Metagenomics of Neotropical Single-Stranded DNA Viruses in Tomato Cultivars with and without the Ty-1 Gene

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tomato Leaf Samples and Confirmation of the Presence/Absence of the Ty-1 Gene/Locus in the Genome of the Tomato Samples by Employing a Cleaved Amplified Polymorphic Sequence (CAPS) Marker System
2.2. Viral Isolates and Preliminary Confirmation of the Presence of Begomoviruses in the Tomato Leaf Samples
2.3. Enrichment via Rolling Circle Amplification of Circular DNA Molecules on Each Individual Sample
2.4. Next-Generation Sequencing (NGS) of the Two Tomato DNA Pools and Analysis of the NGS-Derived Sequences
2.5. Design of a Collection of Viral Species-Specific PCR Primers for Detection in Individual Samples
2.6. Validation of NGS-Derived Information via PCR Assays with Virus-Specific Primers
2.7. Sanger Dideoxy Sequencing Validation of Virus-Specific PCR Amplicons
3. Results
3.1. NGS Detection of Previously Reported Begomovirus Species in the Two Pools of Samples (with and without the Ty-1 Gene)
3.2. NGS Detection of Putative Three Novel Begomovirus Species as Well as a New Alpha-Satellite Species and a Gemycircularvirus (Genomoviridae) in the Tomato Samples
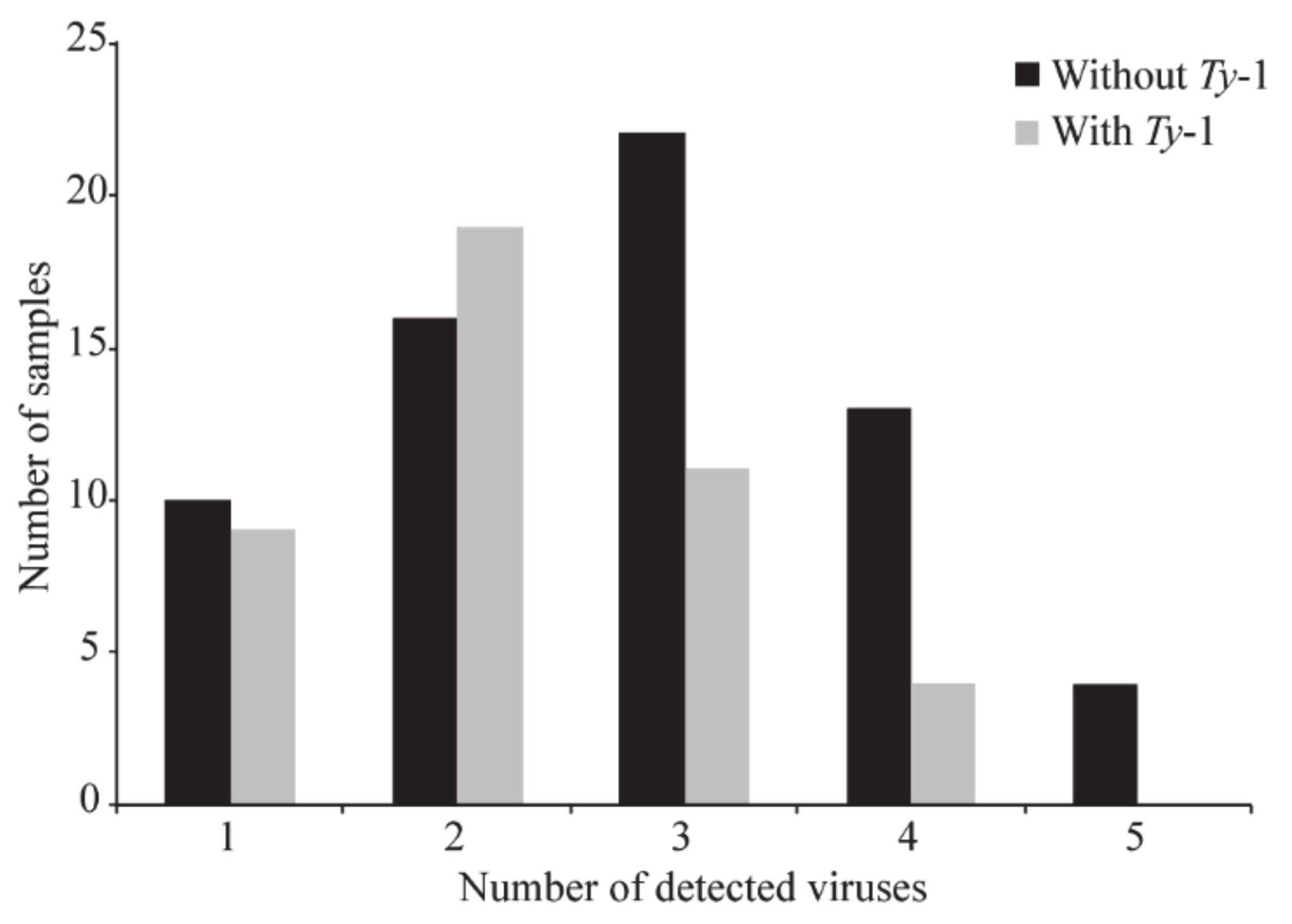
3.3. Confirmation via PCR Assays with Virus-Specific Primers and Sanger Dideoxy Sequencing of the Viral and Subviral ssDNA Species Present in each Individual Tomato Sample and Quantification of Mixed Infections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- ICTV International Committee on Taxonomy of Viruses. Available online: http://www.ictvonline.org (accessed on 6 January 2020).
