Identification, Genetic Analysis, and Pathogenicity of Classical Swine H1N1 and Human-Swine Reassortant H1N1 Influenza Viruses from Pigs in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses Isolation and Identification
2.2. Sequencing and Phylogenetic Analysis
2.3. Mouse Experiments
2.4. Histopathology
2.5. Statistical Analysis
2.6. Ethics Statement
3. Results
3.1. Virus Isolation and Identification
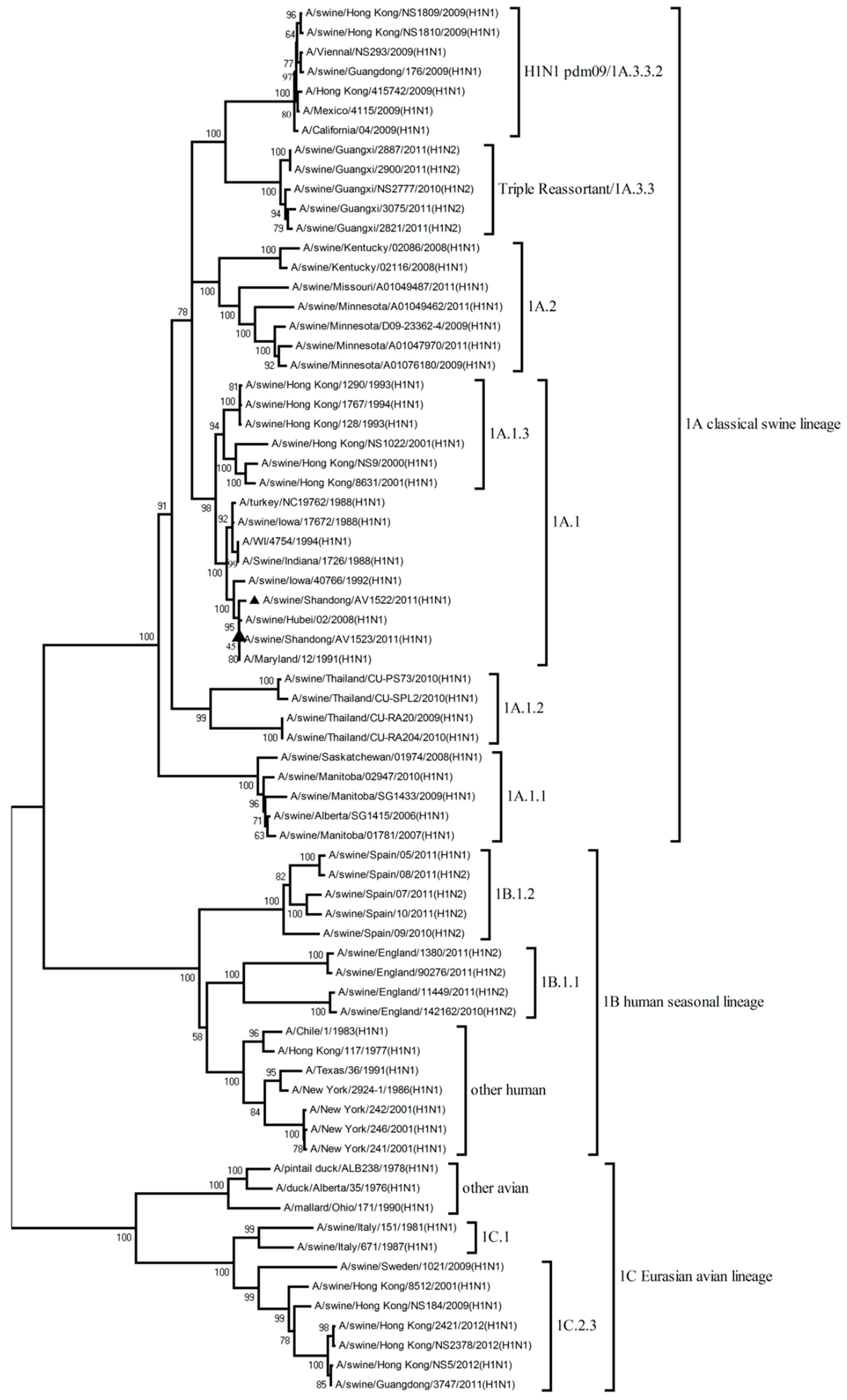
3.2. Phylogenetic Analysis of H1N1 Viruses
3.3. Molecular Characterization
3.4. Pathogenicity of the Two H1N1 Viruses in BALB/c Mice
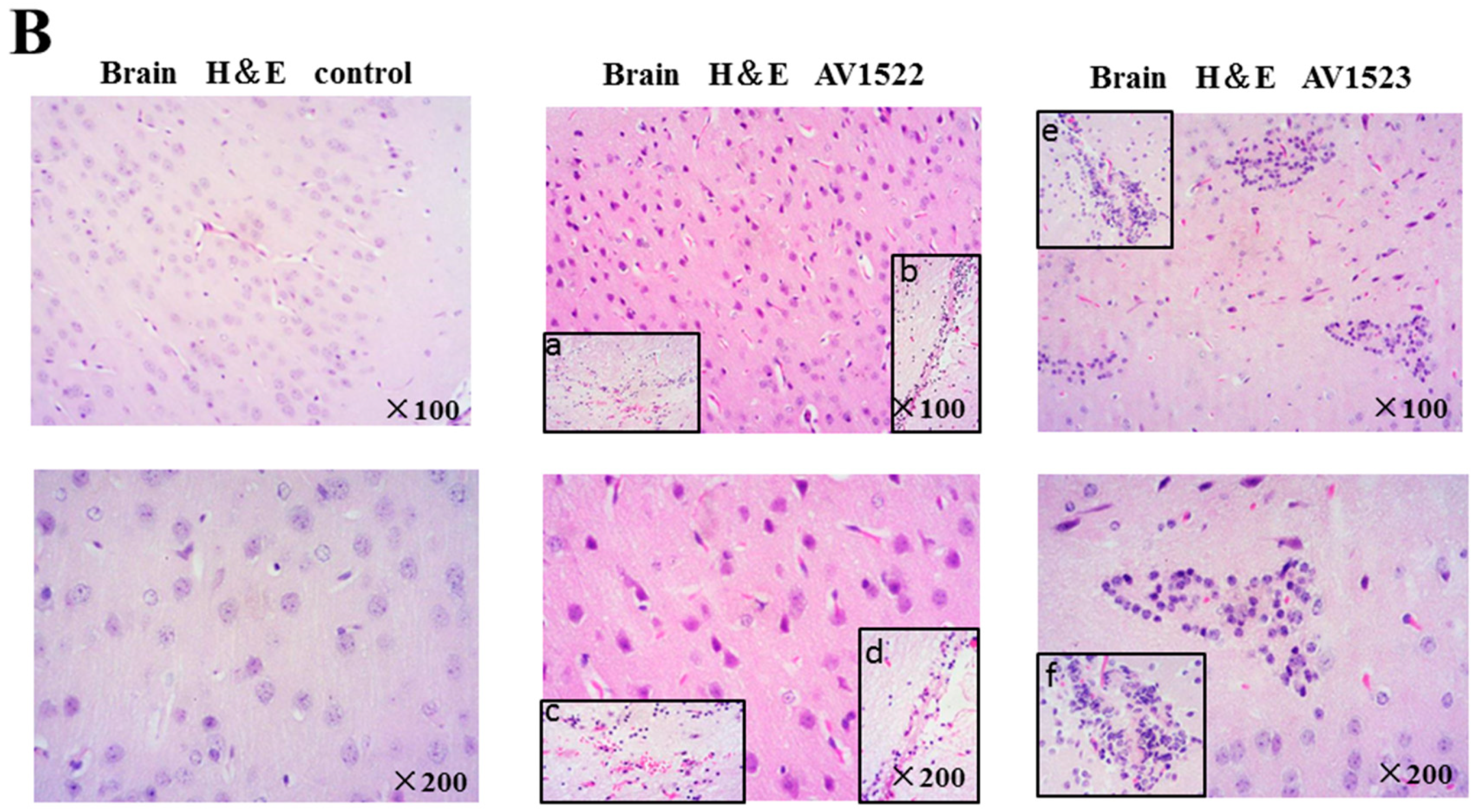
3.5. Histopathological Damage to Mouse Lungs and Brains Caused by AV1522 and AV1523 Viruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alexander, D.J. An overview of the epidemiology of avian influenza. Vaccine 2007, 25, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- Petrova, V.N.; Russell, C.A. The evolution of seasonal influenza viruses. Nat. Rev. Microbiol. 2018, 16, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudon, E.; Peyre, M.; Peiris, M.; Cowling, B.J. Epidemiological features of influenza circulation in swine populations: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0179044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, I.H. History and epidemiology of Swine influenza in Europe. Curr. Top. Microbiol. Immunol. 2013, 370, 133–146. [Google Scholar] [PubMed]
- Karasin, A.I.; Landgraf, J.; Swenson, S.; Erickson, G.; Goyal, S.; Woodruff, M.; Scherba, G.; Anderson, G.; Olsen, C.W. Genetic characterization of H1N2 influenza A viruses isolated from pigs throughout the United States. J. Clin. Microbiol. 2002, 40, 1073–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karasin, A.I.; Schutten, M.M.; Cooper, L.A.; Smith, C.B.; Subbarao, K.; Anderson, G.A.; Carman, S.; Olsen, C.W. Genetic characterization of H3N2 influenza viruses isolated from pigs in North America, 1977–1999: Evidence for wholly human and reassortant virus genotypes. Virus Res. 2000, 68, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.L.; Ma, W.; Lager, K.M.; Gramer, M.R.; Richt, J.A.; Janke, B.H. Characterization of a newly emerged genetic cluster of H1N1 and H1N2 swine influenza virus in the United States. Virus Genes 2009, 39, 176–185. [Google Scholar] [CrossRef]
- Webby, R.J.; Rossow, K.; Erickson, G.; Sims, Y.; Webster, R. Multiple lineages of antigenically and genetically diverse influenza A virus co-circulate in the United States swine population. Virus Res. 2004, 103, 67–73. [Google Scholar] [CrossRef]
