Microtubule Retrograde Motors and Their Role in Retroviral Transport


Abstract
1. Introduction
2. Microtubules and Their Motor Proteins
3. Retrovirus Replication
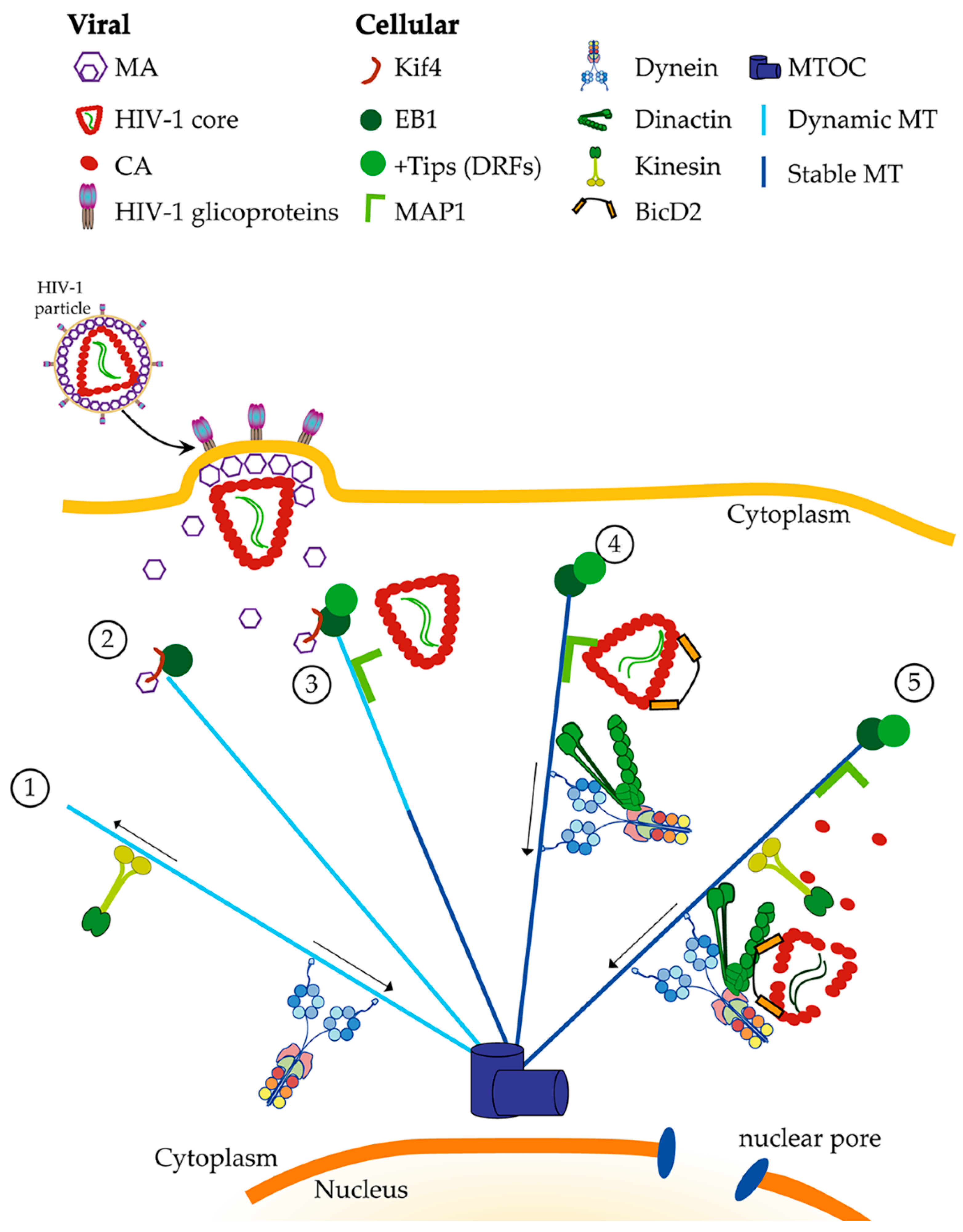
4. HIV-1 and Microtubule Associated Proteins at Early Infection
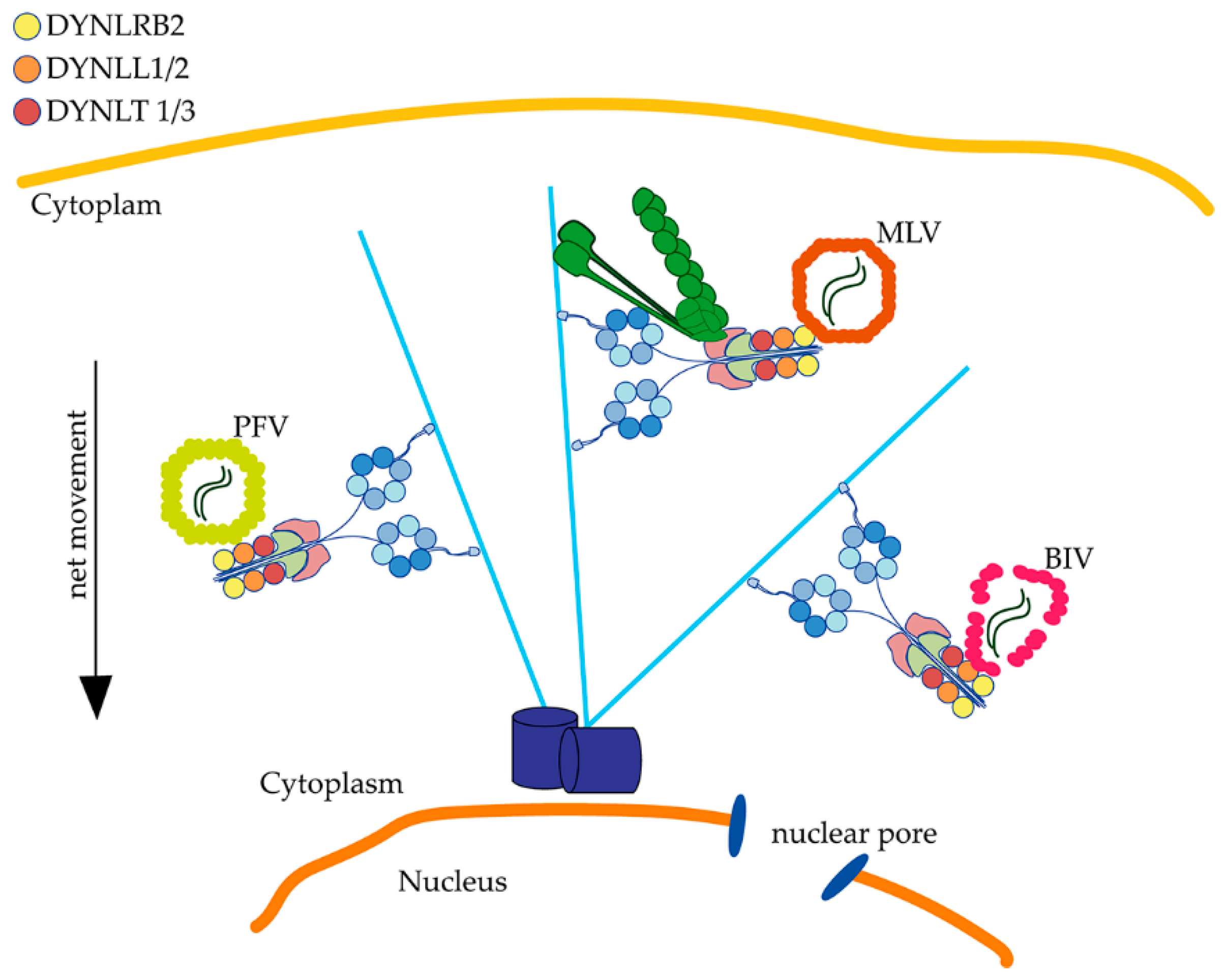
5. BIV, MLV and PFV directly Associate to Dynein Light Chains
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Merino-Gracia, J.; Garcia-Mayoral, M.F.; Rodriguez-Crespo, I. The association of viral proteins with host cell dynein components during virus infection. FEBS J. 2011, 278, 2997–3011. [Google Scholar] [CrossRef] [PubMed]
- Dodding, M.P.; Way, M. Coupling viruses to dynein and kinesin-1. EMBO J. 2011, 30, 3527–3539. [Google Scholar] [CrossRef] [PubMed]
- Leopold, P.L.; Pfister, K.K. Viral strategies for intracellular trafficking: Motors and microtubules. Traffic 2006, 7, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Lyman, M.G.; Enquist, L.W. Herpesvirus interactions with the host cytoskeleton. J. Virol. 2009, 83, 2058–2066. [Google Scholar] [CrossRef] [PubMed]
- Scherer, J.; Vallee, R.B. Adenovirus recruits dynein by an evolutionary novel mechanism involving direct binding to pH-primed hexon. Viruses 2011, 3, 1417–1431. [Google Scholar] [CrossRef]
- Dohner, K.; Nagel, C.H.; Sodeik, B. Viral stop-and-go along microtubules: Taking a ride with dynein and kinesins. Trends Microbiol. 2005, 13, 320–327. [Google Scholar] [CrossRef]
- Roberts, A.J. Emerging mechanisms of dynein transport in the cytoplasm versus the cilium. Biochem. Soc. Trans. 2018, 46, 967–982. [Google Scholar] [CrossRef]
- Hirokawa, N.; Noda, Y.; Tanaka, Y.; Niwa, S. Kinesin superfamily motor proteins and intracellular transport. Nat. Rev. Mol. Cell Biol. 2009, 10, 682–696. [Google Scholar] [CrossRef]
- Verhey, K.J.; Hammond, J.W. Traffic control: Regulation of kinesin motors. Nat. Rev. Mol. Cell Biol. 2009, 10, 765–777. [Google Scholar] [CrossRef]
- Hook, P.; Vallee, R.B. The dynein family at a glance. J. Cell Sci. 2006, 119, 4369–4371. [Google Scholar] [CrossRef]