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Fiallo-Olivé, E.; Briddon, R.W.; Hernández-Zepeda, C.; Idris, A. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef]
- Varsani, A.; Roumagnac, P.; Fuchs, M.; Navas-Castillo, J.; Moriones, E.; Idris, A.; Briddon, R.W.; Rivera-Bustamante, R.; Zerbini, F.M.; Martin, D.P. Capulavirus and Grablovirus: Two new genera in the family Geminiviridae. Arch. Virol. 2017, 162, 1819–1831. [Google Scholar] [CrossRef] [Green Version]
- Rojas, M.R.; Macedo, M.; Maliano, M.; Soto-Aguilar, M.; Souza, J.; Briddon, R.; Kenyon, L.; Rivera Bustamante, R.; Zerbini, F.; Adkins, S.; et al. World management of geminiviruses. Annu. Rev. Phytopathol. 2018, 56, 637–677. [Google Scholar] [CrossRef]
- De Barro, P.; Liu, S.-S.; Boykin, L.; Dinsdale, A. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Ribeiro, S.; Ambrozevicius, L.; Avila, A.; Bezerra, I.; Calegario, R.; Fernandes, J.; Lima, M.; De Mello, R.; Rocha, H.; Zerbini, F.; et al. Distribution and genetic diversity of tomato-infecting begomoviruses in Brazil. Arch. Virol. 2003, 148, 281–295. [Google Scholar] [CrossRef]
- Seal, S.; VandenBosch, F.; Jeger, M. Factors influencing begomovirus evolution and their increasing global significance: Implications for sustainable control. Crit. Rev. Plant Sci. 2006, 25, 23–46. [Google Scholar] [CrossRef]
- Silva, F.N.; Lima, A.; Rocha, C.; Castillo-Urquiza, G.; Alves-Júnior, M.; Zerbini, F. Recombination and pseudorecombination driving the evolution of the begomoviruses Tomato severe rugose virus (ToSRV) and Tomato rugose mosaic virus (ToRMV): Two recombinant DNA-A components sharing the same DNA-B. Virol. J. 2014, 11, 66. [Google Scholar] [CrossRef] [Green Version]
- FAO Food and Agriculture Organization of the United Nations (FAO). Available online: http://www.faostat.fao.org (accessed on 8 January 2020).
- Ribeiro, S.; Mello, L.; Boiteux, L.; Kitajima, E.; Faria, J. Tomato infection by a geminivirus in the Federal District, Brazil. Fitopatol. Bras. 1994, 19, 330. [Google Scholar]
- Rêgo-Machado, C.M.; Nakasu, E.Y.; Blawid, R.; Nagata, T.; Inoue-Nagata, A.K. Complete genome sequence of a new bipartite begomovirus infecting tomato in Brazil. Arch. Virol. 2019, 164, 2873–2875. [Google Scholar] [CrossRef] [PubMed]
- Calegario, R.; Ferreira, S.; Andrade, C.; Zerbini, F. Caracterização de um isolado do begomovírus Sida micrantha mosaic virus (SiMMV) obtido de tomateiro. Fitopatol. Bras. 2004, 29, 150. [Google Scholar]
- Cotrim, M.A.A.; Krause-Sakate, R.; Narita, N.; Zerbini, F.M.; Pavan, M.A. Diversidade genética de begomovírus em cultivos de tomateiro no Centro-Oeste Paulista. Summa Phytopathol. 2007, 33, 300–303. [Google Scholar] [CrossRef] [Green Version]
- Inoue-Nagata, A.K.; Lima, M.F.; Gilbertson, R.L. A review of geminivirus diseases in vegetables and other crops in Brazil: Current status and approaches for management. Hortic. Bras. 2016, 34, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.D.; Omoto, C.; Bleicher, E.; Dourado, P. Monitoring the susceptibility to insecticides in Bemisia tabaci (Gennadius)(Hemiptera: Aleyrodidae) populations from Brazil. Neotrop. Entomol. 2009, 38, 116–125. [Google Scholar]
- Yao, F.L.; Zheng, Y.; Huang, X.Y.; Ding, X.L.; Zhao, J.W.; Desneux, N.; He, Y.X.; Weng, Q.Y. Dynamics of Bemisia tabaci biotypes and insecticide resistance in Fujian province in China during 2005–2014. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zamir, D.; Ekstein-Michelson, I.; Zakay, Y.; Navot, N.; Zeidan, M.; Sarfatti, M.; Eshed, Y.; Harel, E.; Pleban, T.; Van-Oss, H.; et al. Mapping and introgression of a Tomato yellow leaf curl virus tolerance gene, Ty-1. Theor. Appl. Genet. 1994, 88, 141–146. [Google Scholar] [CrossRef]
- Hanson, P.; Green, S.; Kuo, G. Ty-2, a gene on chromosome 11 conditioning geminivirus resistance in tomato. Tomato Genet. Coop. Rep. 2006, 56, 17–18. [Google Scholar]
- Ji, Y.; Scott, J. Ty-3, a begomovirus resistance locus linked to Ty-1 on chromosome 6 of tomato. Tomato Genet. Coop. Rep. 2006, 56, 22–25. [Google Scholar]
- Ji, Y.; Scott, J.; Schuster, D. Molecular mapping of Ty-4, a new Tomato yellow leaf curl virus resistance locus on chromosome 3 of tomato. J. Am. Soc. Hortic. Sci. 2009, 134, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Anbinder, I.; Reuveni, M.; Azari, R.; Paran, I.; Nahon, S.; Shlomo, H.; Chen, L.; Lapidot, M.; Levin, I. Molecular dissection of Tomato leaf curl virus resistance in tomato line TY172 derived from Solanum peruvianum. Theor. Appl. Genet. 2009, 119, 519–530. [Google Scholar] [CrossRef]
- Hutton, S.; Scott, J. Ty-6, a major begomovirus resistance gene located on chromossome 10. Res. Rep. 2014, 64, 14–16. [Google Scholar]
- Giordano, L.; Silva-Lobo, V.; Santana, F.; Fonseca, M.; Boiteux, L. Inheritance of resistance to the bipartite Tomato chlorotic mottle begomovirus derived from Lycopersicon esculentum cv.‘Tyking’. Euphytica 2005, 143, 27–33. [Google Scholar] [CrossRef]
- Bian, X.Y.; Thomas, M.; Rasheed, M.; Saeed, M.; Hanson, P.; De Barro, P.; Rezaian, M. A recessive allele (tgr-1) conditioning tomato resistance to geminivirus infection is associated with impaired viral movement. Phytopathology 2007, 97, 930–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boiteux, L.; Fonseca, M.; Giordano, L.; Melo, P. Melhoramento genético. In Produção de Tomate para Processamento Industrial; Embrapa: Brasília-DF, Brazil, 2012; pp. 31–50. [Google Scholar]
