
Viral Load and Patterns of SARS-CoV-2 Dissemination to the Lungs, Mediastinal Lymph Nodes, and Spleen of Patients with COVID-19 Associated Lymphopenia

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. RNA Extraction and cDNA Production
2.2. Preparation of Calibration Standards
2.3. Quantitative RT-PCR
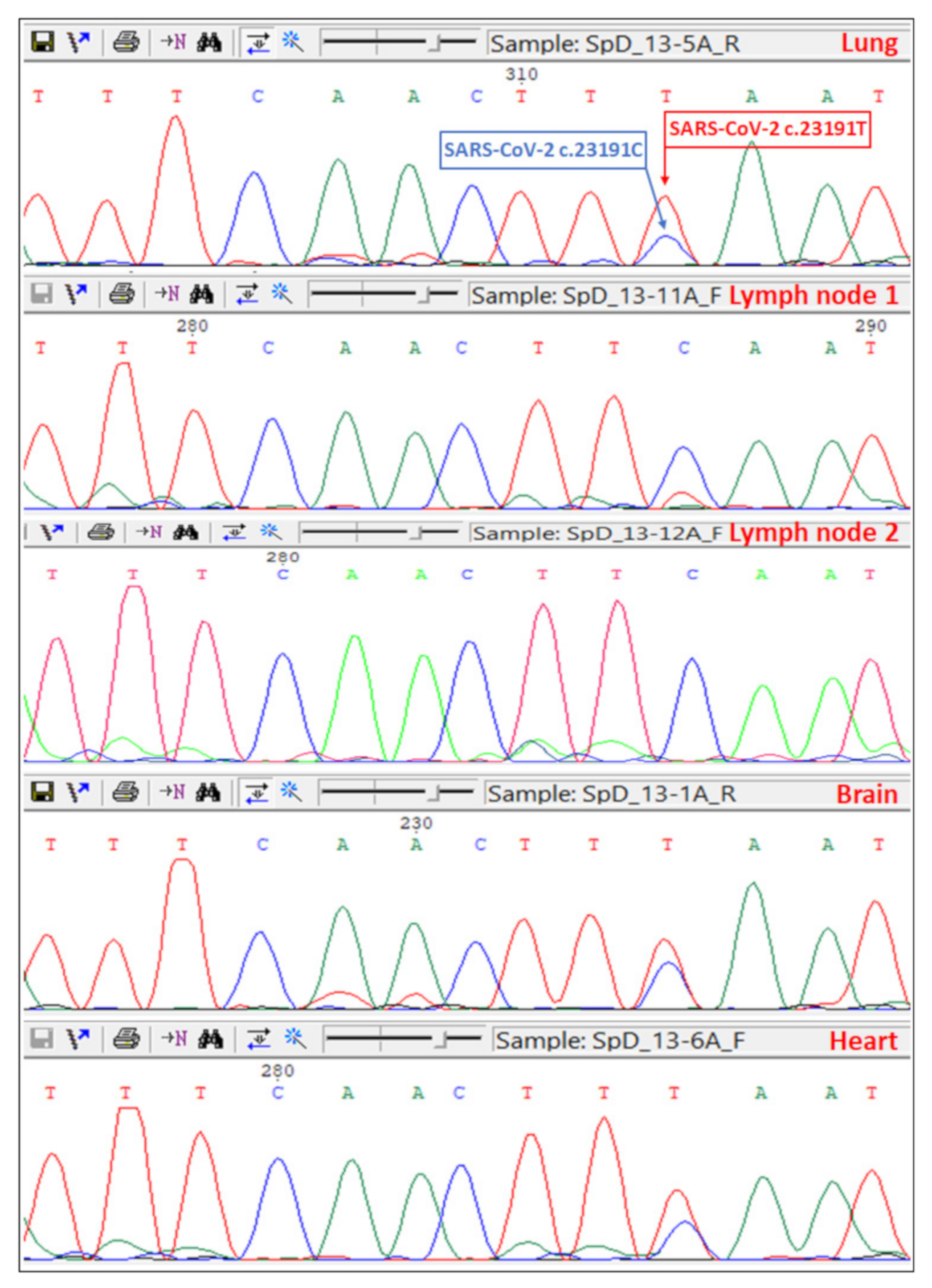
2.4. Sanger Sequencing of the Spike Gene
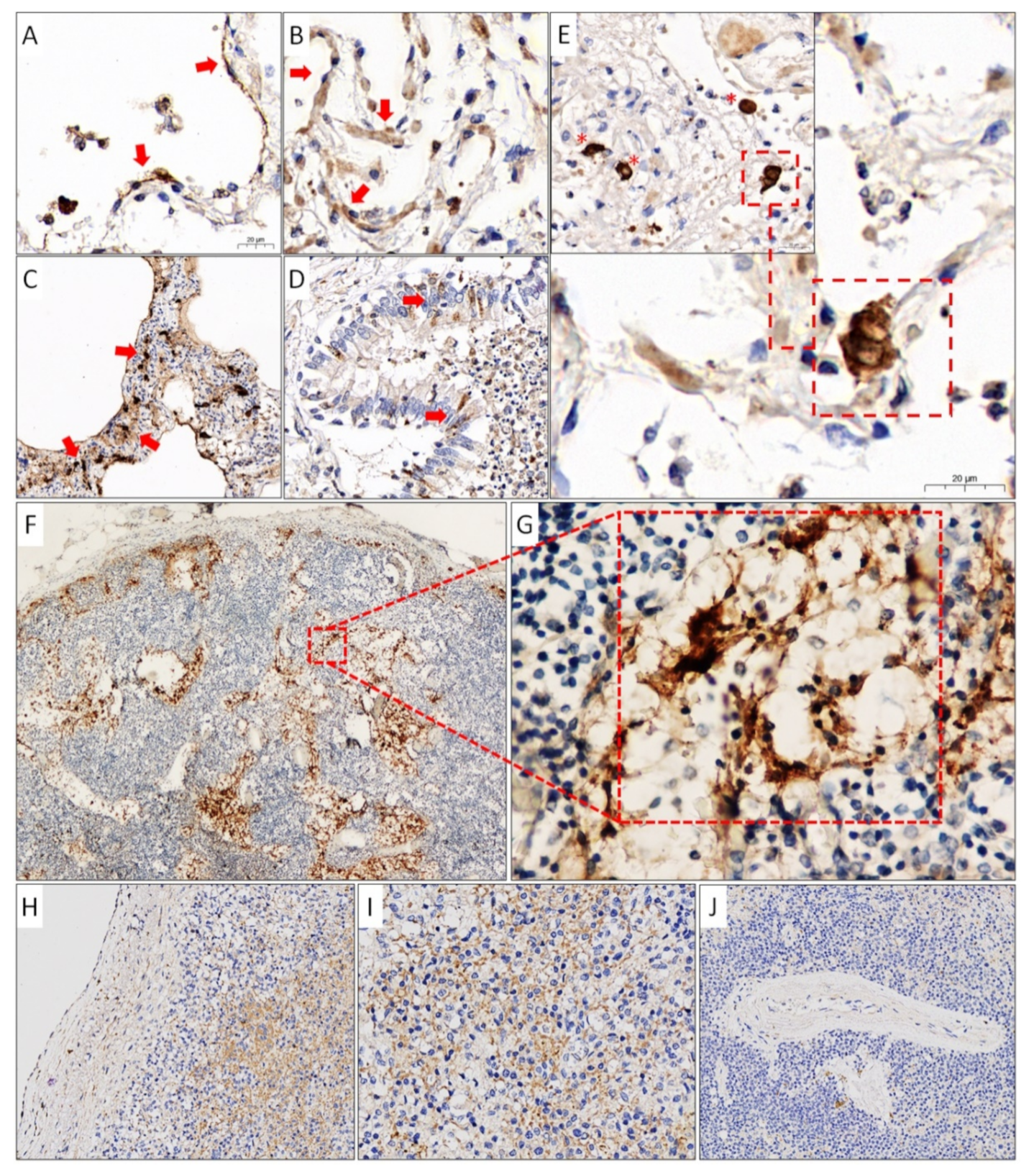
2.5. Immunohistochemistry
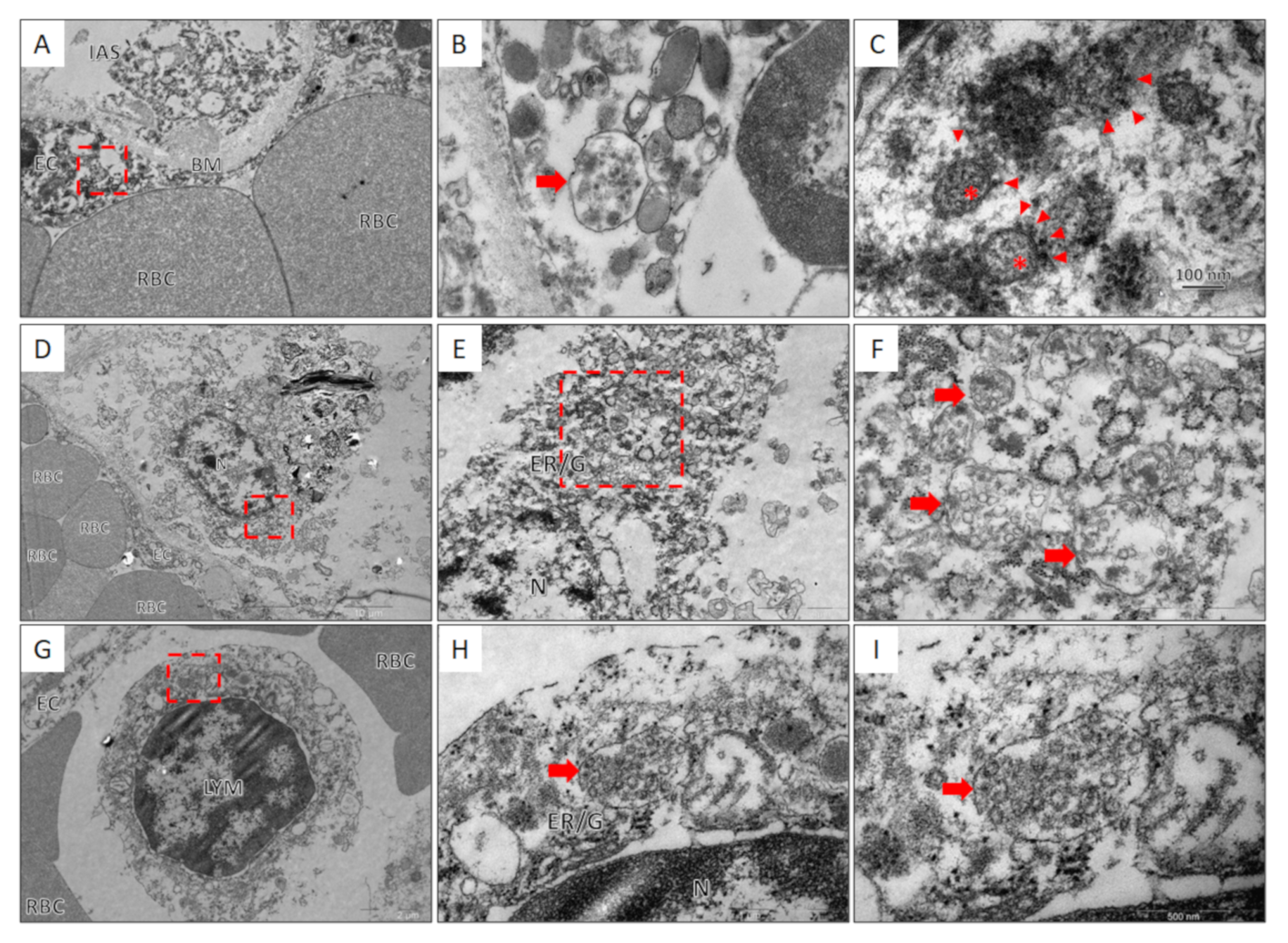
2.6. Transmission Electron Microscopy
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, E.; Du, H.; Gardner, L. An Interactive Web-Based Dashboard to Track COVID-19 in Real Time. Lancet Infect. Dis. 2020, 20, 533–534. [Google Scholar] [CrossRef]
- Khandait, H.; Gandotra, G.; Sachdeva, S.; Kramer, C.A.; Nye, D.; Golamari, R.; Jain, R. COVID-19 and Hematology-What Do We Know So Far? SN Compr. Clin. Med. 2020, 2, 2631–2636. [Google Scholar] [CrossRef] [PubMed]
- Terpos, E.; Ntanasis-Stathopoulos, I.; Elalamy, I.; Kastritis, E.; Sergentanis, T.N.; Politou, M.; Psaltopoulou, T.; Gerotziafas, G.; Dimopoulos, M.A. Hematological Findings and Complications of COVID-19. Am. J. Hematol. 2020, 95, 834–847. [Google Scholar] [CrossRef] [Green Version]
- Miesbach, W.; Makris, M. COVID-19: Coagulopathy, Risk of Thrombosis, and the Rationale for Anticoagulation. Clin. Appl. Thromb. Hemost. 2020, 26, 1076029620938149. [Google Scholar] [CrossRef]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.; Pranata, R. Lymphopenia in Severe Coronavirus Disease-2019 (COVID-19): Systematic Review and Meta-Analysis. J. Intensive Care 2020, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Jafarzadeh, S.; Nozari, P.; Mokhtari, P.; Nemati, M. Lymphopenia an Important Immunological Abnormality in Patients with COVID-19: Possible Mechanisms. Scand. J. Immunol. 2021, 93, e12967. [Google Scholar] [CrossRef] [PubMed]
