
From Protein to Pandemic: The Transdisciplinary Approach Needed to Prevent Spillover and the Next Pandemic


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Transdisciplinary Science to Identify Drivers of Pandemics
2. The Next Pandemic: Preventing Spillover Is the Ultimate Approach
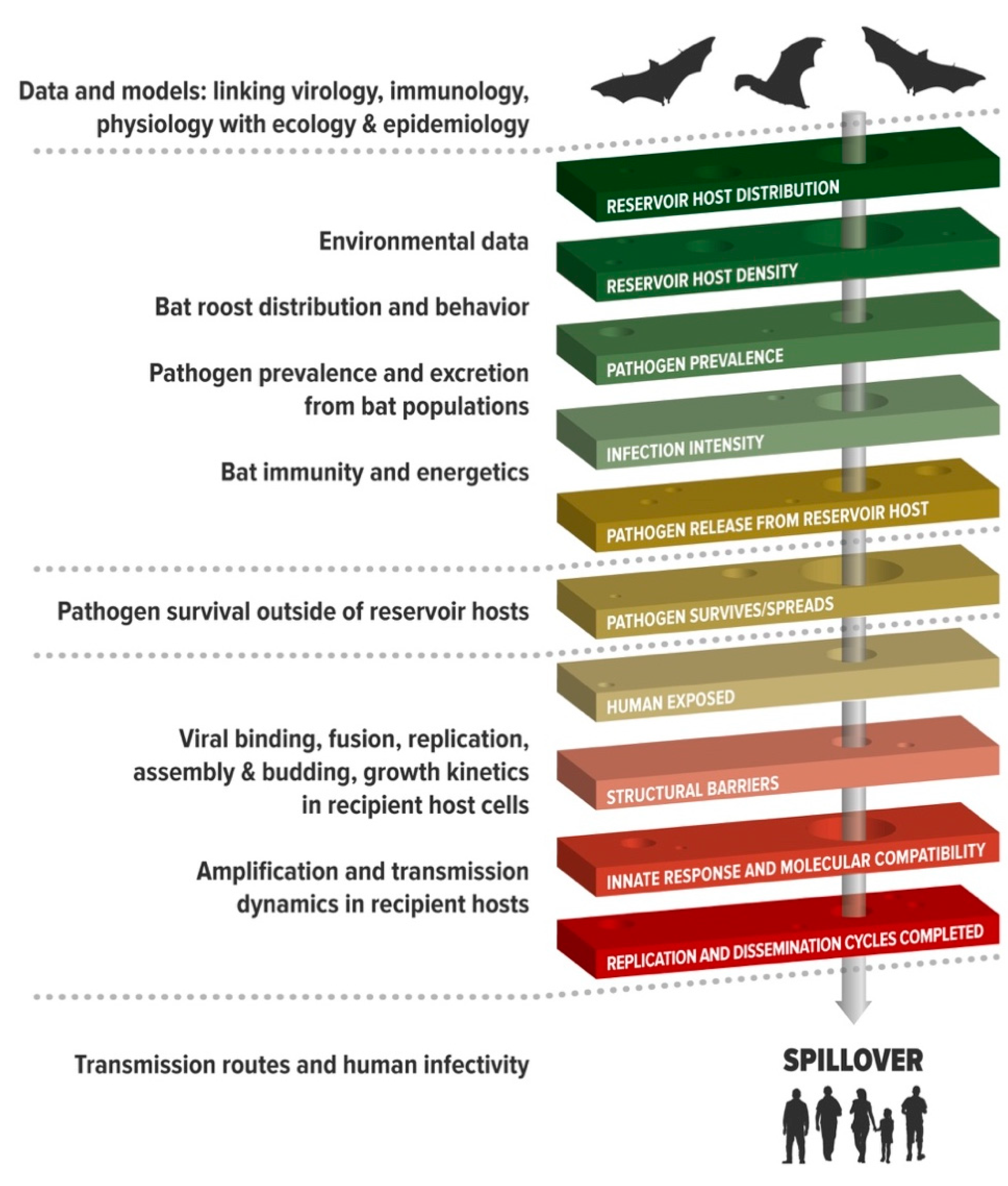
3. Systems with Replicated Spillover Events Provide Insights into the Processes
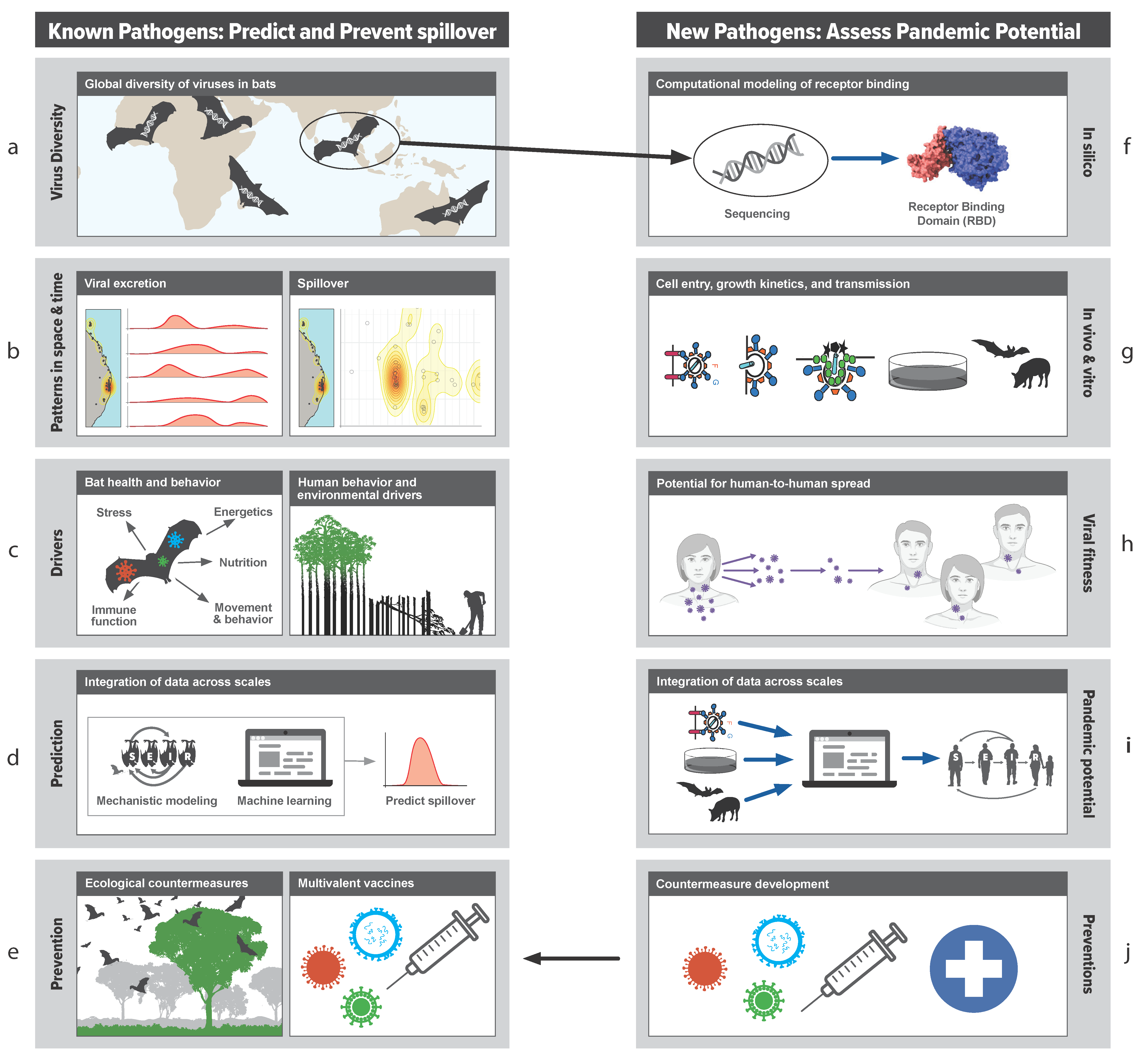
4. Transdisciplinary Approach to Stop Spillover
4.1. Ecology of the Reservoir Host
4.2. Viral Shedding from Reservoir Hosts and Its Drivers
4.3. Bat and Human Behavior and Proximity
4.4. Data Integration
4.5. Functional Characterization of Viruses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.; Ko, A.; Graham, A.L.; Lloyd-Smith, J.O. Pathways to zoonotic spillover. Nat. Rev. Genet. 2017, 15, 502–510. [Google Scholar] [CrossRef] [Green Version]
- Plowright, R.K.; Reaser, J.K.; Locke, H.; Woodley, S.J.; A Patz, J.; Becker, D.J.; Oppler, G.; Hudson, P.J.; Tabor, G.M. Land use-induced spillover: A call to action to safeguard environmental, animal, and human health. Lancet Planet. Health 2021, 5, e237–e245. [Google Scholar] [CrossRef]
- Wasik, B.R.; De Wit, E.; Munster, V.; Lloyd-Smith, J.O.; Martinez-Sobrido, L.; Parrish, C. Onward transmission of viruses: How do viruses emerge to cause epidemics after spillover? Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190017. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Smith, J.O.; George, D.; Pepin, K.M.; Pitzer, V.; Pulliam, J.; Dobson, A.P.; Hudson, P.; Grenfell, B.T. Epidemic Dynamics at the Human-Animal Interface. Science 2009, 326, 1362–1367. [Google Scholar] [CrossRef] [Green Version]
- Cutler, D.M.; Summers, L.H. The COVID-19 Pandemic and the $16 Trillion Virus. JAMA 2020, 324, 1495–1496. [Google Scholar] [CrossRef]
- Morens, D.M.; Folkers, G.K.; Fauci, A.S. The challenge of emerging and re-emerging infectious diseases. Nat. Cell Biol. 2004, 430, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Morens, D.M.; Fauci, A.S. Emerging Infectious Diseases in 2012: 20 Years after the Institute of Medicine Report. mBio 2012, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Morse, S.S.; Mazet, J.A.; Woolhouse, M.; Parrish, C.R.; Carroll, D.; Karesh, W.B.; Zambrana-Torrelio, C.; Lipkin, W.I.; Daszak, P. Prediction and prevention of the next pandemic zoonosis. Lancet 2012, 380, 1956–1965. [Google Scholar] [CrossRef]