- Boiteux, L.; Fonseca, M.; Viera, J.; Pereira-Carvalho, R. Melhoramento para resistência a doenças virais. In Melhoramento de Plantas para Condições de Estresses Bióticos; MG Suprema: Visconde de Rio Branco-MG, Brazil, 2012; pp. 89–119. [Google Scholar]
- Pereira-Carvalho, R.; Boiteux, L.; Fonseca, M.; Díaz-Pendón, J.; Moriones, E.; Fernández-Muñoz, R.; Charchar, J.; Resende, R. Multiple resistance to Meloidogyne spp. and to bipartite and monopartite Begomovirus spp. in wild Solanum (Lycopersicon) accessions. Plant Dis. 2010, 94, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verlaan, M.G.; Szinay, D.; Hutton, S.F.; de Jong, H.; Kormelink, R.; Visser, R.G.; Scott, J.W.; Bai, Y. Chromosomal rearrangements between tomato and Solanum chilense hamper mapping and breeding of the TYLCV resistance gene Ty-1. Plant J. 2011, 68, 1093–1103. [Google Scholar] [CrossRef]
- Maxwell, D.M.C.; Salus, M.; Montes, L.; Mejía, L. Tagging begomovirus resistance gene. Available online: www.plantpath.wisc.edu (accessed on 6 May 2020).
- Caro, M.; Verlaan, M.G.; Julián, O.; Finkers, R.; Wolters, A.-M.A.; Hutton, S.F.; Scott, J.W.; Kormelink, R.; Visser, R.G.; Díez, M.J. Assessing the genetic variation of Ty-1 and Ty-3 alleles conferring resistance to Tomato yellow leaf curl virus in a broad tomato germplasm. Mol. Breed. 2015, 35, 132. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Kim, H.J.; Lee, J.M.; Oh, C.S.; Lee, H.J.; Yeam, I. Gene-based molecular marker system for multiple disease resistances in tomato against Tomato yellow leaf curl virus, late blight, and verticillium wilt. Euphytica 2015, 205, 599–613. [Google Scholar] [CrossRef]
- Cooper, J.; Jones, A. Responses of plants to viruses: Proposals for the use of terms. Phytopathology 1983, 73, 127–128. [Google Scholar] [CrossRef] [Green Version]
- Boiteux, L.; Oliveira, V.; Silva, C.; Makishima, N.; Inoue-Nagata, A.; Fonseca, M.; Giordano, L. Reaction of tomato hybrids carrying the Ty-1 locus to Brazilian bipartite Begomovirus species. Hortic. Bras. 2007, 25, 20–23. [Google Scholar] [CrossRef]
- Voorburg, C.M.; Yan, Z.; Bergua-Vidal, M.; Wolters, A.M.A.; Bai, Y.; Kormelink, R. Ty-1, a universal resistance gene against geminiviruses that is compromised by co-replication of a betasatellite. Mol. Plant Pathol. 2020, 21, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Knief, C. Analysis of plant microbe interactions in the era of next generation sequencing technologies. Front. Plant Sci. 2014, 5, 216. [Google Scholar] [CrossRef] [Green Version]
- Adams, I.; Fox, A. Diagnosis of plant viruses using next-generation sequencing and metagenomic analysis. In Current Research Topics in Plant Virology; Springer: Cham, Switzerland, 2016; pp. 323–335. [Google Scholar]
- Hadidi, A. Next-Generation Sequencing and CRISPR/Cas13 editing in viroid research and molecular diagnostics. Viruses 2019, 11, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villamor, D.; Ho, T.; Al Rwahnih, M.; Martin, R.; Tzanetakis, I. High throughput sequencing for plant virus detection and discovery. Phytopathology 2019, 109, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Gao, S.; Hernandez, A.; Wechter, W.; Fei, Z.; Ling, K.S. Deep sequencing of small RNAs in tomato for virus and viroid identification and strain differentiation. PLoS ONE 2012, 7, e37127. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Sun, X.; Taylor, A.; Jiao, C.; Xu, Y.; Cai, X.; Wang, X.; Ge, C.; Pan, G.; Wang, Q. Diversity, distribution, and evolution of tomato viruses in China uncovered by small RNA sequencing. J. Virol. 2017, 91, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenele, R.S.; Lamas, N.S.; Lacorte, C.; Lacerda, A.L.M.; Varsani, A.; Ribeiro, S.G. A novel geminivirus identified in tomato and cleome plants sampled in Brazil. Virus Res. 2017, 240, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Negrete, E.A.; Morales-Aguilar, J.J.; Domínguez-Duran, G.; Torres-Devora, G.; Camacho-Beltrán, E.; Leyva-López, N.E.; Voloudakis, A.E.; Bejarano, E.R.; Méndez-Lozano, J. High-throughput sequencing reveals differential Begomovirus species diversity in non-cultivated plants in Northern-Pacific Mexico. Viruses 2019, 11, 594. [Google Scholar] [CrossRef] [Green Version]
- Boiteux, L.; Fonseca, M.; Simon, P. Effects of plant tissue and DNA purification method on randomly amplified polymorphic DNA-based genetic fingerprinting analysis in carrot. J. Am. Soc. Hortic. Sci. 1999, 124, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Rojas, M.R.; Gilbertson, R.; Russell, D.; Maxwell, D. Use of degenerate primers in the polymerase chain reaction to detect whitefly-transmitted geminiviruses. Plant Dis. 1993, 77, 340–347. [Google Scholar] [CrossRef]
- Ha, C.; Coombs, S.; Revill, P.; Harding, R.; Vu, M.; Dale, J. Corchorus yellow vein virus, a New World geminivirus from the Old World. J. Gen. Virol. 2006, 87, 997–1003. [Google Scholar] [CrossRef]
- Inoue-Nagata, A.K.; Albuquerque, L.; Rocha, W.; Nagata, T. A simple method for cloning the complete begomovirus genome using the bacteriophage φ29 DNA polymerase. J. Virol. Methods 2004, 116, 209–211. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Argüello-Astorga, G.; Ruiz-Medrano, R. An iteron-related domain is associated to Motif 1 in the replication proteins of geminiviruses: Identification of potential interacting amino acid-base pairs by a comparative approach. Arch. Virol. 2001, 146, 1465–1485. [Google Scholar] [CrossRef] [PubMed]
- Virus-HostDB Host Index. Available online: https://www.genome.jp/virushostdb/index/host/all (accessed on 10 February 2020).