- Rahi, M.S.; Jindal, V.; Reyes, S.-P.; Gunasekaran, K.; Gupta, R.; Jaiyesimi, I. Hematologic Disorders Associated with COVID-19: A Review. Ann. Hematol. 2021, 100, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Feng, Z.; Diao, B.; Tu, C.; Qiao, Q.; Yang, H.; Zhang, Y.; Wang, G.; Wang, H.; Wang, C.; et al. SARS-CoV-2 Induces Lymphocytopenia by Promoting Inflammation and Decimates Secondary Lymphoid Organs. Front. Immunol. 2021, 12, 661052. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Diao, B.; Wang, R.; Wang, G.; Wang, C.; Tan, Y.; Liu, L.; Wang, C.; Liu, Y.; Liu, Y.; et al. The Novel Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Directly Decimates Human Spleens and Lymph Nodes. Available online: https://www.medrxiv.org/content/10.1101/2020.03.27.20045427v1 (accessed on 19 May 2021).
- Sekulic, M.; Harper, H.; Nezami, B.G.; Shen, D.L.; Sekulic, S.P.; Koeth, A.T.; Harding, C.V.; Gilmore, H.; Sadri, N. Molecular Detection of SARS-CoV-2 Infection in FFPE Samples and Histopathologic Findings in Fatal SARS-CoV-2 Cases. Am. J. Clin. Pathol. 2020, 154, 190–200. [Google Scholar] [CrossRef]
- Bradley, B.T.; Maioli, H.; Johnston, R.; Chaudhry, I.; Fink, S.L.; Xu, H.; Najafian, B.; Deutsch, G.; Lacy, J.M.; Williams, T.; et al. Histopathology and Ultrastructural Findings of Fatal COVID-19 Infections in Washington State: A Case Series. Lancet 2020, 396, 320–332. [Google Scholar] [CrossRef]
- Dorward, D.A.; Russell, C.D.; Um, I.H.; Elshani, M.; Armstrong, S.D.; Penrice-Randal, R.; Millar, T.; Lerpiniere, C.E.B.; Tagliavini, G.; Hartley, C.S.; et al. Tissue-Specific Immunopathology in Fatal COVID-19. Am. J. Respir. Crit. Care Med. 2021, 203, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xie, J.; Zhao, L.; Fei, X.; Zhang, H.; Tan, Y.; Nie, X.; Zhou, L.; Liu, Z.; Ren, Y.; et al. Alveolar Macrophage Dysfunction and Cytokine Storm in the Pathogenesis of Two Severe COVID-19 Patients. EBioMedicine 2020, 57, 102833. [Google Scholar] [CrossRef] [PubMed]
- Prilutskiy, A.; Kritselis, M.; Shevtsov, A.; Yambayev, I.; Vadlamudi, C.; Zhao, Q.; Kataria, Y.; Sarosiek, S.R.; Lerner, A.; Sloan, J.M.; et al. SARS-CoV-2 Infection-Associated Hemophagocytic Lymphohistiocytosis. Am. J. Clin. Pathol. 2020, 154, 466–474. [Google Scholar] [CrossRef]