- Can, S.; Lacey, S.; Gur, M.; Carter, A.P.; Yildiz, A. Directionality of dynein is controlled by the angle and length of its stalk. Nature 2019, 566, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Pfister, K.K.; Fisher, E.M.; Gibbons, I.R.; Hays, T.S.; Holzbaur, E.L.; McIntosh, J.R.; Porter, M.E.; Schroer, T.A.; Vaughan, K.T.; Witman, G.B.; et al. Cytoplasmic dynein nomenclature. J. Cell Biol. 2005, 171, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Kardon, J.R.; Vale, R.D. Regulators of the cytoplasmic dynein motor. Nat. Rev. Mol. Cell Biol. 2009, 10, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Caviston, J.P.; Holzbaur, E.L. Microtubule motors at the intersection of trafficking and transport. Trends Cell Biol. 2006, 16, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Gyoeva, F.K.; Sarkisov, D.V.; Khodjakov, A.L.; Minin, A.A. The tetrameric molecule of conventional kinesin contains identical light chains. Biochemistry 2004, 43, 13525–13531. [Google Scholar] [CrossRef]
- Wozniak, M.J.; Allan, V.J. Cargo selection by specific kinesin light chain 1 isoforms. EMBO J. 2006, 25, 5457–5468. [Google Scholar] [CrossRef]
- St-Louis, M.C.; Cojocariu, M.; Archambault, D. The molecular biology of bovine immunodeficiency virus: A comparison with other lentiviruses. Anim. Health Res. Rev. 2004, 5, 125–143. [Google Scholar] [CrossRef]
- Petit, C.; Giron, M.L.; Tobaly-Tapiero, J.; Bittoun, P.; Real, E.; Jacob, Y.; Tordo, N.; de The, H.; Saib, A. Targeting of incoming retroviral Gag to the centrosome involves a direct interaction with the dynein light chain 8. J. Cell Sci. 2003, 116, 3433–3442. [Google Scholar] [CrossRef]
- Su, Y.; Qiao, W.; Guo, T.; Tan, J.; Li, Z.; Chen, Y.; Li, X.; Li, Y.; Zhou, J.; Chen, Q. Microtubule-dependent retrograde transport of bovine immunodeficiency virus. Cell Microbiol. 2010, 12, 1098–1107. [Google Scholar] [CrossRef]
- Valle-Tenney, R.; Opazo, T.; Cancino, J.; Goff, S.P.; Arriagada, G. Dynein Regulators Are Important for Ecotropic Murine Leukemia Virus Infection. J. Virol. 2016, 90, 6896–6905. [Google Scholar] [CrossRef]
- Opazo, T.; Garces, A.; Tapia, D.; Barraza, F.; Bravo, A.; Schwenke, T.; Cancino, J.; Arriagada, G. Functional Evidence of the Involvement of the Dynein Light Chain DYNLRB2 in Murine Leukemia Virus Infection. J. Virol. 2017, 91, e00129-17. [Google Scholar] [CrossRef] [PubMed]