- Dobson, A.P.; Pimm, S.L.; Hannah, L.; Kaufman, L.; Ahumada, J.A.; Ando, A.W.; Bernstein, A.; Busch, J.; Daszak, P.; Engelmann, J.; et al. Ecology and economics for pandemic prevention. Science 2020, 369, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Coltart, C.E.; Lindsey, B.; Ghinai, I.; Johnson, A.M.; Heymann, D.L. The Ebola outbreak, 2013–2016: Old lessons for new epi-demics. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160297. [Google Scholar] [CrossRef]
- Parrish, C.R.; Holmes, E.; Morens, D.M.; Park, E.-C.; Burke, D.S.; Calisher, C.H.; Laughlin, C.A.; Saif, L.J.; Daszak, P. Cross-Species Virus Transmission and the Emergence of New Epidemic Diseases. Microbiol. Mol. Biol. Rev. 2008, 72, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Pepin, K.M.; Lass, S.; Pulliam, J.; Read, A.F.; Lloyd-Smith, J. Identifying genetic markers of adaptation for surveillance of viral host jumps. Nat. Rev. Genet. 2010, 8, 802–813. [Google Scholar] [CrossRef]
- Allison, A.B.; Harbison, C.E.; Pagan, I.; Stucker, K.; Kaelber, J.; Brown, J.D.; Ruder, M.G.; Keel, M.K.; Dubovi, E.J.; Holmes, E.; et al. Role of Multiple Hosts in the Cross-Species Transmission and Emergence of a Pandemic Parvovirus. J. Virol. 2011, 86, 865–872. [Google Scholar] [CrossRef] [Green Version]
- Sauter, D.; Kirchhoff, F. Key Viral Adaptations Preceding the AIDS Pandemic. Cell Host Microbe 2019, 25, 27–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washburne, A.D.; Crowley, D.E.; Becker, D.J.; Manlove, K.R.; Childs, M.L.; Plowright, R.K. Percolation models of pathogen spillover. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, D.J.; Washburne, A.D.; Faust, C.L.; Pulliam, J.R.C.; Mordecai, E.A.; Lloyd-Smith, J.O.; Plowright, R.K. Dynamic and integrative approaches to understanding pathogen spillover. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190014. [Google Scholar] [CrossRef] [PubMed]