- Yan, L.; Mir, M.; Sanchez, P.; Beg, M.; Peters, J.; Enriquez, O.; Gilbert, A. COVID-19 in a Hispanic Woman. Arch. Pathol. Lab. Med. 2020, 144, 1041–1047. [Google Scholar] [CrossRef]
- Lax, S.F.; Skok, K.; Zechner, P.; Kessler, H.H.; Kaufmann, N.; Koelblinger, C.; Vander, K.; Bargfrieder, U.; Trauner, M. Pulmonary Arterial Thrombosis in COVID-19 with Fatal Outcome: Results from a Prospective, Single-Center, Clinicopathologic Case Series. Ann. Intern. Med. 2020, 173, 350–361. [Google Scholar] [CrossRef]
- Bao, C.; Liu, X.; Zhang, H.; Li, Y.; Liu, J. Coronavirus Disease 2019 (COVID-19) CT Findings: A Systematic Review and Meta-Analysis. J. Am. Coll. Radiol. 2020, 17, 701–709. [Google Scholar] [CrossRef]
- Zhu, J.; Zhong, Z.; Li, H.; Ji, P.; Pang, J.; Li, B.; Zhang, J. CT Imaging Features of 4121 Patients with COVID-19: A Meta-Analysis. J. Med. Virol. 2020, 92, 891–902. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Han, X.; Jiang, N.; Cao, Y.; Alwalid, O.; Gu, J.; Fan, Y.; Zheng, C. Radiological Findings from 81 Patients with COVID-19 Pneumonia in Wuhan, China: A Descriptive Study. Lancet Infect. Dis. 2020, 20, 425–434. [Google Scholar] [CrossRef]
- Valette, X.; du Cheyron, D.; Goursaud, S. Mediastinal Lymphadenopathy in Patients with Severe COVID-19. Lancet Infect. Dis. 2020, 20, 1230. [Google Scholar] [CrossRef]
- Haslbauer, J.D.; Matter, M.S.; Stalder, A.K.; Tzankov, A. Histomorphological Patterns of Regional Lymph Nodes in COVID-19 Lungs. Pathologe 2021, 1–9. [Google Scholar] [CrossRef]
- Martines, R.B.; Ritter, J.M.; Matkovic, E.; Gary, J.; Bollweg, B.C.; Bullock, H.; Goldsmith, C.S.; Silva-Flannery, L.; Seixas, J.N.; Reagan-Steiner, S.; et al. Pathology and Pathogenesis of SARS-CoV-2 Associated with Fatal Coronavirus Disease, United States. Emerg. Infect. Dis. 2020, 26, 2005–2015. [Google Scholar] [CrossRef]
