
Potential of an Eco-Sustainable Probiotic-Cleaning Formulation in Reducing Infectivity of Enveloped Viruses

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Probiotic-Based Detergent
2.2. Viruses and Cells
2.3. Antiviral Activity: Suspension Tests
2.4. Antiviral Activity: Surface Tests
2.5. Analysis of Probiotic Enzymatic Activity
2.6. Statistical Analysis
3. Results
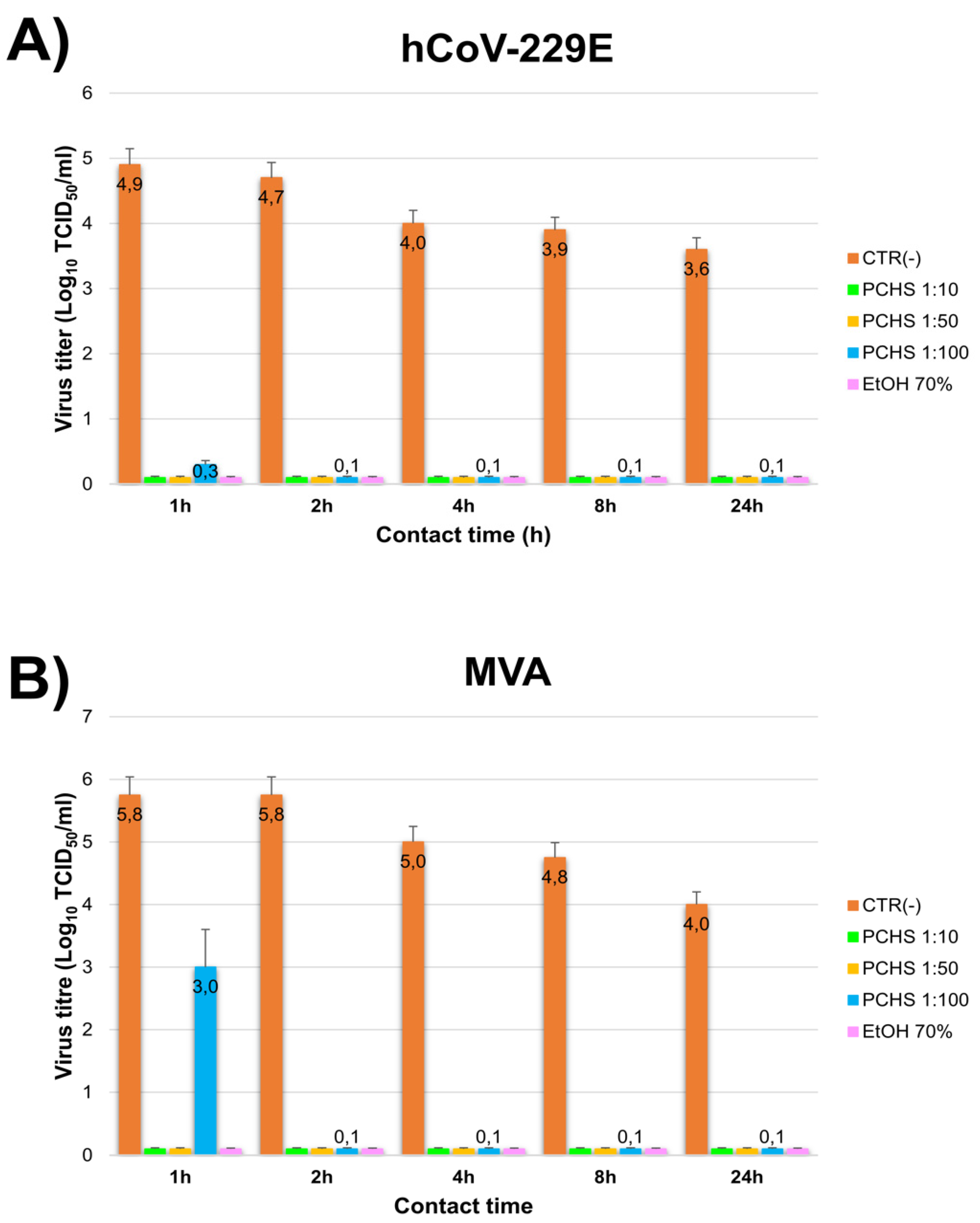
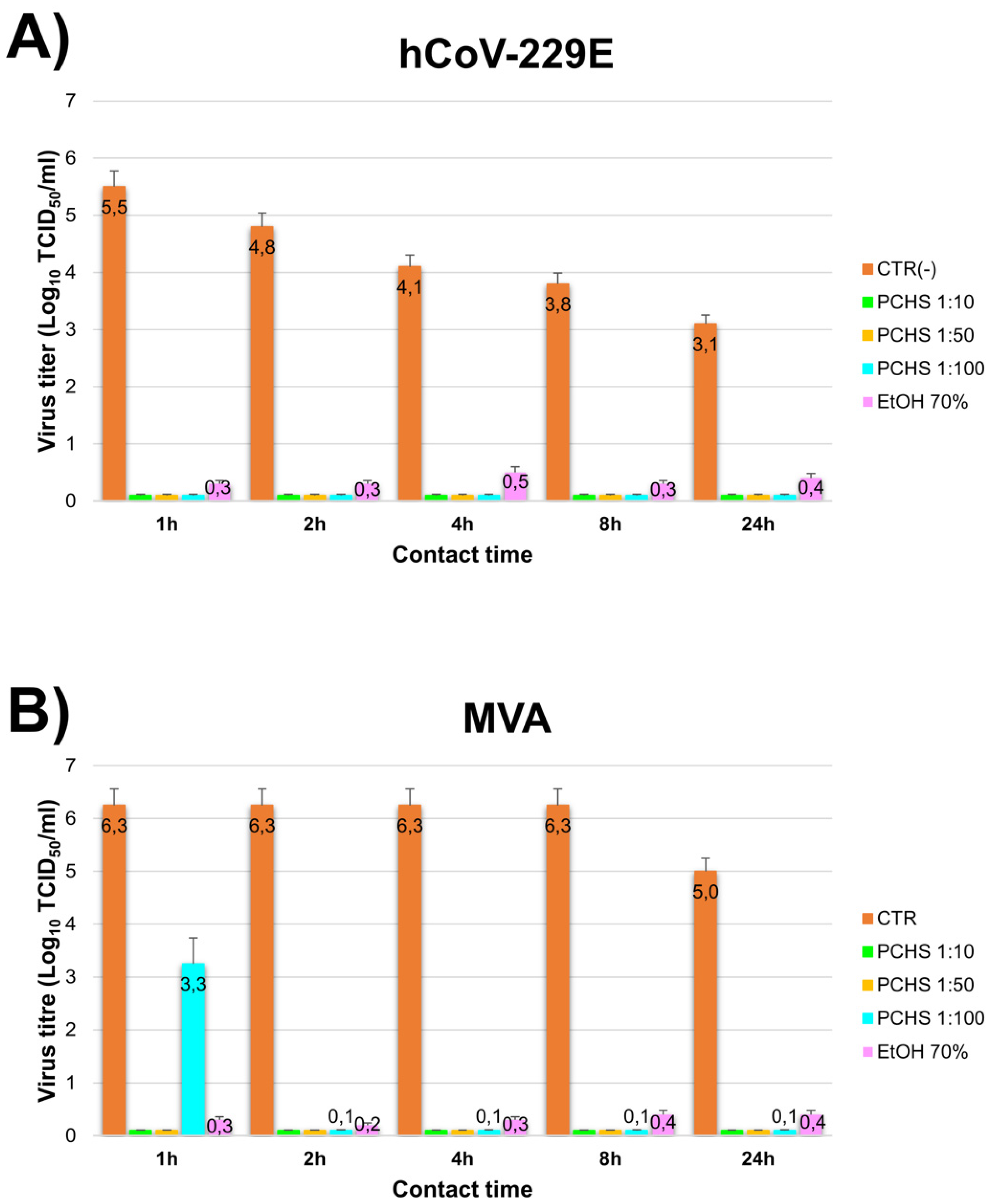
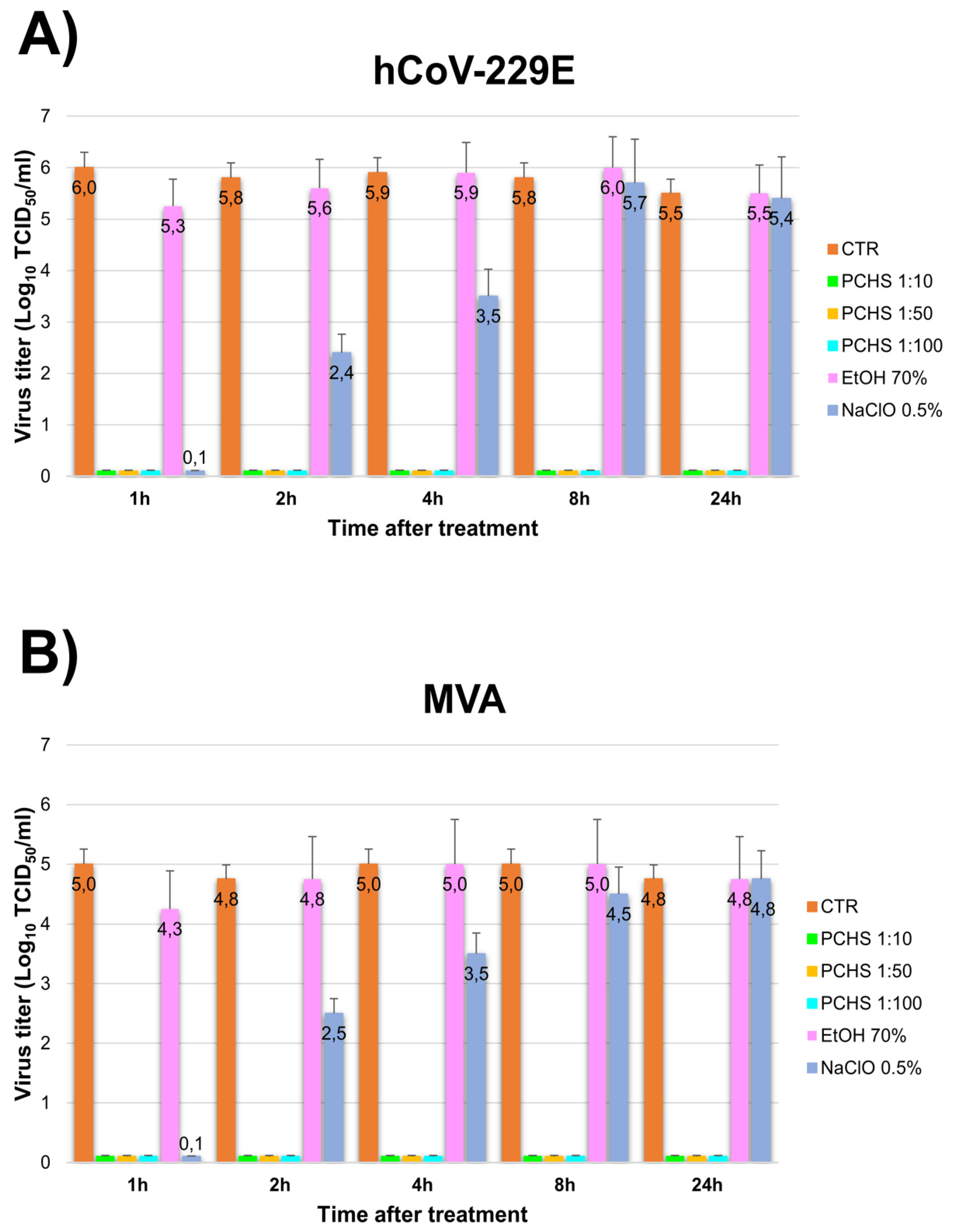
3.1. PCHS Antiviral Activity in Suspension
3.2. PCHS Antiviral Activity on Surface
3.3. Enzymatic Activity of PCHS-Probiotics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 21 October 2021).
- Chin, A.W.H.; Chu, J.T.S.; Perera, M.R.A.; Hui, K.P.Y.; Yen, H.L.; Chan, M.C.W.; Peiris, M.; Poon, L.L.M. Stability of SARS-CoV-2 in different environmental conditions. Lancet Microbe 2020, 1, e10. [Google Scholar] [CrossRef]