- Cross, P.C.; Prosser, D.J.; Ramey, A.M.; Hanks, E.M.; Pepin, K.M. Confronting models with data: The challenges of estimating disease spillover. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restif, O.; Hayman, D.T.; Pulliam, J.R.; Plowright, R.K.; George, D.B.; Luis, A.D.; Cunningham, A.A.; Bowen, R.A.; Fooks, A.R.; O’Shea, T.J. Model-guided fieldwork: Practical guidelines for multidisciplinary research on wildlife ecological and epide-miological dynamics. Ecol. Lett. 2012, 15, 1083–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacono, G.L.; Cunningham, A.A.; Fichet-Calvet, E.; Garry, R.; Grant, D.S.; Leach, M.; Moses, L.M.; Nichols, G.; Schieffelin, J.S.; Shaffer, J.G.; et al. A Unified Framework for the Infection Dynamics of Zoonotic Spillover and Spread. PLoS Negl. Trop. Dis. 2016, 10, e0004957. [Google Scholar] [CrossRef]
- Gorman, M.E.; Jenkins, L.; Plowright, R.K. Human interactions and sustainability. In Sustainability: Multi-Disciplinary Perspectives; Cabezas, H., Diwekar, U., Eds.; Bentham Open E-Books: Sharjah, United Arab Emirates, 2011; pp. 88–111. [Google Scholar]
- Taylor, L.H.; Latham, S.M.; Woolhouse, M. Risk factors for human disease emergence. Philos. Trans. R. Soc. B Biol. Sci. 2001, 356, 983–989. [Google Scholar] [CrossRef]
- Olival, K.J.; Hosseini, P.R.; Zambrana-Torrelio, C.; Ross, N.; Bogich, T.; Daszak, P. Host and viral traits predict zoonotic spillover from mammals. Nat. Cell Biol. 2017, 546, 646–650. [Google Scholar] [CrossRef]
- Cator, L.J.; Johnson, L.R.; Mordecai, E.A.; El Moustaid, F.; Smallwood, T.R.C.; LaDeau, S.L.; Johansson, M.A.; Hudson, P.J.; Boots, M.; Thomas, M.B.; et al. The Role of Vector Trait Variation in Vector-Borne Disease Dynamics. Front. Ecol. Evol. 2020, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Bonneaud, C.; Weinert, L.A.; Kuijper, B. Understanding the emergence of bacterial pathogens in novel hosts. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, D.A.; Read, A.F. Why the evolution of vaccine resistance is less of a concern than the evolution of drug resistance. Proc. Natl. Acad. Sci. USA 2018, 115, 12878–12886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, A.F.; Woods, R.J. Antibiotic resistance management. Evol. Med. Public Heal. 2014, 2014, 147. [Google Scholar] [CrossRef] [PubMed]
- Saéz, A.M.; Weiss, S.; Nowak, K.; Lapeyre, V.; Zimmermann, F.; Düx, A.; Kühl, H.S.; Kaba, M.; Regnaut, S.; Merkel, K.; et al. Investigating the zoonotic origin of the West African Ebola epidemic. EMBO Mol. Med. 2015, 7, 17–23. [Google Scholar] [CrossRef]