- Xu, X.; Chang, X.N.; Pan, H.X.; Su, H.; Huang, B.; Yang, M.; Luo, D.J.; Weng, M.X.; Ma, L.; Nie, X. Pathological changes of the spleen in ten patients with coronavirus disease 2019(COVID-19) by postmortem needle autopsy. Zhonghua Bing Li Xue Za Zhi 2020, 49, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.H.; Li, T.Y.; He, Z.C.; Ping, Y.F.; Liu, H.W.; Yu, S.C.; Mou, H.M.; Wang, L.H.; Zhang, H.R.; Fu, W.J.; et al. A pathological report of three COVID-19 cases by minimal invasive autopsies. Zhonghua Bing Li Xue Za Zhi. 2020, 49, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Bugra, A.; Das, T.; Arslan, M.N.; Ziyade, N.; Buyuk, Y. Postmortem Pathological Changes in Extrapulmonary Organs in SARS-CoV-2 Rt-PCR-Positive Cases: A Single-Center Experience. Ir. J. Med. Sci. 2021, 1–11. [Google Scholar] [CrossRef]
- Chu, H.; Zhou, J.; Wong, B.H.-Y.; Li, C.; Chan, J.F.-W.; Cheng, Z.-S.; Yang, D.; Wang, D.; Lee, A.C.-Y.; Li, C.; et al. Middle East Respiratory Syndrome Coronavirus Efficiently Infects Human Primary T Lymphocytes and Activates the Extrinsic and Intrinsic Apoptosis Pathways. J. Infect. Dis. 2016, 213, 904–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Gong, E.; Zhang, B.; Zheng, J.; Gao, Z.; Zhong, Y.; Zou, W.; Zhan, J.; Wang, S.; Xie, Z.; et al. Multiple Organ Infection and the Pathogenesis of SARS. J. Exp. Med. 2005, 202, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chu, H.; Chan, J.F.-W.; Yuen, K.-Y. Middle East Respiratory Syndrome Coronavirus Infection: Virus-Host Cell Interactions and Implications on Pathogenesis. Virol. J. 2015, 12, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borsa, M.; Mazet, J.M. Attacking the Defence: SARS-CoV-2 Can Infect Immune Cells. Nat. Rev. Immunol. 2020, 20, 592. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Liu, Y.; Cao, L.; Wang, D.; Guo, M.; Jiang, A.; Guo, D.; Hu, W.; Yang, J.; Tang, Z.; et al. Transcriptomic Characteristics of Bronchoalveolar Lavage Fluid and Peripheral Blood Mononuclear Cells in COVID-19 Patients. Emerg. Microbes Infect. 2020, 9, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Taghiloo, S.; Aliyali, M.; Abedi, S.; Mehravaran, H.; Sharifpour, A.; Zaboli, E.; Eslami-Jouybari, M.; Ghasemian, R.; Vahedi-Larijani, L.; Hossein-Nattaj, H.; et al. Apoptosis and Immunophenotyping of Peripheral Blood Lymphocytes in Iranian COVID-19 Patients: Clinical and Laboratory Characteristics. J. Med. Virol. 