- Inoue-Nagata, A.K.; Lopes, C.; Reis, A.; Pereira, R.; Quezao-Durval, A.; Pinheiro, J.; Lima, M. Doenças do Tomateiro. In Manual de Fitopatologia, 5th ed.; Amorim, L.B.F., Rezende, J.A.M., Camargo, L.E.A., Eds.; Agronômica Ceres: Sao Paulo-SP, Brazil, 2016; Volume 2, pp. 697–729. [Google Scholar]
- Rosen, R.; Kanakala, S.; Kliot, A.; Pakkianathan, B.C.; Farich, B.A.; Santana-Magal, N.; Elimelech, M.; Kontsedalov, S.; Lebedev, G.; Cilia, M. Persistent, circulative transmission of begomoviruses by whitefly vectors. Curr. Opin. Virol. 2015, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.K.; Verma, R.K.; Gaur, R.; Sanan-Mishra, N. Complexity and recombination analysis of novel begomovirus associated with spinach yellow vein disease in India. Plant Gene 2018, 13, 42–49. [Google Scholar] [CrossRef]
- Briddon, R.W.; Martin, D.P.; Roumagnac, P.; Navas-Castillo, J.; Fiallo-Olivé, E.; Moriones, E.; Lett, J.M.; Zerbini, F.M.; Varsani, A. Alphasatellitidae: A new family with two subfamilies for the classification of geminivirus-and nanovirus-associated alphasatellites. Arch. Virol. 2018, 163, 2587–2600. [Google Scholar] [CrossRef] [Green Version]
- Paprotka, T.; Metzler, V.; Jeske, H. The first DNA 1-like α satellites in association with New World begomoviruses in natural infections. Virology 2010, 404, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Romay, G.; Chirinos, D.; Geraud-Pouey, F.; Desbiez, C. Association of an atypical alphasatellite with a bipartite New World begomovirus. Arch. Virol. 2010, 155, 1843–1847. [Google Scholar] [CrossRef]
- Fontenele, R.S.; Lacorte, C.; Lamas, N.S.; Schmidlin, K.; Varsani, A.; Ribeiro, S.G. Single stranded DNA viruses associated with capybara faeces sampled in Brazil. Viruses 2019, 11, 710. [Google Scholar] [CrossRef] [Green Version]
- Male, M.F.; Kami, V.; Kraberger, S.; Varsani, A. Genome sequences of Poaceae-associated gemycircularviruses from the Pacific Ocean island of Tonga. Genome Announc. 2015, 3, e01144-15. [Google Scholar] [CrossRef] [Green Version]
- Marzano, S.Y.L.; Domier, L.L. Novel mycoviruses discovered from metatranscriptomics survey of soybean phyllosphere phytobiomes. Virus Res. 2016, 213, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Krupovic, M.; Ghabrial, S.A.; Jiang, D.; Varsani, A. Genomoviridae: A new family of widespread single-stranded DNA viruses. Arch. Virol. 2016, 161, 2633–2643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rezende, R.R.; Mar, T.B.; Páez, L.M.C.; da Silva Xavier, A.; Xavier, C.A.D.; Navas-Castillo, J.; Zerbini, F.M.; Alfenas-Zerbini, P. Complete genome sequences of two gemycircularviruses associated with non-cultivated plants in Brazil. Arch. Virol. 2018, 163, 3163–3166. [Google Scholar] [CrossRef]
- Lima, M.F.; Bezerra, I.; Ribeiro, S.; Ávila, A. Distribuição de geminivírus nas culturas do tomate e pimentão em doze municípios do Submédio do Vale São Francisco. Fitopatol. Bras. 2001, 26, 81–85. [Google Scholar] [CrossRef]
- Castillo-Urquiza, G.; Beserra, J.; Bruckner, F.; Lima, A.; Varsani, A.; Alfenas-Zerbini, P.; Zerbini, F. Six novel begomoviruses infecting tomato and associated weeds in Southeastern Brazil. Arch. Virol. 2008, 153, 1985–1989. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.C.; Albuquerque, L.; Rezende, J.; Inoue-Nagata, A.; Bergamin Filho, A.; Costa, H. Occurrence and molecular characterization of Tomato common mosaic virus (ToCmMV) in tomato fields in Espírito Santo state, Brazil. Trop. Plant Pathol. 2016, 41, 62–66. [Google Scholar] [CrossRef]
- Fernandes, F.R.; Albuquerque, L.C.; de Oliveira, C.L.; Cruz, A.R.; da Rocha, W.B.; Pereira, T.G.; Naito, F.Y.; Dias, N.D.M.; Nagata, T.; Faria, J.C.; et al. Molecular and biological characterization of a new Brazilian begomovirus, Euphorbia yellow mosaic virus (EuYMV), infecting Euphorbia heterophylla plants. Arch. Virol. 2011, 156, 2063. [Google Scholar] [CrossRef] [Green Version]
