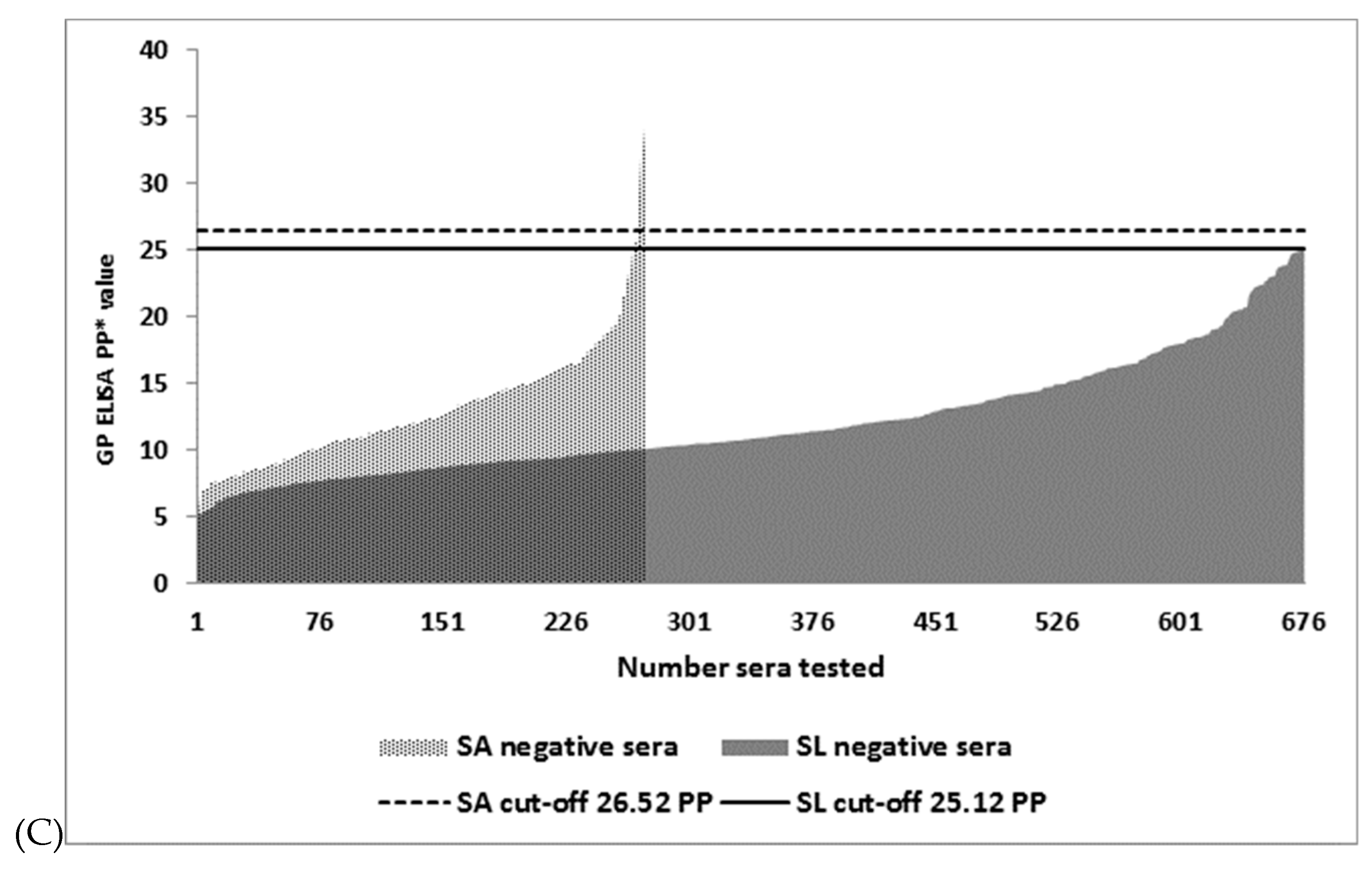
| GP I-ELISA | 26.52 6 | 5/268 | 98.2 (95.8–99.4) | 0/676 | 100 (99.5–100) |
| GP I-ELISA | 25.12 7 | 7/268 | 97.4 (94.8–99) | 2/674 | 99.7 (98.9–100) |
| GP I-ELISA | 26.28 8 | 5/268 | 98.2 (95.8–99.4) | 0/676 | 100 (995–100) |
| Days Post Disease Onset (Number Individuals Tested) | WAg No.pos. 1 (% pos.) | NP No. pos.(% pos.) | GP No. pos.(% pos.) | Only WAg pos. | Wag and NP pos. | Wag and GP pos. | Only NP pos. | NP and GP pos. | Only GP pos. |
|---|---|---|---|---|---|---|---|---|---|
| 1 (n = 62) | 3 2 (4.8) | 4 (6.5) | 1 (1.6) | 0 | 3 | 0 | 0 | 1 | 0 |
| 3 3 (4.8) | 5 (8.1) | 2 (3.2) | 0 | 3 | 0 | 1 | 1 | 1 | |
| 3 4 (4.8) | 4 (6.5) | 1 (1.6) | 0 | 3 | 0 | 0 | 1 | 0 | |
| 2–4 (n = 119) | 20 (16.8) | 27 (22.7) | 13 (11.0) | 1 | 6 | 0 | 8 | 0 | 0 |
| 20 (16.8) | 28(23.5) | 13 (11.0) | 1 | 6 | 0 | 9 | 0 | 0 | |
| 20 (16.8) | 27 (22.7) | 13 (11.0) | 1 | 6 | 0 | 8 | 0 | 0 | |
| 5–7 (n = 93) | 27 (29.0) | 34 (36.6) | 24 (25.8) | 1 | 7 | 0 | 6 | 2 | 3 |
| 27 (29.0) | 34 (36.6) | 26 28.0) | 1 | 7 | 0 | 5 | 3 | 4 | |
| 27 (29.0) | 34 (36.6) | 24 (25.8) | 1 | 7 | 0 | 6 | 2 | 3 | |
