Two Novel Negative-Sense RNA Viruses Infecting Grapevine Are Members of a Newly Proposed Genus within the Family Phenuiviridae

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Total Nucleic Acid Extraction
2.2. High Throughput Sequencing and Completion of Full Genomes
2.3. Accession Numbers
2.4. Genome Analysis
2.5. Phylogenetic Analyses
2.6. Detection, Survey, and Biological Assay
3. Results
3.1. Identification of Two Novel nsRNA Viruses Infecting Grapevine by HTS
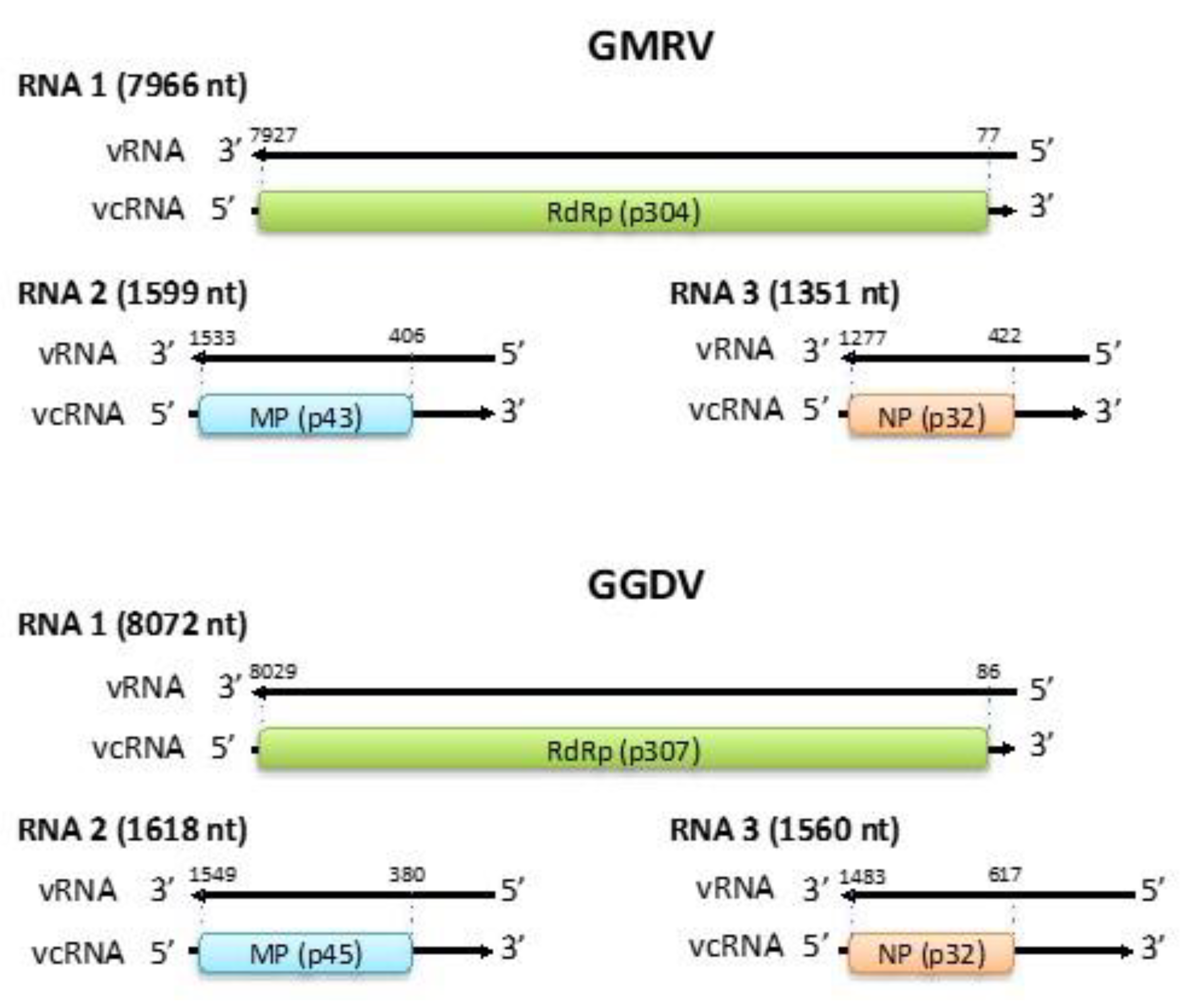
3.2. Genomic Organization of GMRV and GGDV
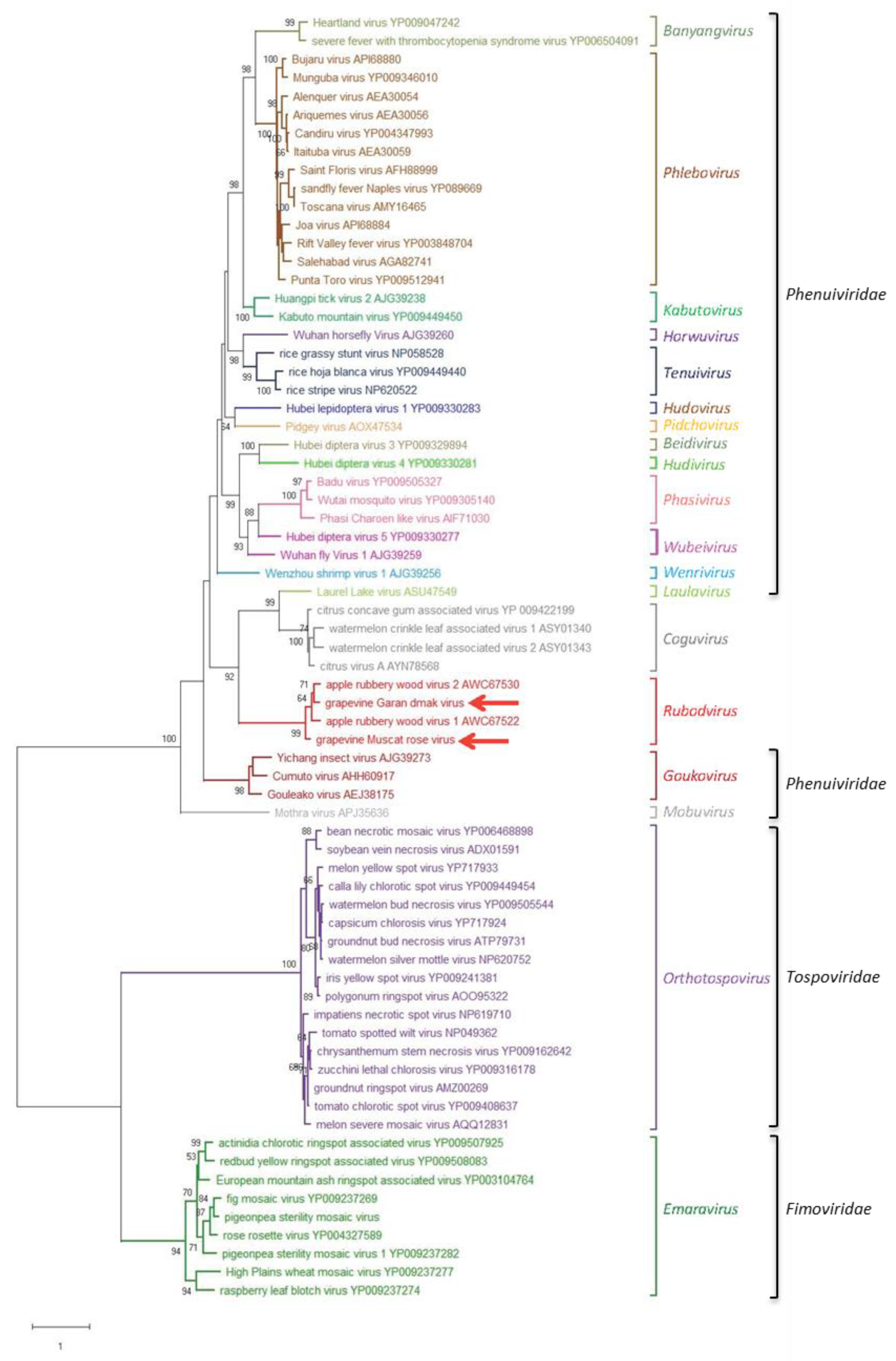