- Rezende, W.; Militão Neto, V.; Goulart, L.; GiovaniniI, M.; Juliatti, F.; Fernandes, J. Mixed infection by geminiviruses in tomato plants detected by LIS-SSCP-PCR. Fitopatol. Bras. 1997, 22, 338–339. [Google Scholar]
- Fernandes, F.R.; Albuquerque, L.; Giordano, L.; Boiteux, L.; Avila, A.; Inoue-Nagata, A. Diversity and prevalence of Brazilian bipartite Begomovirus species associated to tomatoes. Virus Genes 2008, 36, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Rocha, C.S.; Castillo-Urquiza, G.P.; Lima, A.T.; Silva, F.N.; Xavier, C.A.; Hora-Júnior, B.T.; Beserra-Júnior, J.E.; Malta, A.W.; Martin, D.P.; Varsani, A.; et al. Brazilian begomovirus populations are highly recombinant, rapidly evolving, and segregated based on geographical location. J. Virol. 2013, 87, 5784–5799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albuquerque, L.C.; Varsani, A.; Fernandes, F.; Pinheiro, B.; Martin, D.; Ferreira, P.; Lemos, T.; Inoue-Nagata, A. Further characterization of tomato-infecting begomoviruses in Brazil. Arch. Virol. 2012, 157, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.; Martin, D.; Lacorte, C.; Simões, I.; Orlandini, D.; Inoue-Nagata, A. Molecular and biological characterization of Tomato chlorotic mottle virus suggests that recombination underlies the evolution and diversity of Brazilian tomato begomoviruses. Phytopathology 2007, 97, 702–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.J.; Carvalho, M.; Andrade, E.; Brommonschenkel, S.; Fontes, E.; Zerbini, F. Biological and molecular properties of Tomato rugose mosaic virus (ToRMV), a new tomato-infecting begomovirus from Brazil. Plant Pathol. 2006, 55, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Belabess, Z.; Dallot, S.; El-Montaser, S.; Granier, M.; Majde, M.; Tahiri, A.; Blenzar, A.; Urbino, C.; Peterschmitt, M. Monitoring the dynamics of emergence of a non-canonical recombinant of Tomato yellow leaf curl virus and displacement of its parental viruses in tomato. Virology 2015, 486, 291–306. [Google Scholar] [CrossRef] [Green Version]
- Granier, M.; Tomassoli, L.; Manglli, A.; Nannini, M.; Peterschmitt, M.; Urbino, C. First report of TYLCV-IS141, a Tomato yellow leaf curl virus recombinant infecting tomato plants carrying the Ty-1 resistance gene in Sardinia (Italy). Plant Dis. 2019, 103, 1437. [Google Scholar] [CrossRef]
- Panno, S.; Caruso, A.G.; Davino, S. The nucleotide sequence of a recombinant tomato yellow leaf curl virus strain frequently detected in Sicily isolated from tomato plants carrying the Ty-1 resistance gene. Arch. Virol. 2018, 163, 795–797. [Google Scholar] [CrossRef]
- Torre, C.; Agüero, J.; Aranda, M. First evidence of Tomato yellow leaf curl virus-Israel IS76 recombinant isolates associated with severe yellow leaf curl epidemics in resistant tomatoes in Spain. Plant Dis. 2019, 103, 780. [Google Scholar] [CrossRef]
- Rêgo, C.D.M. Diversidade Genômica de Begomovírus em Tomateiros Resistente (BRS SENA) e Susceptível (H-9553); Universidade de Brasília: Brasilia-DF, Brazil, 2016. [Google Scholar]
- Ohnishi, J.; Yamaguchi, H.; Saito, A. Analysis of the mild strain of Tomato yellow leaf curl virus, which overcomes Ty-2 gene–mediated resistance in tomato line H24. Arch. Virol. 2016, 161, 2207–2217. [Google Scholar] [CrossRef]
- Tiwari, N.; Padmalatha, K.; Singh, V.; Haq, Q.; Malathi, V. Tomato leaf curl Bangalore virus (ToLCBV): Infectivity and enhanced pathogenicity with diverse betasatellites. Arch. Virol. 2010, 155, 1343–1347. [Google Scholar] [CrossRef]
- Butterbach, P.; Verlaan, M.G.; Dullemans, A.; Lohuis, D.; Visser, R.G.; Bai, Y.; Kormelink, R. Tomato yellow leaf curl virus resistance by Ty-1 involves increased cytosine methylation of viral genomes and is compromised by Cucumber mosaic virus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 12942–12947. [Google Scholar] [CrossRef] [Green Version]