- Elis, E.; Ehrlich, M.; Prizan-Ravid, A.; Laham-Karam, N.; Bacharach, E. p12 tethers the murine leukemia virus pre-integration complex to mitotic chromosomes. PLoS Pathog. 2012, 8, e1003103. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Vodicka, M.A.; Lucero, G.; Svitkina, T.M.; Borisy, G.G.; Emerman, M.; Hope, T.J. Visualization of the intracellular behavior of HIV in living cells. J. Cell Biol. 2002, 159, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Foster, H.E.; Rondelet, A.; Lacey, S.E.; Bahi-Buisson, N.; Bird, A.W.; Carter, A.P. Cryo-EM Reveals How Human Cytoplasmic Dynein Is Auto-inhibited and Activated. Cell 2017, 169, 1303–1314. [Google Scholar] [CrossRef]
- Urnavicius, L.; Lau, C.K.; Elshenawy, M.M.; Morales-Rios, E.; Motz, C.; Yildiz, A.; Carter, A.P. Cryo-EM shows how dynactin recruits two dyneins for faster movement. Nature 2018, 554, 202–206. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, M.E.; Cianfrocco, M.A.; Htet, Z.M.; Tran, P.T.; Reck-Peterson, S.L.; Leschziner, A.E. Lis1 Has Two Opposing Modes of Regulating Cytoplasmic Dynein. Cell 2017, 170, 1197–1208. [Google Scholar] [CrossRef]
- Reck-Peterson, S.L.; Redwine, W.B.; Vale, R.D.; Carter, A.P. The cytoplasmic dynein transport machinery and its many cargoes. Nat. Rev. Mol. Cell Biol. 2018, 19, 382–398. [Google Scholar] [CrossRef]
- Dharan, A.; Opp, S.; Abdel-Rahim, O.; Keceli, S.K.; Imam, S.; Diaz-Griffero, F.; Campbell, E.M. Bicaudal D2 facilitates the cytoplasmic trafficking and nuclear import of HIV-1 genomes during infection. Proc. Natl. Acad. Sci. USA 2017, 114, E10707–E10716. [Google Scholar] [CrossRef]
- Carnes, S.K.; Zhou, J.; Aiken, C. HIV-1 Engages a Dynein-Dynactin-BICD2 Complex. for Infection and Transport. to the Nucleus. J. Virol. 2018. [Google Scholar] [CrossRef]
- Walczak, C.E.; Gayek, S.; Ohi, R. Microtubule-depolymerizing kinesins. Annu. Rev. Cell Dev. Biol. 2013, 29, 417–441. [Google Scholar] [CrossRef]
- Malikov, V.; da Silva, E.S.; Jovasevic, V.; Bennett, G.; Vieira, D.A.d.A.; Schulte, B.; Diaz-Griffero, F.; Walsh, D.; Naghavi, M.H. HIV-1 capsids bind and exploit the kinesin-1 adaptor FEZ1 for inward movement to the nucleus. Nat. Commun. 2015, 6, 6660. [Google Scholar] [CrossRef] [PubMed]