2021, 93, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Park, S.-S.; Kim, T.Y.; Lee, D.-G.; Kim, D.-W. Lymphopenia as a Biological Predictor of Outcomes in COVID-19 Patients: A Nationwide Cohort Study. Cancers 2021, 13, 471. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, D.; Peaper, D.R.; She, R.C.; Nolte, F.S.; Wojewoda, C.M.; Anderson, N.W.; Pritt, B.S. College of American Pathologists (CAP) Microbiology Committee Perspective: Caution Must Be Used in Interpreting the Cycle Threshold (Ct) Value. Clin. Infect. Dis. 2021, 72, e685–e686. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, R.; Aherfi, S.; Wurtz, N.; Grimaldier, C.; Van Hoang, T.; Colson, P.; Raoult, D.; La Scola, B. Correlation Between 3790 Quantitative Polymerase Chain Reaction-Positives Samples and Positive Cell Cultures, Including 1941 Severe Acute Respiratory Syndrome Coronavirus 2 Isolates. Clin. Infect. Dis. 2021, 72, e921. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.; Hirsch, J.S.; Narasimhan, M.; Crawford, J.M.; McGinn, T.; Davidson, K.W.; Barnaby, D.P.; Becker, L.B.; Chelico, J.D.; Cohen, S.L.; et al. Presenting Characteristics, Comorbidities, and Outcomes Among 5700 Patients Hospitalized with COVID-19 in the New York City Area. JAMA 2020, 323, 2052–2059. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.H.; Bayat Mokhtari, E.; Mudvari, P.; Stein, S.; Stringham, C.D.; Wagner, D.; Ramelli, S.; Ramos-Benitez, M.J.; Strich, J.R.; Davey, R.T.J.; et al. High-Throughput, Single-Copy Sequencing Reveals SARS-CoV-2 Spike Variants Coincident with Mounting Humoral Immunity during Acute COVID-19. PLoS. Pathog. 2021, 17, e1009431. [Google Scholar] [CrossRef]
- Kuipers, J.; Batavia, A.A.; Jablonski, K.P.; Bayer, F.; Borgsmüller, N.; Dondi, A.; Drăgan, M.-A.; Ferreira, P.; Jahn, K.; Lamberti, L.; et al. Within-Patient Genetic Diversity of SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Liu, S.; Shen, J.; Fang, S.; Li, K.; Liu, J.; Yang, L.; Hu, C.-D.; Wan, J. Genetic Spectrum and Distinct Evolution Patterns of SARS-CoV-2. Front. Microbiol. 2020, 11, 593548. [Google Scholar] [CrossRef]