- González, I.; Martínez, L.; Rakitina, D.V.; Lewsey, M.G.; Atencio, F.A.; Llave, C.; Kalinina, N.O.; Carr, J.P.; Palukaitis, P.; Canto, T. Cucumber mosaic virus 2b protein subcellular targets and interactions: Their significance to RNA silencing suppressor activity. Mol. Plant Microbe Interact. 2010, 23, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Hamera, S.; Song, X.; Su, L.; Chen, X.; Fang, R. Cucumber mosaic virus suppressor 2b binds to AGO4-related small RNAs and impairs AGO4 activities. Plant. J. 2012, 69, 104–115. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Geographic Region | Year of Collection | Isolate Code |
|---|---|---|
| Goiás State—GO | 2003 | GO-023, GO-046, GO-109, GO-111, GO-118, GO-120, GO-130, GO-134, GO-136, GO-137, GO-142, GO-143, GO-144, GO-168, GO-169, GO-191, GO-192, GO-221, GO-244, GO-245, GO-248, GO-249, GO-250, GO-251 |
| 2004 | GO-298, GO-299, GO-301, GO-322, GO-336 | |
| 2006 | GO-384, GO-390 | |
| 2011 | GO-493 | |
| 2012 | GO-505, GO-511 | |
| 2015 | GO-594 | |
| Federal District—DF | 2003 | DF-018, DF-023, DF-028, DF-043, DF-045, DF-046, DF-050, DF-062 |
| 2005 | DF-166, DF-167, DF-211 | |
| 2010 | DF-330 | |
| 2011 | DF-447, DF-453 | |
| 2013 | DF-544 | |
| 2014 | DF-566 | |
| 2016 | DF-667 | |
| Minas Gerais State—MG | 2001 | MG-046 |
| 2002 | MG-012, MG-015, MG-016, MG-018, MG-029 | |
| 2010 | MG-073, MG-113, MG-150 | |
| 2012 | MG-325 | |
| 2015 | MG-378, MG-388 |
| Geographic Region | Year of Collection | Isolate Code |
|---|---|---|
| Goiás State—GO | 2003 | GO-145, GO-148, GO-149, GO-151, GO-157, GO-161, GO-164 |
| 2004 | GO-247, GO-305, GO-307, GO-308, GO-320, GO-326, GO-330 | |
| 2007 | GO-371 | |
| 2010 | GO-479, GO-487, GO-490 | |
| 2013 | GO-550, GO-582, GO-583 | |
| Federal District—DF | 2007 | DF-227, DF-236, DF-238 |
| 2008 | DF-252 | |
| 2010 | DF-339 | |
| 2011 | DF-438 | |
| 2013 | DF-529, DF-550, DF-556 | |
| 2016 | DF-640 | |
| Minas Gerais State—MG | 2010 | MG-092, MG-122, MG-169, MG-282, MG-283, MG-284, MG-285, MG-286, MG-287 |
| 2012 | MG-326 | |
| 2015 | MG-383, MG-387 |
| Viral Species | Primer Name | Sequence 5′–3′ | Annealing Temperature (T °C) |
|---|---|---|---|
| Bean golden mosaic virus (BGMV) DNA-A | BGMV-For | GTGCGTGAATCCATGACCGT | 55 |
| BGMV-Rev | ATTCACGCACAGGGGAACG | ||
| Cleome leaf crumple virus (CILCrV) DNA-A | CILCrV-A-For | GACTCGACGTTCTGTGGT | 51 |
| CILCrV-A-Rev | TCCTAGTCGGGGCTCACT | ||
| Cleome leaf crumple virus (CILCrV) DNA-B | CILCrV-B-For | TAGGAAAGCAAAACGAGAATGGAA | 58 |
| CILCrV-B-Rev | GCTTTCCTAAATCGCAATTGATC | ||
| Tomato severe rugose virus (ToSRV) DNA-A | ToSRV-For5.1 | AGCGTCGTTAGCTGTCTGGCA | 58 |
| ToSRV-Rev5 | TGCCGCAGAAGCCTTGAACGCACCT | ||
| Tomato severe rugose virus (ToSRV) DNA-B | ToSRV-B-For | AAACCCACACGAAAGCAGAGTTT | 55 |
| ToSRV-B-Rev | CACCACGTCTATACATATTGTCCAGG | ||
| Euphorbia yellow mosaic virus (EuYMV) DNA-A | EuYMV-A-R-For | GGGGTTCCAAGTCCAATAAAGATGA | 52 |
| EuYMV-A-R-Rev | CAGACACCTTATATTTGCCGGATTC | ||
| Euphorbia yellow mosaic virus (EuYMV) DNA-B | EuYMV-B-R-For | GCCGAGGATAGAGGACACCAA | 60 |
| EuYMV-B-R-Rev | CCAGGCCCAAACGCATTATATTTTATC | ||
| Tomato chlorotic mottle virus (ToCMoV) DNA-A | ToCMoV-A-For | TTTGGGCCGCTCTTTTGGG | 47 |
| ToCMoV-A-Rev | CAAACTGAATGGGCCTTAAA | ||
| Tomato chlorotic mottle virus (ToCMoV) DNA-B | ToCMoV-B-For | GTATTTGTTCTGGGTGCAATCATAAAAC | 55 |
| ToCMoV-B-Rev | TTGTACTAATGACACATTATTCAATCACGA | ||
| Tomato golden vein virus (TGVV) DNA-A | TGVV-A-For1 | AAAGGAAGATAATTCAAATATAGGGA | 51 |
| TGVV-A-Rev1 | ATCTTCCTTTACTCACGTTCCTGAT | ||
| Tomato golden vein virus (TGVV) DNA-B | TGVV-B-S-For | CCCACTTTCCATAACCTACATGAGA | 55 |
| TGVV-B-S-For | GGAGAGAAAATTGATAAGATCGGCATC | ||
| Tomato mottle leaf curl virus (ToMLCV) DNA-A | ToMoLCV-For | TGTGGTCCAGTCAATAAATG | 47 |
| ToMoLCV-Rev | TGACTGGACCACATAGTAAA | ||
| Tomato common mosaic virus (ToCmMV) DNA-A | ToCmMV-For1 | ATTGCTCTCAACTTCTGTGC | 54 |
| ToCmMV-Rev2 | GCAATCCCTGGTGTCCTCAC | ||
| Tomato rugose mosaic virus (ToRMV) DNA-A | ToRMV-A-For | TGAAAGTAATTTTGACCCAATC | 52 |