- Malikov, V.; Naghavi, M.H. Localized Phosphorylation of a Kinesin-1 Adaptor by a Capsid-Associated Kinase Regulates HIV-1 Motility and Uncoating. Cell Rep. 2017, 20, 2792–2799. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lukic, Z.; Dharan, A.; Fricke, T.; Diaz-Griffero, F.; Campbell, E.M. HIV-1 uncoating is facilitated by dynein and kinesin 1. J. Virol. 2014, 88, 13613–13625. [Google Scholar] [CrossRef] [PubMed]
- Goff, S.P. (Ed.) Retroviridae: The Retroviruses and Their Replication, 4th ed.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2001; pp. 1871–1939. [Google Scholar]
- Pereira, L.E.; Clark, J.; Grznarova, P.; Wen, X.; LaCasse, R.; Ruml, T.; Spearman, P.; Hunter, E. Direct evidence for intracellular anterograde co-transport of M-PMV Gag and Env on microtubules. Virology 2014, 449, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Sfakianos, J.N.; Hunter, E. M-PMV capsid transport is mediated by Env/Gag interactions at the pericentriolar recycling endosome. Traffic 2003, 4, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Fontenot, D.R.; den Hollander, P.; Vela, E.M.; Newman, R.; Sastry, J.K.; Kumar, R. Dynein light chain 1 peptide inhibits human immunodeficiency virus infection in eukaryotic cells. Biochem. Biophys. Res. Commun. 2007, 363, 901–907. [Google Scholar] [CrossRef]
- Gallo, D.E.; Hope, T.J. Knockdown of MAP4 and DNAL1 produces a post-fusion and pre-nuclear translocation impairment in HIV-1 replication. Virology 2012, 422, 13–21. [Google Scholar] [CrossRef][Green Version]
- Arhel, N.; Genovesio, A.; Kim, K.A.; Miko, S.; Perret, E.; Olivo-Marin, J.C.; Shorte, S.; Charneau, P. Quantitative four-dimensional tracking of cytoplasmic and nuclear HIV-1 complexes. Nat. Methods 2006, 3, 817–824. [Google Scholar] [CrossRef]
- Huang, P.T.; Summers, B.J.; Xu, C.; Perilla, J.R.; Malikov, V.; Naghavi, M.H.; Xiong, Y. FEZ1 Is Recruited to a Conserved Cofactor Site on Capsid to Promote HIV-1 Trafficking. Cell Rep. 2019, 28, 2373–2385. [Google Scholar] [CrossRef]
- Sabo, Y.; Walsh, D.; Barry, D.S.; Tinaztepe, S.; de los Santos, K.; Goff, S.P.; Gundersen, G.G.; Naghavi, M.H. HIV-1 induces the formation of stable microtubules to enhance early infection. Cell Host Microbe. 2013, 14, 535–546. [Google Scholar] [CrossRef]