- Kampf, G.; Todt, D.; Pfaender, S.; Steinmann, E. Persistence of coronaviruses on inanimate surfaces and their inactivation with biocidal agents. J. Hosp. Infect. 2020, 104, 246–251. [Google Scholar] [CrossRef] [Green Version]
- van Doremalen, N.; Bushmaher, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Azaibi, T.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and surface stability of SARS-CoV-2 as compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Sun, J.; Zhu, A.; Li, H.; Zheng, K.; Zhuang, Z.; Chen, Z.; Shi, Y.; Zhang, Z.; Chen, S.B.; Liu, X.; et al. Isolation of infectious SARS-CoV-2 from urine of a COVID-19 patient. Emerg. Microbes. Infect. 2020, 9, 991–993. [Google Scholar] [CrossRef]
- Wang, J.; Feng, H.; Zhang, S.; Ni, Z.; Ni, L.; Chen, Y.; Zhuo, L.; Zhong, Z.; Qu, T. SARS-CoV-2 rna detection of hospital isolation wards hygiene monitoring during the coronavirus disease 2019 outbreak in a chinese hospital. Int. J. Infect. Dis. 2020, 94, 103–106. [Google Scholar] [CrossRef]
- Chin, A.W.H.; Poon, L.L.M. Stability of SARS-CoV-2 in different environmental conditions—Authors’ reply. Lancet Microbe 2020, 1, e146. [Google Scholar] [CrossRef]
- Liu, Y.; Li, T.; Deng, Y.; Liu, S.; Zhang, D.; Li, H.; Wang, X.; Jia, L.; Han, J.; Bei, Z.; et al. Stability of SARS-CoV-2 on environmental surfaces and in human excreta. J. Hosp. Infect. 2021, 107, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.; Domingo, J.L. Contamination of inert surfaces by SARS-CoV-2: Persistence, stability and infectivity. A review. Environ. Res. 2021, 193, 110559. [Google Scholar] [CrossRef] [PubMed]
- Center for Disease Control and Prevention—CDC. SARS-CoV-2 and Surface (Fomite) Transmission for Indoor Community Environments. 2021. Available online: https://www.cdc.gov/coronavirus/2019-ncov/more/science-and-research/surface-transmission.html (accessed on 9 June 2021).