| 8–14 (n = 61) | 42 (68.9) | 49 (80.3) | 37 (60.7) | 1 | 8 | 0 | 5 | 4 | 0 |
| 43 (70.5) | 50 (82.0) | 39 (64.0) | 1 | 9 | 0 | 3 | 6 | 0 | |
| 42 (68.9) | 49 (80.3) | 37 (60.7) | 1 | 8 | 0 | 5 | 4 | 0 | |
| 15–21 (n = 50) | 50 (100) | 50 (100) | 49 (98.0) | 0 | 0 | 0 | 0 | 0 | 0 |
| 50 (100) | 50 (100) | 49 (98.0) | 0 | 0 | 0 | 0 | 0 | 0 | |
| 50 (100) | 50(100) | 49 (98.0) | 0 | 0 | 0 | 0 | 0 | 0 | |
| 22–36 (n = 38) | 38 (100) | 38 (100) | 37 (97.4) | 0 | 0 | 0 | 0 | 0 | 0 |
| 38 (100) | 38(100) | 38 (100) | 0 | 0 | 0 | 0 | 0 | 0 | |
| 38 (100) | 38 (100) | 37 (97.4) | 0 | 0 | 0 | 0 | 0 | 0 |
| Test | Cut-off PP 1 | SL EBOV RT-PCR Positive Cases Days 8–36 Post Onset (n = 149) | SL EBOV RT-PCR Positive Cases Days 15–36 Post Onset (n = 88) | ||
|---|---|---|---|---|---|
| FN 2/TP 3 | DSe 4 (95% CI 5) | FN/TP | DSe (95% CI) | ||
| WAg I-ELISA | 13.46 6 | 19/130 | 87.2 (80.8–92.1) | 0/88 | 100 (95.9–100) |
| WAg I-ELISA | 13.12 7 | 18/131 | 87.9 (81.6–92.7) | 0/88 | 100 (95.5–100) |
| WAg I-ELISA | 13.53 8 | 19/130 | 87.2 (80.8–92.1) | 0/88 | 100 (95.5–100) |