- World Health Organization. WHO-Convened Global Study of Origins of SARS-CoV-2: China Part; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Boni, M.F.; Lemey, P.; Jiang, X.; Lam, T.T.-Y.; Perry, B.W.; Castoe, T.A.; Rambaut, A.; Robertson, D.L. Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic. Nat. Microbiol. 2020, 5, 1408–1417. [Google Scholar] [CrossRef]
- Anthony, S.J.; Johnson, C.K.; Greig, D.J.; Kramer, S.; Che, X.; Wells, H.; Hicks, A.L.; Joly, D.O.; Wolfe, N.D.; Daszak, P.; et al. Global patterns in coronavirus diversity. Virus Evol. 2017, 3, vex012. [Google Scholar] [CrossRef]
- Letko, M.; Seifert, S.N.; Olival, K.J.; Plowright, R.K.; Munster, V.J. Bat-borne virus diversity, spillover and emergence. Nat. Rev. Genet. 2020, 18, 461–471. [Google Scholar] [CrossRef]
- Holmes, E.C.; Rambaut, A.; Andersen, K.G. Pandemics: Spend on surveillance, not prediction. Nat. Cell Biol. 2018, 558, 180–182. [Google Scholar] [CrossRef]
- Gray, G.; Robie, E.; Studstill, C.; Nunn, C. Mitigating Future Respiratory Virus Pandemics: New Threats and Approaches to Consider. Viruses 2021, 13, 637. [Google Scholar] [CrossRef] [PubMed]
- Plowright, R.K.; Eby, P.; Hudson, P.; Smith, I.L.; Westcott, D.; Bryden, W.L.; Middleton, D.; Reid, P.A.; McFarlane, R.A.; Martin, G.; et al. Ecological dynamics of emerging bat virus spillover. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2015, 282, 20142124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Li, S.-Y.; Yang, X.-L.; Huang, H.-M.; Zhang, Y.-J.; Guo, H.; Luo, C.-M.; Miller, M.; Zhu, G.; Chmura, A.; et al. Serological Evidence of Bat SARS-Related Coronavirus Infection in Humans, China. Virol. Sin. 2018, 33, 104–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suwannarong, K.; Janetanakit, T.; Kanthawee, P.; Suwannarong, K.; Theamboonlers, A.; Poovorawan, Y.; Tun, H.M.; Chanabun, S.; Amonsin, A. Coronavirus seroprevalence among villagers exposed to bats in Thailand. Zoonoses Public Health. 2021. [Google Scholar] [CrossRef]
- McKee, C.D.; Islam, A.; Luby, S.P.; Salje, H.; Hudson, P.J.; Plowright, R.K.; Gurley, E.S. The ecology of nipah virus in Bang-ladesh: A nexus of land-use change and opportunistic feeding behavior in bats. Viruses 2021, 13, 169. [Google Scholar] [CrossRef] [PubMed]