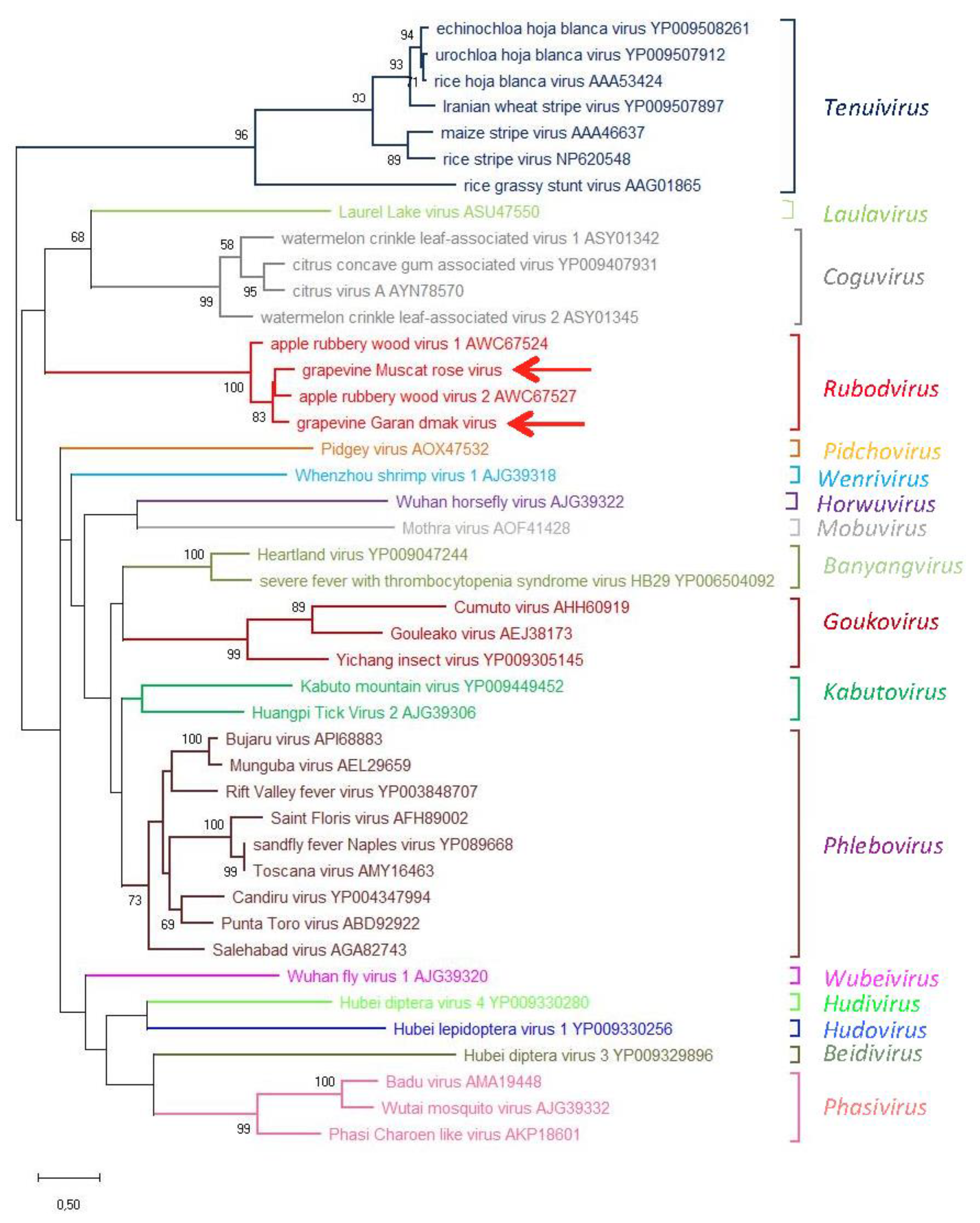
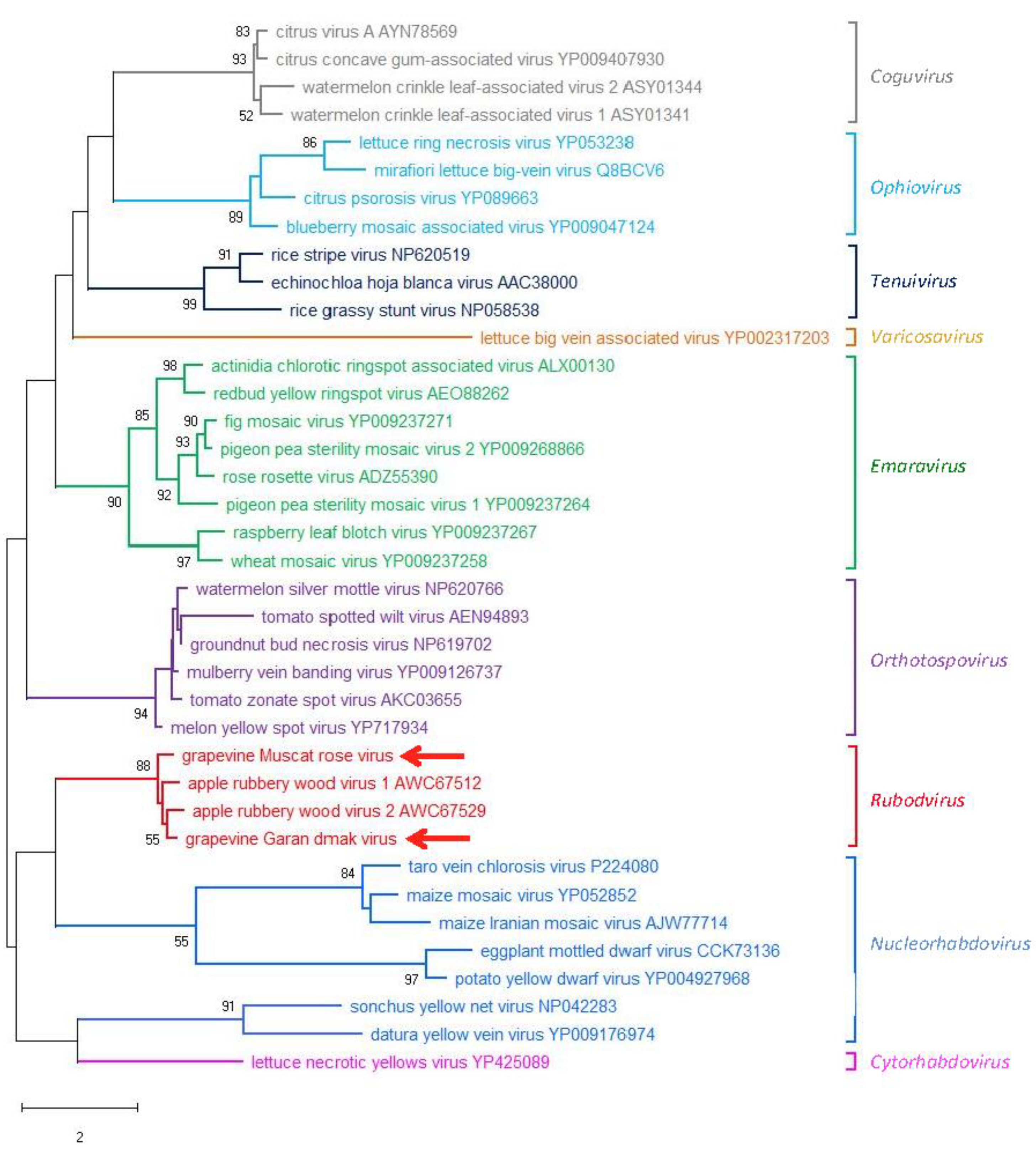
3.3. Phylogenetic Relationships of GMRV and GGDV with Other nsRNA Viruses
3.4. Detection and Prevalence of Novel nsRNA Viruses
3.5. Graft-Transmission of GMRV and GGDV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wolf, Y.; Krupovic, M.; Zhang, Y.Z.; Maes, P.; Dolja, V.; Koonin, E.V.; Kuhn, J.H. Proposal 2017.016 M.A.v2. Megataxonomy of Negative-Sense RNA Viruses. Available online: https://talk.ictvonline.org/ICTV/proposals/2017.006M.R.Negarnaviricota.zip (accessed on 15 October 2018).