- Meyerowitz, E.A.; Richterman, A.; Gandhi, R.T.; Sax, P.E. Transmission of SARS-CoV-2: A review of viral, host, and environmental factors. Ann. Intern. Med. 2021, 174, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Kampf, G.; Bruggemann, Y.; Kaba, H.E.J.; Steinmann, J.; Pfaender, S.; Scheithauer, S.; Steinmann, E. Potential sources, modes of transmission and effectiveness of prevention measures against SARS-CoV-2. J. Hosp. Infect. 2020, 106, 678–697. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.M.; Weir, M.H.; Bloomfield, S.F.; Scott, E.A.; Reynolds, K.A. Modeling COVID-19 infection risks for a single hand-to-fomite scenario and potential risk reductions offered by surface disinfection. Am. J. Infect. Control. 2020, 49, 846–848. [Google Scholar] [CrossRef] [PubMed]
- Pitol, A.; Julian, T. Community transmission of SARS-CoV-2 by surfaces: Risks and risk reduction strategies. Environ. Sci. Technol. Lett. 2021, 8, 6. [Google Scholar] [CrossRef]
- Istituto Superiore di Snità—ISS. Interim Recommendations on Cleaning and Disinfection of Non-Healthcare Settings during COVID-19 Health Emergency: Surfaces, Indoor Environments and Clothing. In ISS COVID-19 Working Group on Biocides, ISS, 2020; Rapporto ISS COVID-19 n. 25/2020. Available online: Chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/viewer.html?pdfurl=https%3A%2F%2Fwww.iss.it%2Fdocuments%2F20126%2F0%2FRapporto%2BISS%2BCOVID-19%2B12_2021.pdf%2F4eeb2ce8-648d-b045-4a8c-5dfe1fc7b56a%3Ft%3D1622463358049&clen=2058236 (accessed on 23 October 2021). (In Italian)
- Vandini, A.; Temmerman, R.; Frabetti, A.; Caselli, E.; Antonioli, P.; Balboni, P.G.; Platano, D.; Branchini, A.; Mazzacane, S. Hard Surface Biocontrol in Hospitals Using Microbial-Based Cleaning Products. PLoS ONE 2014, 9, e108598. [Google Scholar] [CrossRef] [Green Version]
- Ghafoor, D.; Khan, Z.; Khan, A.; Ualiyeva, D.; Zaman, N. Excessive use of disinfectants against COVID-19 posing a potential threat to living beings. Curr. Res. Toxicol. 2021, 2, 159–168. [Google Scholar] [CrossRef]
- Curran, E.T.; Wilkinson, M.; Bradley, T. Chemical disinfectants: Controversies regarding their use in low risk healthcare environments (part 1). J. Infect. Prev. 2019, 20, 76–82. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention—CDC. Cleaning and Disinfecting Your Facility; Cleaning, Disinfecting & Ventilation. 2021. Available online: https://www.cdc.gov/coronavirus/2019-ncov/community/disinfecting-building-facility.html (accessed on 27 October 2021).