- Naghavi, M.H.; Walsh, D. Microtubule Regulation and Function during Virus Infection. J. Virol. 2017, 91, e00538-17. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.; Naghavi, M.H. Exploitation of Cytoskeletal Networks during Early Viral Infection. Trends Microbiol. 2019, 27, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Delaney, M.K.; Malikov, V.; Chai, Q.; Zhao, G.; Naghavi, M.H. Distinct functions of diaphanous-related formins regulate HIV-1 uncoating and transport. Proc. Natl. Acad. Sci. USA 2017, 114, E6932–E6941. [Google Scholar] [CrossRef] [PubMed]
- Halpain, S.; Dehmelt, L. The MAP1 family of microtubule-associated proteins. Genome Biol. 2006, 7, 224. [Google Scholar] [CrossRef]
- Atherton, J.; Houdusse, A.; Moores, C. MAPping out distribution routes for kinesin couriers. Biol. Cell 2013, 105, 465–487. [Google Scholar] [CrossRef]
- Fernandez, J.; Portilho, D.M.; Danckaert, A.; Munier, S.; Becker, A.; Roux, P.; Zambo, A.; Shorte, S.; Jacob, Y.; Vidalain, P.O.; et al. Microtubule-associated proteins 1 (MAP1) promote human immunodeficiency virus type I (HIV-1) intracytoplasmic routing to the nucleus. J. Biol. Chem. 2015, 290, 4631–4646. [Google Scholar] [CrossRef]
- Swan, A.; Nguyen, T.; Suter, B. Drosophila Lissencephaly-1 functions with Bic-D and dynein in oocyte determination and nuclear positioning. Nat. Cell Biol. 1999, 1, 444–449. [Google Scholar] [CrossRef]
- Bullock, S.L.; Ish-Horowicz, D. Conserved signals and machinery for RNA transport in Drosophila oogenesis and embryogenesis. Nature 2001, 414, 611–616. [Google Scholar] [CrossRef]
- Delanoue, R.; Davis, I. Dynein anchors its mRNA cargo after apical transport in the Drosophila blastoderm embryo. Cell 2005, 122, 97–106. [Google Scholar] [CrossRef]
- Hoogenraad, C.C.; Akhmanova, A.; Howell, S.A.; Dortland, B.R.; de Zeeuw, C.I.; Willemsen, R.; Visser, P.; Grosveld, F.; Galjart, N. Mammalian Golgi-associated Bicaudal-D2 functions in the dynein-dynactin pathway by interacting with these complexes. EMBO J. 2001, 20, 4041–4054. [Google Scholar] [CrossRef]