- Luo, M. Negative Strand RNA Virus; World Scientific: Singapore, 2011. [Google Scholar]
- Laney, A.G.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. A discovery 70 years in the making: Characterization of the Rose rosette virus. J. Gen. Virol. 2011, 92, 1727–1732. [Google Scholar] [CrossRef] [PubMed]
- Toriyama, S. Rice stripe virus: Prototype of a new group of viruses that replicate in plants and insects. Microbiol. Sci. 1986, 3, 347–351. [Google Scholar] [PubMed]
- Barthe, G.A.; Ceccardi, T.L.; Manjunath, K.L.; Derrick, K.S. Citrus psorosis virus: Nucleotide sequencing of the coat protein gene and detection by hybridization and RT-PCR. J. Gen. Virol. 1998, 79, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Thekke-Veetil, T.; Ho, T.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. A new ophiovirus is associated with blueberry mosaic disease. Virus Res. 2014, 189, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Kormelink, R.; Garcia, M.L.; Goodin, M.; Sasaya, T.; Haenni, A.-L. Negative-strand RNA viruses: The plant-infecting counterparts. Virus Res. 2011, 162, 184–202. [Google Scholar] [CrossRef]
- Buzkan, N.; Chiumenti, M.; Massart, S.; Sarpkaya, K.; Karadağ, S.; Minafra, A. A new emaravirus discovered in Pistacia from Turkey. Virus Res. 2019, 263, 159–163. [Google Scholar] [CrossRef]
- Navarro, B.; Minutolo, M.; Stradis, A.D.; Palmisano, F.; Alioto, D.; Serio, F.D. The first phlebo-like virus infecting plants: A case study on the adaptation of negative-stranded RNA viruses to new hosts. Mol. Plant Pathol. 2018, 19, 1075–1089. [Google Scholar] [CrossRef]
- Navarro, B.; Zicca, S.; Minutolo, M.; Saponari, M.; Alioto, D.; Di Serio, F. A Negative-Stranded RNA Virus Infecting Citrus Trees: The Second Member of a New Genus Within the Order Bunyavirales. Front. Microbiol. 2018, 9, 2340. [Google Scholar] [CrossRef]
- Xin, M.; Cao, M.; Liu, W.; Ren, Y.; Zhou, X.; Wang, X. Two Negative-Strand RNA Viruses Identified in Watermelon Represent a Novel Clade in the Order Bunyavirales. Front. Microbiol. 2017, 8, 1514. [Google Scholar] [CrossRef]
- Rott, M.E.; Kesanakurti, P.; Berwarth, C.; Rast, H.; Boyes, I.; Phelan, J.; Jelkmann, W. Discovery of Negative-Sense RNA Viruses in Trees Infected with Apple Rubbery Wood Disease by Next-Generation Sequencing. Plant Dis. 2017, 102, 1254–1263. [Google Scholar] [CrossRef]
- Falk, B.W.; Tsai, J.H. Biology and Molecular Biology of Viruses in the Genus Tenuivirus. Annu. Rev. Phytopathol. 1998, 36, 139–163. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P. An Overview on Grapevine Viruses, Viroids, and the Diseases They Cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 31–46. ISBN 978-3-319-57706-7. [Google Scholar]
- Chen, H.; Tzeng, D.; Chen, M. Preliminary studies on the grapevine yellow dwarf, a new virus disease in Taiwan. Natl. Sci. Counc. Mon. 1981, 9, 584–591. [Google Scholar]