- Nabi, G.; Wang, Y.; Hao, Y.; Khan, S.; Wu, Y.; Li, D. Massive use of disinfectants against COVID-19 poses potential risks to urban wildlife. Environ. Res. 2020, 188, 109916. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tang, W.; Chen, Y.; Yin, W. Disinfection threatens aquatic ecosystems. Science 2020, 368, 146–147. [Google Scholar] [CrossRef] [Green Version]
- Caselli, E.; D’Accolti, M.; Vandini, A.; Lanzoni, L.; Camerada, M.T.; Coccagna, M.; Branchini, A.; Antonioli, P.; Balboni, P.G.; Di Luca, D.; et al. Impact of a Probiotic-Based Cleaning Intervention on the Microbiota Ecosystem of the Hospital Surfaces: Focus on the Resistome Remodulation. PLoS ONE 2016, 11, e0148857. [Google Scholar] [CrossRef] [Green Version]
- Knight, G.M.; Glover, R.E.; McQuaid, C.F.; Olaru, I.D.; Gallandat, K.; Leclerc, Q.J.; Fuller, N.M.; Willcocks, S.J.; Hasan, R.; van Kleef, E.; et al. Antimicrobial resistance and COVID-19: Intersections and implications. eLife 2021, 10. [Google Scholar] [CrossRef]
- Kampf, G. Biocidal agents used for disinfection can enhance antibiotic resistance in gram-negative species. Antibiotics 2018, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Wand, M.E.; Bock, L.; Bonney, L.C.; Sutton, J.M. Mechanisms of Increased Resistance to Chlorhexidine and Cross-Resistance to Colistin following Exposure of Klebsiella pneumoniae Clinical Isolates to Chlorhexidine. Antimicrob. Agents Chemother. 2017, 61, e01162-16. [Google Scholar] [CrossRef] [Green Version]
- Eterpi, M.; McDonnell, G.; Thomas, V. Disinfection efficacy against parvoviruses compared with reference viruses. J. Hosp. Infect. 2009, 73, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Dublineau, A.; Batéjat, C.; Pinon, A.; Burguière, A.M.; Leclercq, I.; Manuguerra, J.-C. Persistence of the 2009 Pandemic Influenza A (H1N1) Virus in Water and on Non-Porous Surface. PLoS ONE 2011, 6, e28043. [Google Scholar] [CrossRef]
- Kramer, A.; Schwebke, I.; Kampf, G. How long do nosocomial pathogens persist on inanimate surfaces? A systematic review. BMC Infect. Dis. 2006, 6, 130. [Google Scholar] [CrossRef] [Green Version]
- Sizun, J.; Yu, M.; Talbot, P. Survival of human coronaviruses 229E and OC43 in suspension and after drying onsurfaces: A possible source ofhospital-acquired infections. J. Hosp. Infect. 2000, 46, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Hota, B. Contamination, disinfection, and cross-colonization: Are hospital surfaces reservoirs for nosocomial infection? Clin. Infect. Dis. 2004, 39, 1182–1189. [Google Scholar] [PubMed] [Green Version]
- Warnes, S.L.; Little, Z.R.; Keevil, C.W. Human Coronavirus 229E Remains Infectious on Common Touch Surface Materials. mBio 2015, 6, e01697-15. [Google Scholar] [CrossRef] [Green Version]
- Demmler-Harrison, G.J. Healthcare-Associated Viral Infections: Considerations for Nosocomial Transmission and Infection Control; Springer: Cham, Switzerland, 2018; pp. 229–257. [Google Scholar] [CrossRef]
- Carducci, A.; Verani, M.; Lombardi, R.; Casini, B.; Privitera, G. Environmental survey to assess viral contamination of air and surfaces in hospital settings. J. Hosp. Infect. 2011, 77, 242–247. [Google Scholar] [CrossRef]
- Borbone, N.; Piccialli, G.; Roviello, G.N.; Oliviero, G. Nucleoside analogs and nucleoside precursors as drugs in the fight against SARS-CoV-2 and other coronaviruses. Molecules 2021, 26, 986. [Google Scholar] [CrossRef]
- Cheng, A.; Wong, S.M.; Yuan, Y.A. Structural basis for dsRNA recognition by NS1 protein of influenza A virus. Cell Res. 2008, 19, 187–195. [Google Scholar] [CrossRef]
- Tiwari, A.; Patnayak, D.P.; Chander, Y.; Parsad, M.; Goyal, S.M. Survival of Two Avian Respiratory Viruses on Porous and Nonporous Surfaces. Avian Dis. 2006, 50, 284–287. [Google Scholar] [CrossRef]
- Bean, B.; Moore, B.M.; Sterner, B.; Peterson, L.R.; Gerding, D.N.; Balfour, H.H. Survival of Influenza Viruses on Environmental Surfaces. J. Infect. Dis. 1982, 146, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.P.; Choi, Y.W.; Chappie, D.J.; Rogers, J.V.; Kaye, J.Z. Environmental Persistence of a Highly Pathogenic Avian Influenza (H5N1) Virus. Environ. Sci. Technol. 2010, 44, 7515–7520. [Google Scholar] [CrossRef]
- Whitley, R.J.; Roizman, B. Herpes simplex virus infections. Lancet 2001, 357, 1513–1518. [Google Scholar] [CrossRef]
- World Health Organization—WHO. Cleaning and Disinfection of Environmental Surfaces in the Context of COVID-19. 2020. Available online: https://www.who.int/publications/i/item/cleaning-and-disinfection-of-environmental-surfaces-inthe-context-of-covid-19 (accessed on 25 October 2021).