- Zhou, N.N.; Senne, D.A.; Landgraf, J.S.; Swenson, S.L.; Erickson, G.; Rossow, K.; Liu, L.; Yoon, K.; Krauss, S.; Webster, R.G. Genetic reassortment of avian, swine, and human influenza A viruses in American pigs. J. Virol. 1999, 73, 8851–8856. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Webby, R.; Lam, T.T.; Smith, D.K.; Peiris, J.S.; Guan, Y. History of Swine influenza viruses in Asia. Curr. Top. Microbiol. Immunol. 2013, 370, 57–68. [Google Scholar]
- Bouvier, N.M.; Palese, P. The biology of influenza viruses. Vaccine 2008, 26 (Suppl. 4), D49–D53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Brockwell-Staats, C.; Webster, R.G.; Webby, R.J. Diversity of influenza viruses in swine and the emergence of a novel human pandemic influenza A (H1N1). Influenza Other Respir Viruses 2009, 3, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Imai, M.; Kawaoka, Y. The role of receptor binding specificity in interspecies transmission of influenza viruses. Curr. Opin. Virol. 2012, 2, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Kahn, R.E.; Richt, J.A. The pig as a mixing vessel for influenza viruses: Human and veterinary implications. J. Mol. Genet. Med. 2008, 3, 158–166. [Google Scholar] [CrossRef]
- Peiris, J.S.; Poon, L.L.; Guan, Y. Emergence of a novel swine-origin influenza A virus (S-OIV) H1N1 virus in humans. J. Clin. Virol. 2009, 45, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.J.; Vijaykrishna, D.; Bahl, J.; Lycett, S.J.; Worobey, M.; Pybus, O.G.; Ma, S.K.; Cheung, C.L.; Raghwani, J.; Bhatt, S.; et al. Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. Nature 2009, 459, 1122–1125. [Google Scholar] [CrossRef] [Green Version]
- Tsukamoto, K.; Ashizawa, H.; Nakanishi, K.; Kaji, N.; Suzuki, K.; Okamatsu, M.; Yamaguchi, S.; Mase, M. Subtyping of avian influenza viruses H1 to H15 on the basis of hemagglutinin genes by PCR assay and molecular determination of pathogenic potential. J. Clin. Microbiol. 2008, 46, 3048–3055. [Google Scholar] [CrossRef] [Green Version]
- Tsukamoto, K.; Ashizawa, T.; Nakanishi, K.; Kaji, N.; Suzuki, K.; Shishido, M.; Okamatsu, M.; Mase, M. Use of reverse transcriptase PCR to subtype N1 to N9 neuraminidase genes of avian influenza viruses. J. Clin. Microbiol. 2009, 47, 2301–2303. [Google Scholar] [CrossRef] [Green Version]