- Hoogenraad, C.C.; Wulf, P.; Schiefermeier, N.; Stepanova, T.; Galjart, N.; Small, J.V.; Grosveld, F.; de Zeeuw, C.I.; Akhmanova, A. Bicaudal D induces selective dynein-mediated microtubule minus end-directed transport. EMBO J. 2003, 22, 6004–6015. [Google Scholar] [CrossRef] [PubMed]
- Matanis, T.; Akhmanova, A.; Wulf, P.; del Nery, E.; Weide, T.; Stepanova, T.; Galjart, N.; Grosveld, F.; Goud, B.; de Zeeuw, C.I.; et al. Bicaudal-D regulates COPI-independent Golgi-ER transport by recruiting the dynein-dynactin motor complex. Nat. Cell Biol. 2002, 4, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Pare, C.; Suter, B. Subcellular localization of Bic-D::GFP is linked to an asymmetric oocyte nucleus. J. Cell Sci. 2000, 113, 2119–2127. [Google Scholar] [PubMed]
- Larsen, K.S.; Xu, J.; Cermelli, S.; Shu, Z.; Gross, S.P. BicaudalD actively regulates microtubule motor activity in lipid droplet transport. PLoS ONE 2008, 3, e3763. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Xu, M.; Huang, Q.; Gates, A.; Zhang, X.; Castle, J.; Stec, E.; Ferrer, M.; Strulovici, B.; Hazuda, D. Genome-scale RNAi screen for host factors required for HIV replication. Cell Host Microbe. 2008, 4, 495–504. [Google Scholar] [CrossRef]
- Brass, A.L.; Dykxhoorn, D.M.; Benita, Y.; Yan, N.; Engelman, A.; Xavier, R.J.; Lieberman, J.; Elledge, S.J. Identification of host proteins required for HIV infection through a functional genomic screen. Science 2008, 319, 921–926. [Google Scholar] [CrossRef]
- Konig, R.; Zhou, Y.; Elleder, D.; Diamond, T.L.; Bonamy, G.M.; Irelan, J.T.; Chiang, C.Y.; Tu, B.P.; de Jesus, P.D.; Lilley, C.E.; et al. Global analysis of host-pathogen interactions that regulate early-stage HIV-1 replication. Cell 2008, 135, 49–60. [Google Scholar] [CrossRef]
- Marquez, C.L.; Lau, D.; Walsh, J.; Shah, V.; McGuinness, C.; Wong, A.; Aggarwal, A.; Parker, M.W.; Jacques, D.A.; Turville, S.; et al. Kinetics of HIV-1 capsid uncoating revealed by single-molecule analysis. eLife 2018, 7, e34772. [Google Scholar] [CrossRef]
- Francis, A.C.; Melikyan, G.B. Single HIV-1 Imaging Reveals Progression of Infection through CA-Dependent Steps of Docking at the Nuclear Pore, Uncoating, and Nuclear Transport. Cell Host Microbe. 2018, 23, 536–548. [Google Scholar] [CrossRef]