- Buchrieser, J.; Dufloo, J.; Hubert, M.; Monel, B.; Planas, D.; Rajah, M.M.; Planchais, C.; Porrot, F.; Guivel-Benhassine, F.; Van der Werf, S.; et al. Syncytia Formation by SARS-CoV-2-Infected Cells. EMBO J. 2020, 39, e106267. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, C.S.; Miller, S.E.; Martines, R.B.; Bullock, H.A.; Zaki, S.R. Electron Microscopy of SARS-CoV-2: A Challenging Task. Lancet 2020, 395, e99. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Liu, H.; Li, W.; Lin, F.; Jiang, L.; Li, X.; Xu, P.; Zhang, L.; Zhao, L.; et al. SARS-CoV-2 Infection of the Liver Directly Contributes to Hepatic Impairment in Patients with COVID-19. J. Hepatol. 2020, 73, 807–816. [Google Scholar] [CrossRef]
- Turoňová, B.; Sikora, M.; Schürmann, C.; Hagen, W.J.H.; Welsch, S.; Blanc, F.E.C.; von Bülow, S.; Gecht, M.; Bagola, K.; Hörner, C.; et al. In Situ Structural Analysis of SARS-CoV-2 Spike Reveals Flexibility Mediated by Three Hinges. Science 2020, 370, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial Cell Infection and Endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Garrido Ruiz, M.C.; Santos-Briz, Á.; Santos-Briz, Á.; Sánchez, A.; Alonso-Riaño, M.; Burgos, J.; Medina-Miguelañez, M.; Puebla, L.; Román-Curto, C.; Roncero-Riesco, M.; et al. Spectrum of Clinicopathologic Findings in COVID-19-Induced Skin Lesions: Demonstration of Direct Viral Infection of the Endothelial Cells. Am. J. Surg. Pathol. 2021, 45, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Colmenero, I.; Santonja, C.; Alonso-Riaño, M.; Noguera-Morel, L.; Hernández-Martín, A.; Andina, D.; Wiesner, T.; Rodríguez-Peralto, J.L.; Requena, L.; Torrelo, A. SARS-CoV-2 Endothelial Infection Causes COVID-19 Chilblains: Histopathological, Immunohistochemical and Ultrastructural Study of Seven Paediatric Cases. Br. J. Dermatol. 2020, 183, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Lei, T.; Patel, P.S.; Lee, C.H.; Monaghan-Nichols, P.; Xin, H.-B.; Qiu, J.; Fu, M. Direct Activation of Endothelial Cells by SARS-CoV-2 Nucleocapsid Protein Is Blocked by Simvastatin. bioRxiv. 2021. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Sex | Age | Time 1 | LYM Count 2 | Phase of DAD 3/Other Pulmonary Findings | XVL in the Lungs 4 | VL in LN 5 | VL in Spleen 6 |