- Vijaykrishna, D.; Smith, G.J.; Pybus, O.G.; Zhu, H.; Bhatt, S.; Poon, L.L.; Riley, S.; Bahl, J.; Ma, S.K.; Cheung, C.L.; et al. Long-term evolution and transmission dynamics of swine influenza A virus. Nature 2011, 473, 519–522. [Google Scholar] [CrossRef]
- Caton, A.J.; Brownlee, G.G.; Yewdell, J.W.; Gerhard, W. The antigenic structure of the influenza virus A/PR/8/34 hemagglutinin (H1 subtype). Cell 1982, 31, 417–427. [Google Scholar] [CrossRef]
- Wiley, D.C.; Wilson, I.A.; Skehel, J.J. Structural identification of the antibody-binding sites of Hong Kong influenza haemagglutinin and their involvement in antigenic variation. Nature 1981, 289, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Galvin, H.D.; Haw, T.Y.; Nutsford, A.N.; Husain, M. Drug resistance in influenza A virus: The epidemiology and management. Infect. Drug Resist. 2017, 10, 121–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, R.X.; Liu, L.A.; Wei, D.Q. Structural and energetic analysis of drug inhibition of the influenza A M2 proton channel. Trends Pharmacol. Sci. 2013, 34, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Reid, A.H.; Lourens, R.M.; Wang, R.; Jin, G.; Fanning, T.G. Characterization of the 1918 influenza virus polymerase genes. Nature 2005, 437, 889–893. [Google Scholar] [CrossRef] [PubMed]
- Salomon, R.; Franks, J.; Govorkova, E.A.; Ilyushina, N.A.; Yen, H.L.; Hulse-Post, D.J.; Humberd, J.; Trichet, M.; Rehg, J.E.; Webby, R.J.; et al. The polymerase complex genes contribute to the high virulence of the human H5N1 influenza virus isolate A/Vietnam/1203/04. J. Exp. Med. 2006, 203, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhu, W.; Feng, Z.; Gao, R.; Guo, J.; Li, X.; Liu, J.; Wang, D.; Shu, Y. Substitution of D701N in the PB2 protein could enhance the viral replication and pathogenicity of Eurasian avian-like H1N1 swine influenza viruses. Emerg. Microbes Infect. 2018, 7, 75. [Google Scholar] [CrossRef]
- Seo, S.H.; Hoffmann, E.; Webster, R.G. Lethal H5N1 influenza viruses escape host anti-viral cytokine responses. Nat. Med. 2002, 8, 950–954. [Google Scholar] [CrossRef]
- Jiao, P.; Tian, G.; Li, Y.; Deng, G.; Jiang, Y.; Liu, C.; Liu, W.; Bu, Z.; Kawaoka, Y.; Chen, H. A single-amino-acid substitution in the NS1 protein changes the pathogenicity of H5N1 avian influenza viruses in mice. J. Virol. 2008, 82, 1146–1154. [Google Scholar] [CrossRef] [Green Version]
- Shope, R.E. Swine Influenza: Iii. Filtration Experiments and Etiology. J. Exp. Med. 1931, 54, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Koen, J.S. A practical method for field diagnosis of swine diseases. Am. J. Vet. Med. 1919, 14, 468–470. [Google Scholar]