- Nikolay, B.; Salje, H.; Hossain, M.J.; Khan, A.D.; Sazzad, H.; Rahman, M.; Daszak, P.; Ströher, U.; Pulliam, J.R.; Kilpatrick, A.M.; et al. Transmission of Nipah Virus—14 Years of Investigations in Bangladesh. N. Engl. J. Med. 2019, 380, 1804–1814. [Google Scholar] [CrossRef]
- Luby, S.P.; Hossain, M.J.; Gurley, E.S.; Ahmed, B.-N.; Banu, S.; Khan, M.S.U.; Homaira, N.; Rota, P.A.; Rollin, P.; Comer, J.A.; et al. Recurrent Zoonotic Transmission of Nipah Virus into Humans, Bangladesh, 2001–2007. Emerg. Infect. Dis. 2009, 15, 1229–1235. [Google Scholar] [CrossRef]
- Dobzhansky, T. Nothing in Biology Makes Sense except in the Light of Evolution. Am. Biol. Teach. 1973, 35, 125–129. [Google Scholar] [CrossRef]
- Plowright, R.K.; Becker, D.J.; McCallum, H.; Manlove, K.R. Sampling to elucidate the dynamics of infections in reservoir hosts. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180336. [Google Scholar] [CrossRef] [Green Version]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mamm. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef]
- Voigt, C.C.; Kingston, T. Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer Nature: Basingstoke, UK, 2016. [Google Scholar]
- Romero, L.M.; Wingfield, J.C. Tempests, Poxes, Predators, and People: Stress in Wild Animals and How They Cope; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Voigt, C.C.; Frick, W.F.; Holderied, M.W.; Holland, R.; Kerth, G.; Mello, M.A.R.; Plowright, R.K.; Swartz, S.; Yovel, Y. Principles and Patterns of Bat Movements: From Aerodynamics to Ecology. Q. Rev. Biol. 2017, 92, 267–287. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.S.; Epstein, J.H.; Breed, A.; Plowright, R.K.; Olival, K.J.; De Jong, C.; Daszak, P.; Field, H.E. Satellite Telemetry and Long-Range Bat Movements. PLoS ONE 2011, 6, e14696. [Google Scholar] [CrossRef] [Green Version]
- Giles, J.R.; Eby, P.; Parry, H.; Peel, A.J.; Plowright, R.K.; Westcott, D.A.; McCallum, H. Environmental drivers of spatiotemporal foraging intensity in fruit bats and implications for Hendra virus ecology. Sci. Rep. 2018, 8, 9555. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.B.; Addison, B.; Bean, A.G.; Buchanan, K.L.; Crino, O.L.; Eastwood, J.; Flies, A.S.; Hamede, R.; Hill, G.E.; Klaassen, M.; et al. Extreme Competence: Keystone Hosts of Infections. Trends Ecol. Evol. 2019, 34, 303–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, D.J.; Crowley, D.E.; Washburne, A.D.; Plowright, R.K. Temporal and spatial limitations in global surveillance for bat filoviruses and henipaviruses. Biol. Lett. 2019, 15, 20190423. [Google Scholar] [CrossRef] [PubMed]