| ToRMV-A-Rev | CAATTCATATGAGTTTTAGAGCAGC | ||
| Sida micrantha mosaic virus (SiMMV) DNA-A | SiMMV-For | GATCTCGCTCCCCCCTCT | 58 |
| SiMMV-Rev | AGATCGCACGACAACCAG | ||
| Plant-associated genomovirus 2 | Gemy-For | GCTCTGAATCAAATCTCGCTTACTTG | 54 |
| Gemy-Rev | CGATGTTGATTGGTTGGAAGCAAA | ||
| New Begomovirus Species #1 DNA-A | DF-640-A-For | GTTGACTGACATTTGCCTT | 47 |
| DF-640-A-Rev | TGTCAGTCAACAATCTATACACA | ||
| New Begomovirus Species #1 DNA-B | DF-640-B-For | GTTGTTTCAAGGGCGTCGAC | 55 |
| DF-640-B-Rev | CAACATCAGACATCCAGCAATAATAAACT | ||
| New Begomovirus Species #2 DNA-A | 1ToBYMV-A-For | ATCCATGTCCTCGGCAGTCT | 55 |
| 1ToBYMV-A-Rev | TCACGCACAGAGGAACGC | ||
| New Begomovirus Species #3 DNA-A | Abuti-A-For | GGACTCCAGGGGGCAAAA | 55 |
| Abuti-A-Rev | AGTCCCGTCCGTACCACTTG | ||
| Alpha-satellite | Alfa-For | TGGTGTCCTGGCTTATAT | 46 |
| Alfa-Rev | GGCGGAGTCCTTTTTTTT |
| Viral Species | No. of Reads | Size (nts) | GenBank Accession Numbers |
|---|---|---|---|
| Bean golden mosaic virus (BGMV) DNA-A | 63,525 | 2626 | MT214083 |
| Cleome leaf crumple virus (CILCrV) DNA-A | 566 | 2560 | MN337873 |
| Cleome leaf crumple virus (CILCrV) DNA-B | 702 | 2664 | MN337872 |
| Tomato severe rugose virus (ToSRV) DNA-A | 3,225,120 | 2593 | MT214084 |
| Tomato severe rugose virus (ToSRV) DNA-B | 4,018,351 | 2572 | MT214085 |
| Euphorbia yellow mosaic virus (EuYMV) DNA-A | 1122 | 2609 | MN746971 |
| Euphorbia yellow mosaic virus (EuYMV) DNA-B | 1822 | 2579 | MN746970 |
| Tomato chlorotic mottle virus (ToCMoV) DNA-A | 5,971,019 | 2620 | MT214086 |
| Tomato chlorotic mottle virus (ToCMoV) DNA-B | 1,111,227 | 2600 | MT214087 |
| Tomato golden vein virus (TGVV) DNA-A | 2,639,961 | 2562 | MN928610 |
| Tomato golden vein virus (TGVV) DNA-B | 977,027 | 2512 | MN928611 |
| Tomato mottle leaf curl virus (ToMLCV) DNA-A | 1,784,881 | 2632 | MT214088 |
| Tomato common mosaic virus (ToCmMV) DNA-A | 1,070,674 | 2560 | MT214089 |
| Tomato rugose mosaic virus (ToRMV) DNA-A | 3,267,808 | 2619 | MT214090 |
| Tomato rugose mosaic virus (ToRMV) DNA-B | 4,742,730 | 2571 | MT214091 |
| Sida micrantha mosaic virus (SiMMV) DNA-A | 1,221,062 | 2688 | MT214092 |
| Plant-associated genomovirus 2 | 119 | 2189 | MT214094 |
| New Begomovirus Species #2 DNA-A | 427,646 | 2649 | MT214095 |
| New Begomovirus Species #3 DNA-A | 2839 | 2636 | MT214096 |
| New alpha-satellite | 155,793 | 1321 | MT214093 |
| Viral Species | N° of Reads | Size (nts) | GenBank Accession Numbers |
|---|---|---|---|
| Tomato severe rugose virus (ToSRV) DNA-A | 7,181,771 | 2592 | MT215001 |
| Tomato severe rugose virus (ToSRV) DNA-B | 5,782,296 | 2570 | MT215002 |
| Tomato golden vein virus (TGVV) DNA-A | 2,358,838 | 2561 | MN928612 |
| Tomato golden vein virus (TGVV) DNA-B | 1,401,684 | 2590 | MN928613 |
| Tomato chlorotic mottle virus (ToCMoV) DNA-A | 4,519,040 | 2623 | MT215003 |
| Tomato chlorotic mottle virus (ToCMoV) DNA-B | 811,733 | 2565 | MT215004 |
| Tomato mottle leaf curl virus (ToMLCV) DNA-A | 2,644,606 | 2631 | MT215005 |
| Tomato rugose mosaic virus (ToRMV) DNA-A | 7,964,942 | 2618 | MT215006 |
| Tomato rugose mosaic virus (ToRMV) DNA-B | 5,780,864 | 2649 | MT215007 |
| New Begomovirus Species #1 DNA-A | 1,270,494 | 2605 | MN147863 |
| New Begomovirus Species #1 DNA-B | 84,022 | 2625 | MN147864 |
| Viral Species* Followed by the Respective Number of Occurrences in Each Region | Goiás State (GO) | Federal District (DF) | Minas Gerais State (MG) |
|---|---|---|---|
| ToSRV (32 + 9 + 5) = 46 | GO-046, GO-109, GO-118, GO-120, GO-130, GO-134, GO-136, GO-137, GO-142, GO-143, GO-144, GO-168, GO-169, GO-191, GO-192, GO-221, GO-244, GO-245, GO-248, GO-249, GO-250, GO-251, GO-298, GO-299, GO-301, GO-322, GO-336, GO-390, GO-493, GO-505, GO-511, GO-594 | DF-043, DF-166, DF-167, DF-211, DF-447, DF-453, DF-544, DF-566, DF-667 | MG-012, MG-018, MG-029, MG-150, MG-388 |