- Stobbs, L.W.; Broadbent, A.B. Susceptibility of grapevine cultivars to Tomato spotted wilt virus in southern Ontario, Canada. Plant Dis. 1993, 77, 318. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Klaassen, V.; Stevens, K.; Sudarshana, M.R.; Rowhani, A.; Maree, H.J.; Chooi, K.M.; Blouin, A.G.; Habili, N.; Song, Y.; et al. Characterization of grapevine leafroll-associated virus 3 genetic variants and application towards RT-qPCR assay design. PLOS ONE 2018, 13, e0208862. [Google Scholar] [CrossRef] [PubMed]
- Osman, F.; Leutenegger, C.; Golino, D.; Rowhani, A. Real-time RT-PCR (TaqMan®) assays for the detection of Grapevine Leafroll associated viruses 1–5 and 9. J. Virol. Methods 2007, 141, 22–29. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Rowhani, A.; Westrick, N.; Stevens, K.; Diaz-Lara, A.; Trouillas, F.P.; Preece, J.; Kallsen, C.; Farrar, K.; Golino, D. Discovery of Viruses and Virus-Like Pathogens in Pistachio using High-Throughput Sequencing. Plant Dis. 2018, 102, 1419–1425. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Eddy, S.R. A new generation of homology search tools based on probabilistic inference. In Genome Informatics 2009; Imperial College Press: London, UK; World Scientific Publishing Co.: Yokohama, Japan, 2009; pp. 205–211. ISBN 978-1-84816-562-5. [Google Scholar]
- Bateman, A.; Coin, L.; Durbin, R.; Finn, R.D.; Hollich, V.; Griffiths-Jones, S.; Khanna, A.; Marshall, M.; Moxon, S.; Sonnhammer, E.L.L.; et al. The Pfam protein families database. Nucleic Acids Res. 2004, 32, D138–D141. [Google Scholar] [CrossRef]
- Pei, J.; Kim, B.-H.; Grishin, N.V. PROMALS3D: A tool for multiple protein sequence and structure alignments. Nucleic Acids Res. 2008, 36, 2295–2300. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformation 2007, 23, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformation 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformation 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Aradhya, M.K.; Preece, J.; Kluepfel, D.A. Genetic conservation, characterization and utilization of wild relatives of fruit and nut crops at the USDA germplasm repository in Davis, California. Acta Hortic. 2015, 1074, 95–104. [Google Scholar] [CrossRef]
- Puckett, J.; Al Rwahnih, M.; Klassen, V.; Golino, D. The Davis Grapevine Virus Collection—A Current Perspective. In Proceedings of the 19th Congress of the ICVG, Santiago, Chile, 9–12 April 2018; pp. 186–187. [Google Scholar]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Islas, C.; Rowhani, A. Comparison of Next-Generation Sequencing Versus Biological Indexing for the Optimal Detection of Viral Pathogens in Grapevine. Phytopathology 2015, 105, 758–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guu, T.S.; Zheng, W.; Tao, Y.J. Bunyavirus: Structure and replication. In Viral Molecular Machines; Springer: Boston, MA, USA, 2012; pp. 245–266. [Google Scholar]
- Reguera, J.; Weber, F.; Cusack, S. Bunyaviridae RNA Polymerases (L-Protein) Have an N-Terminal, Influenza-Like Endonuclease Domain, Essential for Viral Cap-Dependent Transcription. PLOS Pathog. 2010, 6, e1001101. [Google Scholar] [CrossRef] [PubMed]