- D’Accolti, M.; Soffritti, I.; Passaro, A.; Zuliani, G.; Antonioli, P.; Mazzacane, S.; Manfredini, R.; Caselli, E. SARS-CoV-2 rna contamination on surfaces of a COVID-19 ward in a hospital of northern italy: What risk of transmission? Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 9202–9207. [Google Scholar] [PubMed]
- Suleyman, G.; Alangaden, G.; Bardossy, A.C. The Role of Environmental Contamination in the Transmission of Nosocomial Pathogens and Healthcare-Associated Infections. Curr. Infect. Dis. Rep. 2018, 20, 12. [Google Scholar] [CrossRef]
- Doll, M.; Stevens, M.; Bearman, G. Environmental cleaning and disinfection of patient areas. Int. J. Infect. Dis. 2018, 67, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caselli, E.; Brusaferro, S.; Coccagna, M.; Arnoldo, L.; Berloco, F.; Antonioli, P.; Tarricone, R.; Pelissero, G.; Nola, S.; La Fauci, V.; et al. Reducing healthcare-associated infections incidence by a probiotic-based sanitation system: A multicentre, prospective, intervention study. PLoS ONE 2018, 13, e0199616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caselli, E.; Antonioli, P.; Mazzacane, S. Safety of probiotics used for hospital environmental sanitation. J. Hosp. Infect. 2016, 94, 193–194. [Google Scholar] [CrossRef]
- Caselli, E.; Arnoldo, L.; Rognoni, C.; D’Accolti, M.; Soffritti, I.; Lanzoni, L.; Bisi, M.; Volta, A.; Tarricone, R.; Brusaferro, S.; et al. Impact of a probiotic-based hospital sanitation on antimicrobial resistance and HAI-associated antimicrobial consumption and costs: A multicenter study. Infect. Drug Resist. 2019, 12, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Caselli, E.; D’Accolti, M.; Soffritti, I.; Lanzoni, L.; Bisi, M.; Volta, A.; Berloco, F.; Mazzacane, S. An Innovative Strategy for the Effective Reduction of MDR Pathogens from the Nosocomial Environment. In Advances in Microbiology, Infectious Diseases and Public Health; Springer: Cham, Switzerland, 2019; Volume 1214, pp. 79–91. [Google Scholar] [CrossRef]
- D’Accolti, M.; Soffritti, I.; Mazzacane, S.; Caselli, E. Fighting AMR in the Healthcare Environment: Microbiome-Based Sanitation Approaches and Monitoring Tools. Int. J. Mol. Sci. 2019, 20, 1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarka, P.; Nitsch-Osuch, A. Evaluating the Virucidal Activity of Disinfectants According to European Union Standards. Viruses 2021, 13, 534. [Google Scholar] [CrossRef] [PubMed]
- Spearman, C. The method of “right and wrong cases” (“constant stimuli”) without gauss’s formulae. Br. J. Psychol. 1908, 2, 227–242. [Google Scholar] [CrossRef]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- European Committee for Standardization—CEN. En 14476:2019: Chemical Disinfectants and Antiseptics—Quantitative Suspension Test for the Evaluation of Virucidal Activity in the Medical Area—Test Method And Requirements (Phase 2/Step 1); European Committee for Standardization: Brussels, Belgium, 2019. [Google Scholar]
- European Committee for Standardization—CEN. En 16777:2019: Chemical Disinfectants and Antiseptics—Quantitative Non-Porous Surface Test Without Mechanical Action for the Evaluation of Virucidal Activity of Chemical Disinfectants Used in the Medical Area—Test Method and Requirements (Phase 2/Step 2); European Committee for Standardization: Brussels, Belgium, 2019. [Google Scholar]
- Chow, E.; Mermel, L.A. Hospital-Acquired Respiratory Viral Infections: Incidence, Morbidity, and Mortality in Pediatric and Adult Patients. Open Forum Infect. Dis. 2017, 4, ofx006. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, M.; Wolff, S.; Ludwig, S.; Schafer, W.; Keiner, B.; Roth, N.J.; Widmer, E. Rapid SARS-CoV-2 inactivation by commonly available chemicals on inanimate surfaces. J. Hosp. Infect. 2020, 106, 633–634. [Google Scholar] [CrossRef]
- Colaneri, M.; Seminari, E.; Piralla, A.; Zuccaro, V.; Filippo, A.D.; Baldanti, F.; Bruno, R.; Mondelli, M.U.; COVID 19 IRCCS San Matteo Pavia Task force. Lack of SARS-CoV-2 rna environmental contamination in a tertiary referral hospital for infectious diseases in northern italy. J. Hosp. Infect. 2020, 105, 3. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, R.; Lo, C.W.; Wada, S.; Somei, J.; Ochiai, H.; Murakami, T.; Saito, N.; Ogawa, T.; Shinjo, A.; Benno, Y.; et al. SARS-CoV-2 disinfection of air and surface contamination by tio2 photocatalyst-mediated damage to viral morphology, RNA, and protein. Viruses 2021, 13, 942. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention—CDC. Hazard Communication for Disinfectants Used against Viruses. 2020. Available online: https://www.cdc.gov/niosh/topics/disinfectant/default.html (accessed on 23 October 2021).