| NP I-ELISA | 16.13 6 | 12/137 | 91.9 (86.4–95.8) | 0/88 | 100 (95.5–100) |
| NP I-ELISA | 15.15 7 | 11/138 | 92.6 (87.2–96.3) | 0/88 | 100 (95.9–100) |
| NP I-ELISA | 16.44 8 | 12/137 | 91.9 (86.4–95.8) | 0/88 | 100 (95.5–100) |
| GP I-ELISA | 26.52 6 | 26/123 | 82.6 (75.5–88.3) | 2/86 | 97.7 (92–99.7) |
| GP I-ELISA | 25.12 7 | 23/126 | 84.6 (77.7–90) | 1/87 | 98.9 (93.8–100) |
| GP I-ELISA | 26.28 8 | 26/123 | 82.6 (75.5–88.3) | 2/86 | 97.7 (92–99.7) |
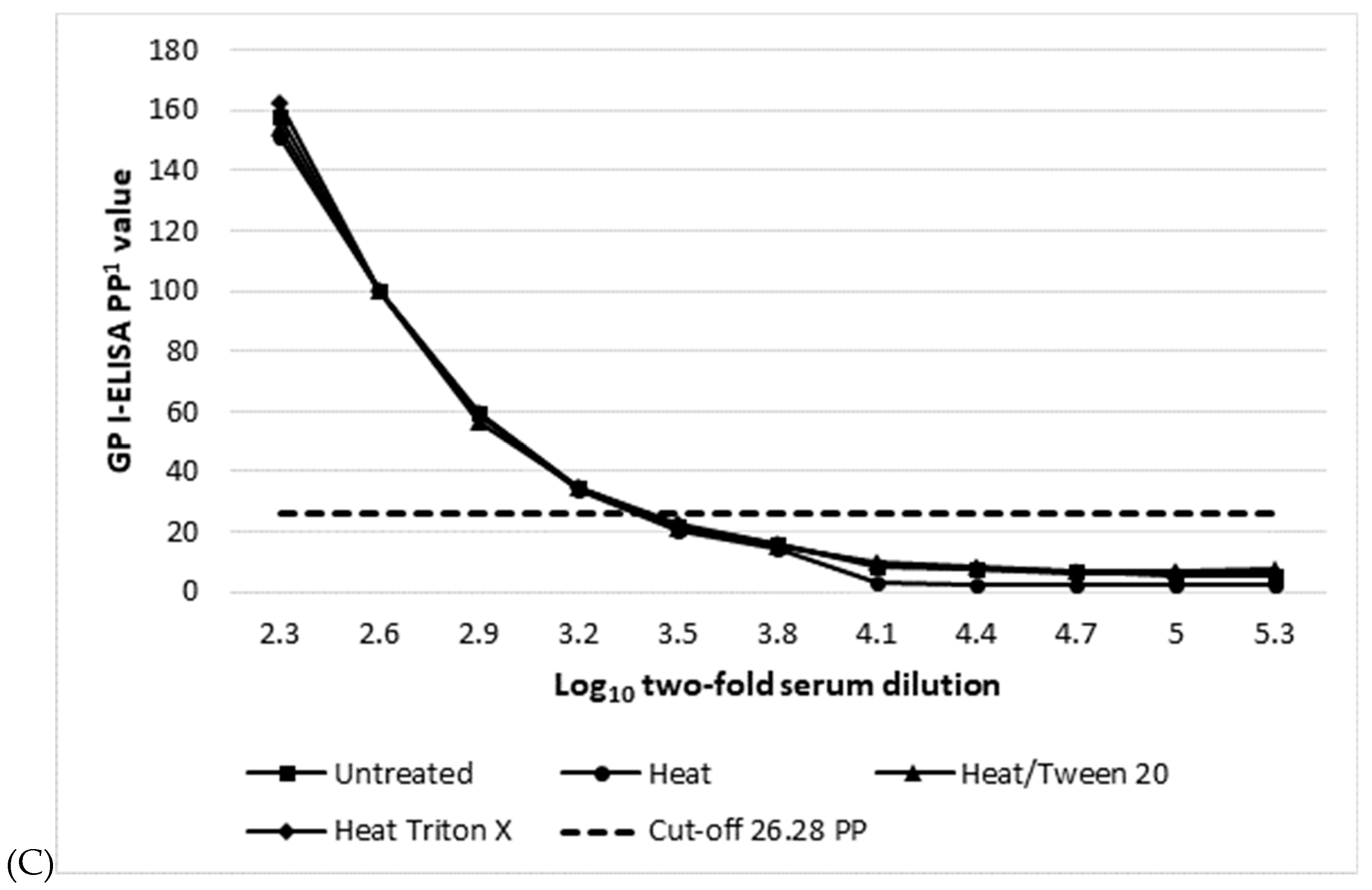
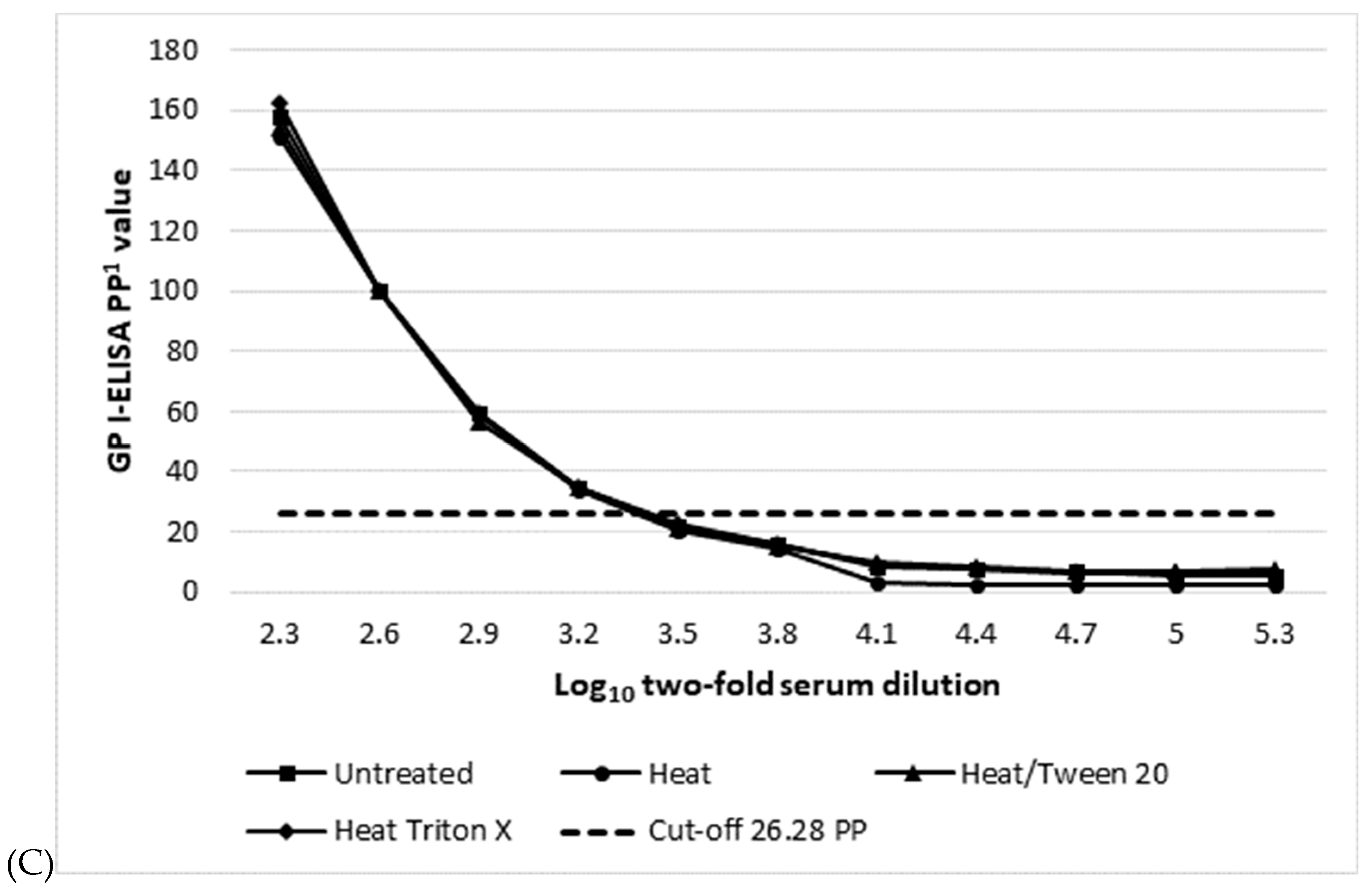
| Assay (Cut-off PP Value) 1 | Mean log10 Serum Titre 2/Dose Response Curve R Square 3 | |||
|---|---|---|---|---|
| Untreated | 60° 1 h | 0.5%Tween 20 15 min 60° | 0.5%Triton X-100 15 min 60° | |
| WAg I-ELISA (13.53) | 3.5/0.945 | 3.5/0.950 | 3.5/0.942 | 3.5/0.952 |
| NP I-ELISA (16.44 PP) | 3.8/0.959 | 3.5/0.961 | 3.5/0.956 | 3.5/0.957 |
| GP I-ELISA (26.28) | 3.2/0.932 | 3.2/0.942 | 3.2/0.924 | 3.2/0.928 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paweska, J.T.; Moolla, N.; Storm, N.; Msimang, V.; Conteh, O.; Weyer, J.; Vuren, P.J.v. Evaluation of Diagnostic Performance of Three Indirect Enzyme-Linked Immunosorbent Assays for the Detection of IgG Antibodies to Ebola Virus in Human Sera. Viruses 2019, 11, 678. https://doi.org/10.3390/v11080678
Paweska JT, Moolla N, Storm N, Msimang V, Conteh O, Weyer J, Vuren PJv. Evaluation of Diagnostic Performance of Three Indirect Enzyme-Linked Immunosorbent Assays for the Detection of IgG Antibodies to Ebola Virus in Human Sera. Viruses. 2019; 11(8):678. https://doi.org/10.3390/v11080678
Chicago/Turabian StylePaweska, Janusz T., Naazneen Moolla, Nadia Storm, Veerle Msimang, Ousman Conteh, Jacqueline Weyer, and Petrus Jansen van Vuren. 2019. "Evaluation of Diagnostic Performance of Three Indirect Enzyme-Linked Immunosorbent Assays for the Detection of IgG Antibodies to Ebola Virus in Human Sera" Viruses 11, no. 8: 678. https://doi.org/10.3390/v11080678
APA StylePaweska, J. T., Moolla, N., Storm, N., Msimang, V., Conteh, O., Weyer, J., & Vuren, P. J. v. (2019). Evaluation of Diagnostic Performance of Three Indirect Enzyme-Linked Immunosorbent Assays for the Detection of IgG Antibodies to Ebola Virus in Human Sera. Viruses, 11(8), 678. https://doi.org/10.3390/v11080678