- Ambrose, Z.; Aiken, C. HIV-1 uncoating: Connection to nuclear entry and regulation by host proteins. Virology 2014, 454–455, 371–379. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Structure | Cellular Function | Relevance for Viral Infection |
|---|---|---|
Dynein | - Participates in retrograde organelle transport inside the cell. - Plays crucial role regulating stages of cell cycle. - Requires other molecules to work optimally that also play a role in cargo binding NudEL/NudE, Lis1, bicaudal D2). | - Participates in virus retrograde transport, carrying viral PIC towards the nucleus [23]. - Interacts with HIV-1 by associating with BicD2 [28,29]. - Interacts directly with BIV capsid for retrograde transport [19]. |
Dynactin | - Cofactor that activates dynein mediates vesicle mobility. - Links dynein to various cargos. - Strengthens dynein binding to microtubules. | - Required for retroviral infection, possibly mediating association to dynein [20,29]. |
Kinesin | - Participates in anterograde transport within the cell. - Contains a kinesin “tail” where cargo binds and a motor domain responsible for its movement. | - Kinesin 4, 8 and 13 actively regulate microtubule dynamics, aiding in the process of infection [30]. - Kinesin-1 adaptor, FEZ1 has been shown to regulate early transport of viral particles by associating to HIV-1 capsid [31]. - Kinesin might also play a role in uncoating of HIV-1 thanks to FEZ1 and Kif5B [32,33]. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietrantoni, G.; Ibarra-Karmy, R.; Arriagada, G. Microtubule Retrograde Motors and Their Role in Retroviral Transport. Viruses 2020, 12, 483. https://doi.org/10.3390/v12040483
Pietrantoni G, Ibarra-Karmy R, Arriagada G. Microtubule Retrograde Motors and Their Role in Retroviral Transport. Viruses. 2020; 12(4):483. https://doi.org/10.3390/v12040483
Chicago/Turabian StylePietrantoni, Gianfranco, Rodrigo Ibarra-Karmy, and Gloria Arriagada. 2020. "Microtubule Retrograde Motors and Their Role in Retroviral Transport" Viruses 12, no. 4: 483. https://doi.org/10.3390/v12040483
APA StylePietrantoni, G., Ibarra-Karmy, R., & Arriagada, G. (2020). Microtubule Retrograde Motors and Their Role in Retroviral Transport. Viruses, 12(4), 483. https://doi.org/10.3390/v12040483