- Rutala, W.A.; Kanamori, H.; Gergen, M.; Weber, D.J. Microbial Assessment of Recontamination with Acinetobacter in Patient Room Environment in Burn Units. Am. J. Infect. Control. 2020, 48, S20. [Google Scholar] [CrossRef]
- Rutala, W.A.; Gergen, M.F.; Sickbert-Bennett, E.E.; Anderson, D.J.; Weber, D.J.; Program, F.T.C.P.E. Antimicrobial activity of a continuously active disinfectant against healthcare pathogens. Infect. Control. Hosp. Epidemiol. 2019, 40, 1284–1286. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization—WHO. Tackling Antimicrobial Resistance in the COVID-19 Pandemic. 2020. Available online: https://www.who.int/bulletin/volumes/98/7/20-268573.pdf (accessed on 20 October 2021).
- Mahnert, A.; Moissl-Eichinger, C.; Zojer, M.; Bogumil, D.; Mizrahi, I.; Rattei, T.; Martinez, J.L.; Berg, G. Man-made microbial resistances in built environments. Nat. Commun. 2019, 10, 968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, D.; Prussin, A.J., 2nd; Marr, L.C.; Vikesland, P.J.; Edwards, M.A.; Pruden, A. Factors shaping the human exposome in the built environment: Opportunities for engineering control. Environ. Sci. Technol. 2017, 51, 7759–7774. [Google Scholar] [CrossRef] [Green Version]
- Al-Ghalith, G.A.; Knights, D. Bygiene: The New Paradigm of Bidirectional Hygiene. Yale J. Boil. Med. 2015, 88, 359–365. [Google Scholar]
- Caselli, E. Hygiene: Microbial strategies to reduce pathogens and drug resistance in clinical settings. Microb. Biotechnol. 2017, 10, 1079–1083. [Google Scholar] [CrossRef] [Green Version]
- D’Accolti, M.; Soffritti, I.; Piffanelli, M.; Bisi, M.; Mazzacane, S.; Caselli, E. Efficient removal of hospital pathogens from hard surfaces by a combined use of bacteriophages and probiotics: Potential as sanitizing agents. Infect. Drug Resist. 2018, 11, 1015–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kormuth, K.A.; Lin, K.; Qian, Z.; Myerburg, M.M.; Marr, L.C.; Lakdawala, S.S. Environmental Persistence of Influenza Viruses Is Dependent upon Virus Type and Host Origin. mSphere 2019, 4, e00552-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahromi, R.; Mogharab, V.; Jahromi, H.; Avazpour, A. Synergistic effects of anionic surfactants on coronavirus (SARS-CoV-2) virucidal efficiency of sanitizing fluids to fight COVID-19. Food Chem. Toxicol. 2020, 145, 111702. [Google Scholar] [CrossRef]
- Latorre, J.D.; Hernandez-Velasco, X.; Wolfenden, R.E.; Vicente, J.L.; Wolfenden, A.D.; Menconi, A.; Bielke, L.R.; Hargis, B.M.; Tellez, G. Evaluation and Selection of Bacillus Species Based on Enzyme Production, Antimicrobial Activity, and Biofilm Synthesis as Direct-Fed Microbial Candidates for Poultry. Front. Vet. Sci. 2016, 3, 95. [Google Scholar] [CrossRef] [Green Version]
- Danilova, I.; Sharipova, M. The Practical Potential of Bacilli and Their Enzymes for Industrial Production. Front. Microbiol. 2020, 11, 1782. [Google Scholar] [CrossRef]
- Delhalle, L.; Taminiau, B.; Fastrez, S.; Fall, A.; Ballesteros, M.; Burteau, S.; Daube, G. Evaluation of Enzymatic Cleaning on Food Processing Installations and Food Products Bacterial Microflora. Front. Microbiol. 2020, 11, 1827. [Google Scholar] [CrossRef]