- Keeling, M.J.; Bjørnstad, O.N.; Grenfell, B.T. Metapopulation Dynamics of Infectious Diseases. In Ecology, Genetics and Evolution of Metapopulations; Elsevier BV: Amsterdam, The Netherlands, 2004; pp. 415–445. [Google Scholar]
- Plowright, R.K.; Peel, A.; Streicker, D.G.; Gilbert, A.T.; McCallum, H.; Wood, J.; Baker, M.L.; Restif, O. Transmission or Within-Host Dynamics Driving Pulses of Zoonotic Viruses in Reservoir–Host Populations. PLoS Negl. Trop. Dis. 2016, 10, e0004796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paez, D.J.; Giles, J.; McCallum, H.; Field, H.; Jordan, D.; Peel, A.J.; Plowright, R.K. Conditions affecting the timing and mag-nitude of Hendra virus shedding across pteropodid bat populations in Australia. Epidemiol. Infect. 2017, 145, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Smith, J.O.; Schreiber, S.J.; Kopp, P.E.; Getz, W. Superspreading and the effect of individual variation on disease emergence. Nat. Cell Biol. 2005, 438, 355–359. [Google Scholar] [CrossRef]
- Becker, D.J.; Albery, G.F.; Kessler, M.K.; Lunn, T.J.; Falvo, C.A.; Czirják, G.Á.; Martin, L.B.; Plowright, R.K. Macroimmunology: The drivers and consequences of spatial patterns in wildlife immune defence. J. Anim. Ecol. 2020, 89, 972–995. [Google Scholar] [CrossRef]
- Peel, A.J.; Wells, K.; Giles, J.; Boyd, V.; Burroughs, A.; Edson, D.; Crameri, G.; Baker, M.L.; Field, H.; Wang, L.-F.; et al. Synchronous shedding of multiple bat paramyxoviruses coincides with peak periods of Hendra virus spillover. Emerg. Microbes Infect. 2019, 8, 1314–1323. [Google Scholar] [CrossRef] [Green Version]
- Hoegh, A.; Peel, A.; Madden, W.; Ruiz-Aravena, M.; Morris, A.; Washburne, A.; Plowright, R. Estimating Viral Prevalence with Data Integration for Adaptive Two-Phase Pooled Sampling. Ecol. Evol. accepted. [CrossRef]
- Giles, J.R.; Peel, A.J.; Wells, K.; Plowright, R.K.; McCallum, H.; Restif, O. Optimizing non-invasive sampling of a zoonotic bat virus. Ecol. Evol. in press.