|---|---|---|---|---|---|---|---|---|
| Group 1: (n = 3) SARS-CoV-2 has not been detected | ||||||||
| 7 | М | 62 | 2 | 0.9 | Proliferative | 0 | 0 | 0 |
| 17 | F | 77 | 15 | 1.3 | Proliferative/emphysema | 0 | 0 | 0 |
| 23 | М | 90 | 9 | 2.4 | Bronchopneumonia with hemorrhages and fibrosis | 0 | 0 | 0 |
| Group 2: (n = 12) SARS-CoV-2 dissemination limited to lungs | ||||||||
| 5 | F | 73 | 18 | 4 | Proliferative/bronchopneumonia with hemorrhages | 18 | 0 | 0 |
| 34 | F | 73 | 23 | 1.3 | Bronchopneumonia | 27 | 0 | 0 |
| 29 | F | 79 | 24 | 0.7 | Bronchopneumonia | 38 | 0 | 0 |
| 22 | М | 90 | 10 | 1.9 | Proliferative | 63 | 0 | 0 |
| 15 | М | 68 | 19 | 0.9 | Bronchopneumonia | 190 | 0 | 0 |
| 31 | F | 78 | 10 | 0.8 | Proliferative | 208 | 0 | 0 |
| 20 | F | 85 | 23 | 0.4 | Bronchopneumonia | 270 | 0 | 0 |
| 6 | М | 66 | 27 | 0.3 | Hemorrhages and fibrosis | 313 | 0 | 0 |
| 8 | М | 64 | 19 | 1.2 | Proliferative | 677 | 0 | 0 |
| 32 | М | 85 | 10 | 1.6 | Proliferative | 694 | 0 | 0 |
| 11 | М | 84 | 17 | 0.7 | Proliferative | 706 | - | n/a |
| 14 | М | 80 | 10 | 0.6 | Proliferative | 1952 | 0 | 0 |
| Group 3: (n = 21) SARS-CoV-2 dissemination to lungs, lymph nodes, and spleen | ||||||||
| 33 | F | 86 | 3 | 0.6 | Proliferative | 810 | 96 | 0 |
| 13 | F | 71 | 1 | 0.7 | Exudative | 834 | 1318 | 629 |
| 35 | F | 73 | 2 | 1.4 | Proliferative | 910 | 215 | 197 |
| 21 | М | 67 | 13 | 0.4 | Proliferative | 982 | 180 | 0 |
| 30 | М | 61 | 5 | 1.3 | Proliferative/bronchopneumonia | 1002 | 237 | 0 |
| 1 | М | 52 | 1 | 2.7 | Proliferative/emphysema | 1016 | 112 | 0 |
| 25 | М | 74 | 22 | 0.5 | Proliferative | 1032 | 825 | 0 |
| 16 | М | 67 | 9 | 0.7 | Proliferative/bronchopneumonia with hemorrhages | 2657 | 1110 | 0 |
| 4 | М | 76 | 5 | 0.8 | Exudative and early proliferative/hemorrhages | 4909 | 195 | 71 |
| 19 | М | 84 | 24 | 0.5 | Exudative | 7476 | 1807 | 0 |
| 9 | F | 72 | 12 | 0.6 | Exudative/bronchopneumonia | 12,116 | 3801 | - |