- Tarricone, R.; Rognoni, C.; Arnoldo, L.; Mazzacane, S.; Caselli, E. A Probiotic-Based Sanitation System for the Reduction of Healthcare Associated Infections and Antimicrobial Resistances: A Budget Impact Analysis. Pathogens 2020, 9, 502. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Standardization—CEN. En 14885:2018: Chemical Disinfectants and Antiseptics—Application of European Standards for Chemical Disinfectants and Antiseptics; European Committee for Standardization: Brussels, Belgium, 2018. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Strain | Target Cells |
|---|---|---|
| modified Vaccinia virus Ankara (MVA) | ATCC VR-1508 | baby hamster kidney fibroblast BHK-21 cell line (ATCC CCL-10) |
| herpes simplex virus type 1 (HSV-1) | ATCC VR-260 | monkey kidney fibroblast Vero-E6 cell line (ATCC CRL-1586) |
| human alpha-coronavirus 229E (hCoV-229E) | ATCC VR-740 | human lung fibroblast MRC-5 cells (ATCC CCL-171) |
| human beta-coronavirus SARS-CoV-2 1 | // | monkey kidney fibroblast Vero-E6 cell line (ATCC CRL-1586) |
| human H3N2 influenza virus 2 | A/Wisconsin/67/2005 | Madin-Darby Canine Kidney MDCK cell line (ATCC CCL-34) |
| avian H10N1 influenza virus 2 | A/mallard/Italy/4518/2012 | Madin-Darby Canine Kidney MDCK cell line (ATCC CCL-34) |
| swine H1N2 influenza virus 2 | A/swine/Italy/4159/2006 | Madin-Darby Canine Kidney MDCK cell line (ATCC CCL-34) |
| Enzyme | B. subtilis | B. pumilus | B. megaterium |
|---|---|---|---|
| NTC Alkaline phosphatase Esterase | 0 | 0 | 0 |
| 2 | 5 | 4 | |
| 4 | 4 | 2 | |
| Esterase lipase Lipase | 4 | 4 | 2 |
| 0 | 0 | 0 | |
| Leucine arylamidase Valine arylamidase Cysteine arylamidase Trypsin | 0 | 3 | 3 |
| 0 | 1 | 1 | |
| 0 | 0 | 0 | |
| 0 | 0 | 0 | |
| α-chimotrypsin Acid phosphatase | 0 | 1 | 1 |
| 2 | 4 | 5 | |
| Naphtol-phosphohydrolase | 1 | 1 | 2 |
| α-galactosidase | 2 | 1 | 0 |
| β-galactosidase | 0 | 4 | 2 |
| β-glucuronidase | 0 | 0 | 0 |
| α-glucosidase | 4 | 1 | 2 |
| β-glucosidase | 5 | 5 | 1 |
| N-acetyl-β-glucosamidase | 1 | 1 | 1 |
| α-mannosidase | 0 | 3 | 0 |
| α-fucosidase | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Accolti, M.; Soffritti, I.; Bonfante, F.; Ricciardi, W.; Mazzacane, S.; Caselli, E. Potential of an Eco-Sustainable Probiotic-Cleaning Formulation in Reducing Infectivity of Enveloped Viruses. Viruses 2021, 13, 2227. https://doi.org/10.3390/v13112227
D’Accolti M, Soffritti I, Bonfante F, Ricciardi W, Mazzacane S, Caselli E. Potential of an Eco-Sustainable Probiotic-Cleaning Formulation in Reducing Infectivity of Enveloped Viruses. Viruses. 2021; 13(11):2227. https://doi.org/10.3390/v13112227
Chicago/Turabian StyleD’Accolti, Maria, Irene Soffritti, Francesco Bonfante, Walter Ricciardi, Sante Mazzacane, and Elisabetta Caselli. 2021. "Potential of an Eco-Sustainable Probiotic-Cleaning Formulation in Reducing Infectivity of Enveloped Viruses" Viruses 13, no. 11: 2227. https://doi.org/10.3390/v13112227
APA StyleD’Accolti, M., Soffritti, I., Bonfante, F., Ricciardi, W., Mazzacane, S., & Caselli, E. (2021). Potential of an Eco-Sustainable Probiotic-Cleaning Formulation in Reducing Infectivity of Enveloped Viruses. Viruses, 13(11), 2227. https://doi.org/10.3390/v13112227