- Chen, Y.; Zhang, J.; Qiao, C.; Yang, H.; Zhang, Y.; Xin, X.; Chen, H. Co-circulation of pandemic 2009 H1N1, classical swine H1N1 and avian-like swine H1N1 influenza viruses in pigs in China. Infect. Genet. Evol. 2013, 13, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.K.; Macken, C.A.; Lewis, N.S.; Scheuermann, R.H.; Van Reeth, K.; Brown, I.H.; Swenson, S.L.; Simon, G.; Saito, T.; Berhane, Y.; et al. A Phylogeny-Based Global Nomenclature System and Automated Annotation Tool for H1 Hemagglutinin Genes from Swine Influenza A Viruses. MSphere 2016, 1, e00275-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.; Lam, T.T.; Fan, X.; Chen, X.; Zeng, Y.; Zhou, J.; Duan, L.; Tse, M.; Chan, C.H.; Li, L.; et al. Expansion of genotypic diversity and establishment of 2009 H1N1 pandemic-origin internal genes in pigs in China. J. Virol. 2014, 88, 10864–10874. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Zhou, Y.J.; Li, G.X.; Zhang, G.H.; Liu, H.L.; Yan, L.P.; Liao, M.; Tong, G.Z. Further evidence for infection of pigs with human-like H1N1 influenza viruses in China. Virus Res. 2009, 140, 85–90. [Google Scholar] [CrossRef]
- Brown, I.H. The epidemiology and evolution of influenza viruses in pigs. Vet. Microbiol. 2000, 74, 29–46. [Google Scholar] [CrossRef]
- Karasin, A.I.; Carman, S.; Olsen, C.W. Identification of human H1N2 and human-swine reassortant H1N2 and H1N1 influenza A viruses among pigs in Ontario, Canada (2003 to 2005). J. Clin. Microbiol. 2006, 44, 1123–1126. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Yin, Y.; Sun, Z.; Gao, L.; Gao, G.F.; Liu, S.; Sun, L.; Liu, W. Genetic correlation between current circulating H1N1 swine and human influenza viruses. J. Clin. Virol. 2010, 49, 186–191. [Google Scholar] [CrossRef]
- Tumpey, T.M.; Basler, C.F.; Aguilar, P.V.; Zeng, H.; Solorzano, A.; Swayne, D.E.; Cox, N.J.; Katz, J.M.; Taubenberger, J.K.; Palese, P.; et al. Characterization of the reconstructed 1918 Spanish influenza pandemic virus. Science 2005, 310, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Pulit-Penaloza, J.A.; Pappas, C.; Belser, J.A.; Sun, X.; Brock, N.; Zeng, H.; Tumpey, T.M.; Maines, T.R. Comparative In Vitro and In Vivo Analysis of H1N1 and H1N2 Variant Influenza Viruses Isolated from Humans between 2011 and 2016. J. Virol. 2018, 92, e01444-18. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Shinya, K.; Kiso, M.; Watanabe, T.; Sakoda, Y.; Hatta, M.; Muramoto, Y.; Tamura, D.; Sakai-Tagawa, Y.; Noda, T.; et al. In vitro and in vivo characterization of new swine-origin H1N1 influenza viruses. Nature 2009, 460, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Wadford, D.A.; Pappas, C.; Gustin, K.M.; Maines, T.R.; Pearce, M.B.; Zeng, H.; Swayne, D.E.; Pantin-Jackwood, M.; Katz, J.M.; et al. Pathogenesis of pandemic influenza A (H1N1) and triple-reassortant swine influenza A (H1) viruses in mice. J. Virol. 2010, 84, 4194–4203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tumpey, T.M.; Garcia-Sastre, A.; Mikulasova, A.; Taubenberger, J.K.; Swayne, D.E.; Palese, P.; Basler, C.F. Existing antivirals are effective against influenza viruses with genes from the 1918 pandemic virus. Proc. Natl. Acad. Sci. USA 2002, 99, 13849–13854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memoli, M.J.; Tumpey, T.M.; Jagger, B.W.; Dugan, V.G.; Sheng, Z.M.; Qi, L.; Kash, J.C.; Taubenberger, J.K. An early ‘classical’ swine H1N1 influenza virus shows similar pathogenicity to the 1918 pandemic virus in ferrets and mice. Virology 2009, 393, 338–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Zhang, H.; Xiang, X.; Zhong, L.; Yang, L.; Guo, J.; Xie, Y.; Li, F.; Deng, Z.; Feng, H.; et al. Reassortant Eurasian Avian-Like Influenza A(H1N1) Virus from a Severely Ill Child, Hunan Province, China, 2015. Emerg. Infect. Dis. 2016, 22, 1930–1936. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.G. Increased virulence of a mouse-adapted variant of influenza A/FM/1/47 virus is controlled by mutations in genome segments 4, 5, 7, and 8. J. Virol. 1990, 64, 4523–4533. [Google Scholar]
- Kobasa, D.; Takada, A.; Shinya, K.; Hatta, M.; Halfmann, P.; Theriault, S.; Suzuki, H.; Nishimura, H.; Mitamura, K.; Sugaya, N.; et al. Enhanced virulence of influenza A viruses with the haemagglutinin of the 1918 pandemic virus. Nature 2004, 431, 703–707. [Google Scholar] [CrossRef]
- Pappas, C.; Aguilar, P.V.; Basler, C.F.; Solorzano, A.; Zeng, H.; Perrone, L.A.; Palese, P.; Garcia-Sastre, A.; Katz, J.M.; Tumpey, T.M. Single gene reassortants identify a critical role for PB1, HA, and NA in the high virulence of the 1918 pandemic influenza virus. Proc. Natl. Acad. Sci. USA 2008, 105, 3064–3069. [Google Scholar] [CrossRef] [Green Version]