| 28 | F | 95 | 7 | 0.4 | Exudative | 14,522 | 839 | 34 |
| 3 | М | 84 | 11 | 1.2 | Exudative and early proliferative/bronchopneumonia with necrosis | 14,937 | 162 | 9 |
| 27 | М | 84 | 16 | 0.3 | Exudative | 16,867 | 1180 | 1320 |
| 18 | М | 85 | 16 | 0.5 | Exudative | 17,817 | n/a | 218 |
| 36 | М | 73 | 2 | 0.6 | Exudative | 18,219 | 2691 | 181 |
| 26 | F | 93 | 7 | 0.9 | Exudative | 27,349 | 1629 | 213 |
| 2 | F | 86 | 25 | 1 | Exudative | 73,214 | 789 | 72 |
| 12 | F | 85 | 7 | 0.9 | Exudative | 151,183 | 513 | 1891 |
| 24 | F | 76 | 11 | 2.7 | Exudative | 159,217 | 11,586 | 551 |
| 10 | М | 61 | 6 | 0.4 | Exudative | 250,281 | 5958 | 2899 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdullaev, A.; Odilov, A.; Ershler, M.; Volkov, A.; Lipina, T.; Gasanova, T.; Lebedin, Y.; Babichenko, I.; Sudarikov, A. Viral Load and Patterns of SARS-CoV-2 Dissemination to the Lungs, Mediastinal Lymph Nodes, and Spleen of Patients with COVID-19 Associated Lymphopenia. Viruses 2021, 13, 1410. https://doi.org/10.3390/v13071410
Abdullaev A, Odilov A, Ershler M, Volkov A, Lipina T, Gasanova T, Lebedin Y, Babichenko I, Sudarikov A. Viral Load and Patterns of SARS-CoV-2 Dissemination to the Lungs, Mediastinal Lymph Nodes, and Spleen of Patients with COVID-19 Associated Lymphopenia. Viruses. 2021; 13(7):1410. https://doi.org/10.3390/v13071410
Chicago/Turabian StyleAbdullaev, Adhamjon, Akmaljon Odilov, Maxim Ershler, Alexey Volkov, Tatiana Lipina, Tatiana Gasanova, Yuri Lebedin, Igor Babichenko, and Andrey Sudarikov. 2021. "Viral Load and Patterns of SARS-CoV-2 Dissemination to the Lungs, Mediastinal Lymph Nodes, and Spleen of Patients with COVID-19 Associated Lymphopenia" Viruses 13, no. 7: 1410. https://doi.org/10.3390/v13071410
APA StyleAbdullaev, A., Odilov, A., Ershler, M., Volkov, A., Lipina, T., Gasanova, T., Lebedin, Y., Babichenko, I., & Sudarikov, A. (2021). Viral Load and Patterns of SARS-CoV-2 Dissemination to the Lungs, Mediastinal Lymph Nodes, and Spleen of Patients with COVID-19 Associated Lymphopenia. Viruses, 13(7), 1410. https://doi.org/10.3390/v13071410