- Melcher, U. The ‘30K’ superfamily of viral movement proteins. J. Gen. Virol. 2000, 81, 257–266. [Google Scholar] [CrossRef]
- Ferron, F.; Li, Z.; Danek, E.I.; Luo, D.; Wong, Y.; Coutard, B.; Lantez, V.; Charrel, R.; Canard, B.; Walz, T.; et al. The Hexamer Structure of the Rift Valley Fever Virus Nucleoprotein Suggests a Mechanism for its Assembly into Ribonucleoprotein Complexes. PLOS Pathog. 2011, 7, e1002030. [Google Scholar] [CrossRef]
- Katz, A.; Freiberg, A.N.; Backström, V.; Schulz, A.R.; Mateos, A.; Holm, L.; Pettersson, R.F.; Vaheri, A.; Flick, R.; Plyusnin, A. Oligomerization of Uukuniemi virus nucleocapsid protein. Virol. J. 2010, 7, 187. [Google Scholar] [CrossRef]
- Lu, G.; Li, J.; Zhou, Y.; Zhou, X.; Tao, X. Model-based structural and functional characterization of the Rice stripe tenuivirus nucleocapsid protein interacting with viral genomic RNA. Virology 2017, 506, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Mottram, T.J.; Li, P.; Dietrich, I.; Shi, X.; Brennan, B.; Varjak, M.; Kohl, A. Mutational analysis of Rift Valley fever phlebovirus nucleocapsid protein indicates novel conserved, functional amino acids. PLOS Negl. Trop. Dis. 2017, 11, e0006155. [Google Scholar] [CrossRef] [PubMed]
- Raymond, D.D.; Piper, M.E.; Gerrard, S.R.; Skiniotis, G.; Smith, J.L. Phleboviruses encapsidate their genomes by sequestering RNA bases. PNAS 2012, 109, 19208–19213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.-X.; Shi, M.; Tian, J.-H.; Lin, X.-D.; Kang, Y.-J.; Chen, L.-J.; Qin, X.-C.; Xu, J.; Holmes, E.C.; Zhang, Y.-Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. elife 2015, 4, e05378. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, É.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: Update 2019. Arch. Virol. 2019, 164, 1949–1965. [Google Scholar] [CrossRef] [PubMed]
- Maes, P.; Alkhovsky, S.V.; Bào, Y.; Beer, M.; Birkhead, M.; Briese, T.; Buchmeier, M.J.; Calisher, C.H.; Charrel, R.N.; Choi, I.R.; et al. Taxonomy of the family Arenaviridae and the order Bunyavirales: Update 2018. Arch. Virol. 2018, 163, 2295–2310. [Google Scholar] [CrossRef]
- Dolja, V.V.; Koonin, E.V. Common origins and host-dependent diversity of plant and animal viromes. Curr. Opin. Virol. 2011, 1, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Dolja, V.V.; Krupovic, M. Origins and evolution of viruses of eukaryotes: The ultimate modularity. Virology 2015, 479, 2–25. [Google Scholar] [CrossRef]
- Tokarz, R.; Sameroff, S.; Tagliafierro, T.; Jain, K.; Williams, S.H.; Cucura, D.M.; Rochlin, I.; Monzon, J.; Carpi, G.; Tufts, D.; et al. Identification of Novel Viruses in Amblyomma americanum, Dermacentor variabilis, and Ixodes scapularis Ticks. mSphere 2018, 3, e00614-17. [Google Scholar] [CrossRef]