- Glennon, E.E.; Becker, D.J.; Peel, A.; Garnier, R.; Suu-Ire, R.D.; Gibson, L.; Hayman, D.T.S.; Wood, J.L.N.; Cunningham, A.A.; Plowright, R.K.; et al. What is stirring in the reservoir? Modelling mechanisms of henipavirus circulation in fruit bat hosts. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190021. [Google Scholar] [CrossRef] [Green Version]
- Kessler, M.K.; Becker, D.J.; Peel, A.J.; Justice, N.V.; Lunn, T.; Crowley, D.E.; Jones, D.N.; Eby, P.; Sánchez, C.A.; Plowright, R.K. Changing resource landscapes and spillover of henipaviruses. Ann. N. Y. Acad. Sci. 2018, 1429, 78–99. [Google Scholar] [CrossRef] [PubMed]
- Flies, A.S.; Wild Comparative Immunology Consortium. Rewilding immunology. Science 2020, 369, 37–38. [Google Scholar] [CrossRef]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel insights into immune systems of bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Plowright, R.K.; Sokolow, S.; E Gorman, M.; Daszak, P.; E Foley, J. Causal inference in disease ecology: Investigating ecological drivers of disease emergence. Front. Ecol. Environ. 2008, 6, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Mendez, D.H.; Kelly, J.; Buttner, P.; Nowak, M.; Speare, R. Management of the slowly emerging zoonosis, Hendra virus, by private veterinarians in Queensland, Australia: A qualitative study. BMC Veter Res. 2014, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Plowright, R.K.; Becker, D.J.; Crowley, D.E.; Washburne, A.D.; Huang, T.; Nameer, P.O.; Gurley, E.S.; Han, B.A. Prioritizing surveillance of Nipah virus in India. PLoS Negl. Trop. Dis. 2019, 13, e0007393. [Google Scholar] [CrossRef] [Green Version]
- Guenther, S.K.; Shanahan, E.A. Communicating risk in human-wildlife interactions: How stories and images move minds. PLoS ONE 2020, 15, e0244440. [Google Scholar] [CrossRef]
- E Field, H. Hendra virus ecology and transmission. Curr. Opin. Virol. 2016, 16, 120–125. [Google Scholar] [CrossRef]
- Peel, A.J.; Field, H.E.; Reid, P.A.; Plowright, R.K.; Broder, C.C.; Skerratt, L.F.; Hayman, D.T.S.; Restif, O.; Taylor, M.; Martin, G.; et al. The equine Hendra virus vaccine remains a highly effective preventative measure against infection in horses and humans: ‘The imperative to develop a human vaccine for the Hendra virus in Australia’. Infect. Ecol. Epidemiol. 2016, 6, 31658. [Google Scholar] [CrossRef]
- Middleton, D.; Pallister, J.; Klein, R.; Feng, Y.-R.; Haining, J.; Arkinstall, R.; Frazer, L.; Huang, J.-A.; Edwards, N.; Wareing, M.; et al. Hendra Virus Vaccine, a One Health Approach to Protecting Horse, Human, and Environmental Health. Emerg. Infect. Dis. 2014, 20, 372–379. [Google Scholar] [CrossRef]
- Khan, S.U.; Gurley, E.S.; Hossain, M.J.; Nahar, N.; Sharker, M.A.Y.; Luby, S.P. A Randomized Controlled Trial of Interventions to Impede Date Palm Sap Contamination by Bats to Prevent Nipah Virus Transmission in Bangladesh. PLoS ONE 2012, 7, e42689. [Google Scholar] [CrossRef]
- Webster, R.G. Wet markets—A continuing source of severe acute respiratory syndrome and influenza? Lancet 2004, 363, 234–236. [Google Scholar] [CrossRef] [Green Version]
- Saylors, K.; PREDICT Consortium; Wolking, D.J.; Hagan, E.; Martinez, S.; Francisco, L.; Euren, J.; Olson, S.H.; Miller, M.; Fine, A.E.; et al. Socializing One Health: An innovative strategy to investigate social and behavioral risks of emerging viral threats. One Heal. Outlook 2021, 3, 1–16. [Google Scholar] [CrossRef]