| TGVV (23 + 8 + 5) = 36 | GO-046, GO-109, GO-130, GO-134, GO-137, GO-142, GO-143, GO-168, GO-169, GO-191, GO-192, GO-221, GO-244, GO-245, GO-248, GO-249, GO-250, GO-298, GO-299, GO-301, GO-322, GO-336, GO-493 | DF-023, DF-028, DF-045, DF-046, DF-050, DF-062, DF-167, DF-211 | MG-015, MG-016, MG-018, MG-029, MG-046 |
| ToCMoV (21 + 8+5) = 34 | GO-023, GO-046, GO-109, GO-111, GO-120, GO-130, GO-134, GO-136, GO-137, GO-143, GO-144, GO-191, GO-245, GO-249, GO-250, GO-251, GO-298, GO-299, GO-301, GO-322, GO-390 | DF-018, DF-028, DF-043, DF-045, DF-046, DF-050, DF-167, DF-566 | MG-015, MG-018, MG-046, MG-073, MG-150 |
| ToCmMV (1 + 0 + 1) = 2 | GO-023 | --- | MG-388 |
| BGMV (1 + 2 + 0) = 3 | GO-142 | DF-045, DF-046 | --- |
| CILCrV (0 + 0 + 1) = 1 | --- | --- | MG-150 |
| EuYMV (0 + 0 + 2) = 2 | --- | --- | MG-012, MG-016 |
| ToMLCV (4 + 8 + 1) = 13 | GO-299, GO-384, GO-505, GO-594 | DF-018, DF-023, DF-028, DF-050, DF-062, DF-330, DF-453, DF-566 | MG-325. |
| ToRMV (19 + 1 + 1) = 21 | GO-109, GO-118, GO-130, GO-134, GO-136, GO-137, GO-143, GO-144, GO-168, GO-191, GO-192, GO-244, GO-248, GO-250, GO-251, GO-298, GO-322, GO-336, GO-505 | DF-043 | MG-150 |
| SiMMV (8 + 3 + 0) = 11 | GO-118, GO-120, GO-134, GO-168, GO-245, GO-248, GO-301, GO-511 | DF-045, DF-050, DF-166 | --- |
| Plant-associated genomovirus 2 (2 + 0 + 0) = 2 | GO-298, GO-301 | --- | --- |
| Alpha-satellite (0 + 4 + 0) = 4 | --- | DF-023, DF-028, DF-050, DF-062 | --- |
| New Begomovirus species #2 (0 + 0 + 1) = 1 | --- | --- | MG-378 |
| New Begomovirus species #3 (1 + 0 + 0) = 1 | GO-169 | --- | --- |
| Viral Species* Followed by the Respective Number of Occurrences in Each Region | Goiás State (GO) | Federal District (DF) | Minas Gerais State (MG) |
|---|---|---|---|
| ToSRV (14 + 5 + 7) = 26 | GO-145, GO-148, GO-151, GO-157, GO-161, GO-164, GO-247, GO-330, GO-371, GO-479, GO-487, GO-490, GO-550, GO-582 | DF-236, DF-339, DF-438, DF-550, DF-556 | MG-169, MG-285, MG-286, MG-287, MG-326, MG-383, MG-387 |
| TGVV (12 + 3 + 0) = 15 | GO-145, GO-148, GO-149, GO-151, GO-305, GO-320, GO-326, GO-371, GO-479, GO-490, GO-582, GO-583 | DF-236, DF-238, DF-438 | --- |
| ToCMoV (12 + 1 + 9) = 22 | GO-145, GO-148, GO-149, GO-305, GO-320, GO-326, GO-330, GO-371, GO-479, GO-490, GO-582, GO-583 | DF-550 | MG-092, MG-122, MG-169, MG-282, MG-283, MG-284, MG-285, MG-286, MG-287 |
| ToMLCV (4 + 8 + 1) = 13 | GO-307, GO-320, GO-326, GO-582 | DF-227, DF-236, DF-252, DF-339, DF-438, DF-529, DF-550, DF-556 | MG-326 |
| ToRMV (13 + 1 + 8) = 22 | GO-145, GO-148, GO-149, GO-151, GO-157, GO-161, GO-164, GO-247, GO-307, GO-308, GO-320, GO-330, GO-479 | DF-227 | MG-092, MG-122, MG-169, MG-282, MG-283, MG-284, MG-285, MG-286 |
| New Begomovirus species #1 (0 + 1 + 0) = 1 | --- | DF-640 | --- |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Nazaré Almeida dos Reis, L.; Fonseca, M.E.d.N.; Ribeiro, S.G.; Naito, F.Y.B.; Boiteux, L.S.; Pereira-Carvalho, R.d.C. Metagenomics of Neotropical Single-Stranded DNA Viruses in Tomato Cultivars with and without the Ty-1 Gene. Viruses 2020, 12, 819. https://doi.org/10.3390/v12080819
de Nazaré Almeida dos Reis L, Fonseca MEdN, Ribeiro SG, Naito FYB, Boiteux LS, Pereira-Carvalho RdC. Metagenomics of Neotropical Single-Stranded DNA Viruses in Tomato Cultivars with and without the Ty-1 Gene. Viruses. 2020; 12(8):819. https://doi.org/10.3390/v12080819
Chicago/Turabian Stylede Nazaré Almeida dos Reis, Luciane, Maria Esther de Noronha Fonseca, Simone Graça Ribeiro, Fernanda Yuri Borges Naito, Leonardo Silva Boiteux, and Rita de Cássia Pereira-Carvalho. 2020. "Metagenomics of Neotropical Single-Stranded DNA Viruses in Tomato Cultivars with and without the Ty-1 Gene" Viruses 12, no. 8: 819. https://doi.org/10.3390/v12080819
APA Stylede Nazaré Almeida dos Reis, L., Fonseca, M. E. d. N., Ribeiro, S. G., Naito, F. Y. B., Boiteux, L. S., & Pereira-Carvalho, R. d. C. (2020). Metagenomics of Neotropical Single-Stranded DNA Viruses in Tomato Cultivars with and without the Ty-1 Gene. Viruses, 12(8), 819. https://doi.org/10.3390/v12080819