- Lot, H.; Campbell, R.N.; Souche, S.; Milne, R.G.; Roggero, P. Transmission by Olpidium brassicae of Mirafiori lettuce virus and Lettuce big-vein virus, and Their Roles in Lettuce Big-Vein Etiology. Phytopathology 2002, 92, 288–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csorba, T.; Kontra, L.; Burgyán, J. viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Velasco, L.; Arjona-Girona, I.; Cretazzo, E.; López-Herrera, C. Viromes in Xylariaceae fungi infecting avocado in Spain. Virology 2019, 532, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-H.; Fujita, M.; Chiba, S.; Hyodo, K.; Andika, I.B.; Suzuki, N.; Kondo, H. Two novel fungal negative-strand RNA viruses related to mymonaviruses and phenuiviruses in the shiitake mushroom (Lentinula edodes). Virology 2019, 533, 125–136. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus * | BLASTp Results | Pairwise Identity (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RdRp | MP | NP | RdRp | RdRp Core | MP | NP | ||||||||
| E Value | Identities | E Value | Identities | E value | Identities | |||||||||
| GGDV | Apple rubbery wood virus 2 | 0.0 | 1610/2384 (68%) | 0.0 | 320/389 (82%) | 5.00E-175 | 258/288 (90%) | 67.24 | 78.57 | 67.10 | 75.87 | |||
| Apple rubbery wood virus 1 | 0.0 | 1383/2380 (58%) | 8.00E-162 | 241/385 (63%) | 5.00E-139 | 196/284 (69%) | 58.00 | 74.39 | 60.68 | 68.18 | ||||
| Grapevine Muscat rose virus | 0.0 | 1411/2660 (53%) | 2.00E-172 | 226/387 (58%) | 6.00E-156 | 211/284 (74%) | 53.48 | 70.41 | 59.52 | 74.30 | ||||
| GMRV | Apple rubbery wood virus 2 | 0.0 | 1337/2373 (56%) | 6.00E-166 | 228/375 (61%) | 6.00E-142 | 210/284 (74%) | 55.83 | 69.94 | 59.20 | 73.94 | |||
| Apple rubbery wood virus 1 | 0.0 | 1306/2387 (55%) | 8.00E-155 | 226/390 (58%) | 3.00E-142 | 194/284 (68%) | 54.60 | 70.61 | 57.33 | 67.96 | ||||
| Grapevine Garan dmak virus | 0.0 | 1411/2660 (53%) | 2.00E-172 | 226/387 (58%) | 6.00E-156 | 211/284 (74%) | 53.48 | 70.41 | 59.52 | 74.30 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diaz-Lara, A.; Navarro, B.; Di Serio, F.; Stevens, K.; Hwang, M.S.; Kohl, J.; Vu, S.T.; Falk, B.W.; Golino, D.; Al Rwahnih, M. Two Novel Negative-Sense RNA Viruses Infecting Grapevine Are Members of a Newly Proposed Genus within the Family Phenuiviridae. Viruses 2019, 11, 685. https://doi.org/10.3390/v11080685
Diaz-Lara A, Navarro B, Di Serio F, Stevens K, Hwang MS, Kohl J, Vu ST, Falk BW, Golino D, Al Rwahnih M. Two Novel Negative-Sense RNA Viruses Infecting Grapevine Are Members of a Newly Proposed Genus within the Family Phenuiviridae. Viruses. 2019; 11(8):685. https://doi.org/10.3390/v11080685
Chicago/Turabian StyleDiaz-Lara, Alfredo, Beatriz Navarro, Francesco Di Serio, Kristian Stevens, Min Sook Hwang, Joshua Kohl, Sandra Thuy Vu, Bryce W. Falk, Deborah Golino, and Maher Al Rwahnih. 2019. "Two Novel Negative-Sense RNA Viruses Infecting Grapevine Are Members of a Newly Proposed Genus within the Family Phenuiviridae" Viruses 11, no. 8: 685. https://doi.org/10.3390/v11080685
APA StyleDiaz-Lara, A., Navarro, B., Di Serio, F., Stevens, K., Hwang, M. S., Kohl, J., Vu, S. T., Falk, B. W., Golino, D., & Al Rwahnih, M. (2019). Two Novel Negative-Sense RNA Viruses Infecting Grapevine Are Members of a Newly Proposed Genus within the Family Phenuiviridae. Viruses, 11(8), 685. https://doi.org/10.3390/v11080685