- Lunn, T.J.; Restif, O.; Peel, A.; Munster, V.; De Wit, E.; Sokolow, S.; Van Doremalen, N.; Hudson, P.; McCallum, H. Dose–response and transmission: The nexus between reservoir hosts, environment and recipient hosts. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190016. [Google Scholar] [CrossRef] [Green Version]
- Han, B.A.; Kramer, A.; Drake, J.M. Global Patterns of Zoonotic Disease in Mammals. Trends Parasitol. 2016, 32, 565–577. [Google Scholar] [CrossRef]
- Becker, D.; Albery, G.F.; Sjodin, A.R.; Poisot, T.; Dallas, T.; Eskew, E.A.; Farrell, M.J.; Guth, S.; Han, B.A.; Simmons, N.B. Pre-dicting wildlife hosts of betacoronaviruses for SARS-CoV-2 sampling prioritization: A modeling study. bioRxiv 2020. [Google Scholar] [CrossRef]
- Becker, D.J.; Washburne, A.D.; Faust, C.L.; Mordecai, E.A.; Plowright, R.K. The problem of scale in the prediction and man-agement of pathogen spillover. Philos. Trans. R. Soc. B 2019, 374, 20190224. [Google Scholar] [CrossRef] [PubMed]
- Reaser, J.K.; Witt, A.; Tabor, G.M.; Hudson, P.J.; Plowright, R.K. Ecological countermeasures for preventing zoonotic disease outbreaks: When ecological restoration is a human health imperative. Restor. Ecol. 2021, 29, e13357. [Google Scholar] [CrossRef] [PubMed]
- Crowley, D.; Becker, D.; Washburne, A.; Plowright, R. Identifying Suspect Bat Reservoirs of Emerging Infections. Vaccines 2020, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Pernet, O.; E Wang, Y.; Lee, B. Henipavirus Receptor Usage and Tropism. Curr. Top. Microbiol. Immunol. 2012, 359, 59–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, H.C.; Iorio, R.M. Henipavirus Membrane Fusion and Viral Entry. Curr. Top. Microbiol. Immunol. 2012, 359, 79–94. [Google Scholar] [CrossRef]
- Lee, B.; Ataman, Z.A. Modes of paramyxovirus fusion: A Henipavirus perspective. Trends Microbiol. 2011, 19, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Fischhoff, I.R.; Castellanos, A.A.; Rodrigues, J.P.; Varsani, A.; Han, B.A. Predicting the zoonotic capacity of mammal species for SARS-CoV-2. bioRxiv 2021. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Feldmann, H.; Broder, C.C. Animal Challenge Models of Henipavirus Infection and Pathogenesis. Curr. Top. Microbiol. Immunol. 2012, 359, 153–177. [Google Scholar] [CrossRef]
- Russell, C.A.; Kasson, P.M.; Donis, R.O.; Riley, S.; Dunbar, J.; Rambaut, A.; Asher, J.; Burke, S.; Davis, C.T.; Garten, R.J. Science Forum: Improving pandemic influenza risk assessment. Elife 2014, 3, e03883. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plowright, R.K.; Hudson, P.J. From Protein to Pandemic: The Transdisciplinary Approach Needed to Prevent Spillover and the Next Pandemic. Viruses 2021, 13, 1298. https://doi.org/10.3390/v13071298
Plowright RK, Hudson PJ. From Protein to Pandemic: The Transdisciplinary Approach Needed to Prevent Spillover and the Next Pandemic. Viruses. 2021; 13(7):1298. https://doi.org/10.3390/v13071298
Chicago/Turabian StylePlowright, Raina K., and Peter J. Hudson. 2021. "From Protein to Pandemic: The Transdisciplinary Approach Needed to Prevent Spillover and the Next Pandemic" Viruses 13, no. 7: 1298. https://doi.org/10.3390/v13071298
APA StylePlowright, R. K., & Hudson, P. J. (2021). From Protein to Pandemic: The Transdisciplinary Approach Needed to Prevent Spillover and the Next Pandemic. Viruses, 13(7), 1298. https://doi.org/10.3390/v13071298