- Uraki, R.; Kiso, M.; Shinya, K.; Goto, H.; Takano, R.; Iwatsuki-Horimoto, K.; Takahashi, K.; Daniels, R.S.; Hungnes, O.; Watanabe, T.; et al. Virulence determinants of pandemic A(H1N1)2009 influenza virus in a mouse model. J. Virol. 2013, 87, 2226–2233. [Google Scholar] [CrossRef] [Green Version]
- The Food and Agricultural Trade Dataset. Available online: http://www.fao.org/faostat/en/#data/TA (accessed on 9 January 2019).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Gene | Virus with Highest Similarity | Homology (%) |
|---|---|---|---|
| A/swine/Shandong/AV1522/2011(H1N1) | HA | A/Maryland/12/1991(H1N1) | 99.5 |
| NA | A/Maryland/12/1991(H1N1) | 100 | |
| PB2 | A/Maryland/12/1991(H1N1) | 99.8 | |
| PB1 | A/Maryland/12/1991(H1N1) | 100 | |
| PA | A/Maryland/12/1991(H1N1) | 99.8 | |
| NP | A/Maryland/12/1991(H1N1) | 99.7 | |
| M | A/Maryland/12/1991(H1N1) | 99.9 | |
| NS | A/Maryland/12/1991(H1N1) | 100 | |
| A/swine/Shandong/AV1523/2011(H1N1) | HA | A/Maryland/12/1991(H1N1) | 100 |
| NA | A/Alaska/1935(H1N1) | 99.4 | |
| PB2 | A/Alaska/1935(H1N1) | 100 | |
| PB1 | A/Alaska/1935(H1N1) | 100 | |
| PA | A/Victoria/36/1988(H1N1) | 99.9 | |
| NP | A/Ann Arbor/6/1960(H2N2) | 99.7 | |
| M | A/New Jersey/1976(H1N1) | 99.8 | |
| NS | A/Alaska/1935(H1N1) | 99.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Zhang, Y.; Zhang, B.; Chen, L.; Zhang, M.; Wang, J.; Jiang, Y.; Yang, C.; Jiang, T. Identification, Genetic Analysis, and Pathogenicity of Classical Swine H1N1 and Human-Swine Reassortant H1N1 Influenza Viruses from Pigs in China. Viruses 2020, 12, 55. https://doi.org/10.3390/v12010055
Song Y, Zhang Y, Zhang B, Chen L, Zhang M, Wang J, Jiang Y, Yang C, Jiang T. Identification, Genetic Analysis, and Pathogenicity of Classical Swine H1N1 and Human-Swine Reassortant H1N1 Influenza Viruses from Pigs in China. Viruses. 2020; 12(1):55. https://doi.org/10.3390/v12010055
Chicago/Turabian StyleSong, Yafen, Yong Zhang, Bing Zhang, Ling Chen, Min Zhang, Jingwen Wang, Ying Jiang, Chenghuai Yang, and Taozhen Jiang. 2020. "Identification, Genetic Analysis, and Pathogenicity of Classical Swine H1N1 and Human-Swine Reassortant H1N1 Influenza Viruses from Pigs in China" Viruses 12, no. 1: 55. https://doi.org/10.3390/v12010055
APA StyleSong, Y., Zhang, Y., Zhang, B., Chen, L., Zhang, M., Wang, J., Jiang, Y., Yang, C., & Jiang, T. (2020). Identification, Genetic Analysis, and Pathogenicity of Classical Swine H1N1 and Human-Swine Reassortant H1N1 Influenza Viruses from Pigs in China. Viruses, 12(1), 55. https://doi.org/10.3390/v12010055