Abstract
Flaviviruses comprise several important human pathogens which cause significant morbidity and mortality worldwide. Like any other virus, they are obligate intracellular parasites. Therefore, studying the host cellular factors that promote or restrict their replication and pathogenesis becomes vital. Since inhibiting the host dependency factors or activating the host restriction factors can suppress the viral replication and propagation in the cell, identifying them reveals potential targets for antiviral therapeutics. Clustered regularly interspaced short palindromic repeats (CRISPR) technology has provided an effective means of producing customizable genetic modifications and performing forward genetic screens in a broad spectrum of cell types and organisms. The ease, rapidity, and high reproducibility of CRISPR technology have made it an excellent tool for carrying out genome-wide screens to identify and characterize viral host dependency factors systematically. Here, we review the insights from various Genome-wide CRISPR screens that have advanced our understanding of Flavivirus-Host interactions.
1. Introduction
The family Flaviviridae constitutes many enveloped single-stranded positive-sense RNA viruses. This RNA genome encodes a single open reading frame that is translated at the Endoplasmic Reticulum (ER) to give rise to a polyprotein, which is subsequently cleaved by viral and host cell proteases. This processing forms ten functional proteins, including the three structural proteins, Capsid (C), Pre-membrane (prM), and Envelope (E), and seven non-structural proteins, which include NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5. The family Flaviviridae consists of three viral genera, Flavivirus, Pestivirus, and Hepatitis C virus (HCV), with a total of more than 70 viruses, most of which have their polyproteins organized in a similar way. NS1 has a role in replication [1], as well as innate immune evasion by interfering with the Toll-Like Receptor (TLR) signaling pathway [2]. NS2A has a direct role in the replication of viral RNA. NS3 serves as the helicase and also interacts with NS2B as a cofactor (NS2B3) to form the viral protease [3,4]. NS4B inhibits the interferon (IFN)-dependent signaling pathway [5]. NS5 is both the RNA-dependent RNA-polymerase, which catalyzes genome replication and methyltransferase, which caps the nascent RNA genomes [6,7]. In addition to these, the viral RNA genome also contains 3′ and 5′ untranslated regions (UTRs) that contribute to genome stability and translation [8]. Flavivirus genome replication occurs within ER membrane involuted structures called virus replication compartments (RCs) [9]. These remodeled ER substructures assist in concentrating the replication substrates and shielding the viral RNAs from detection by the host immune system. NS4A protein is the key organizer of these ER membrane structures [10]. Within these RCs, the NS5 protein, along with other viral and human host factors, performs the enzymatic steps of genomic viral RNA replication [11]. The components then assemble and bud off from the ER through the trans-Golgi pathway and are trafficked out of the cell via exocytosis. The maturation step is associated with the cleavage of prM by viral and host furin-like proteases. Some features that differ among the three genera include the existence of an additional cleavage site in the NS5 region of HCV and pestiviruses, but not flaviviruses. This site separates the N-terminal portion (NS5A) from the viral polymerase (NS5B) [12].
There are 53 defined species in the genus Flavivirus. Some medically important flaviviruses are associated with a spectrum of diseases ranging from mild self-limiting febrile illness to severe life-threatening encephalitis, hepatitis, vascular shock syndrome, congenital abnormalities, and Viral hemorrhagic fevers (VHFs). These include Dengue (DENV), Japanese Encephalitis (JEV), West Nile (WNV), Zika (ZIKV), and yellow Fever (YFV) viruses. Currently, DENV fever is the most prevalent arthropod-borne viral disease globally, with an estimated 390 million total infections, 100 million clinically apparent cases, and 500,000 presentations of severe dengue per year worldwide, with at least 2.5 billion people at risk [13]. ZIKV has been correlated with severe congenital abnormalities such as microcephaly and other birth defects in unborn children due to potential ZIKV exposure to the mother during pregnancy [14]. JEV, which primarily affects children, is estimated to cause approximately 14,000 to 20,000 fatal cases annually [15]. WNV virus is maintained in nature in a mosquito–bird–mosquito transmission cycle but has spilled over and caused disease in humans and horses. The spread of WNV and JEV across different geographical locations has been attributed to the seasonal migrations of birds [16].
Most of the viruses in the genus Flavivirus survive in nature by replicating alternately in a vertebrate host and a hematophagous arthropod (mosquitoes and ticks) and hence are classified as arboviruses. These arthropod vectors acquire the virus by biting a viremic host. The virus then replicates in the vector’s tissues. The transmission to another vertebrate host happens through salivary secretions of these arthropod vectors as they take up blood meal. The virus then multiplies within the vertebrate host, causing viremia and disease. Non-human primates, mostly wild mammals and birds form the principal vertebrate hosts for most flaviviruses, thus leading to the sylvatic transmission cycle [17]. The natural zoonotic cycle of these viruses does not usually involve humans. However, a few viruses can jump into a human–mosquito–human cycle. This is called the urban cycle, where humans catch the disease when they encroach on forest habitat and are bitten by the arbovirus-infected mosquitoes. When these people move to densely populated urban settings, such infections are transmitted by highly anthropophilic urban mosquitoes and can lead to explosive outbreaks. This is when a sylvatic transmission cycle is said to have ‘spilled over’ into an urban transmission cycle. An excellent example is the emergence of sylvatic YFV during an epidemic in the Gambia [18,19]. Thus, because of their high transmission potential, these viruses can cause severe and widespread outbreaks in many tropical and subtropical regions of the world, depending upon the presence of appropriate insect vectors.
Since most of the flaviviruses are vertically transmitted in the arthropods, they may elicit long-term persistence and potential re-emergence. Therefore, improving or coming up with novel strategies to blunt flavivirus disease is of the essence, even if better vaccines or antivirals become available. The cellular interactions of flaviviruses with their human or mosquito hosts are critical for manifesting the viral life cycle and decoding this information can be very useful for engineering novel strategies to disrupt the disease or its transmission. Genome-wide CRISPR knockout (GeCKO) screens offer an exceptional approach to identifying such host factors. The revelation and validation of these host dependency factors can be followed by discovering or designing small molecule drugs to suppress them. It is important to note that therapeutic intervention of host factors, rather than viral proteins, is associated with a much higher barrier to drug resistance. In recent years, GeCKO screens have successfully identified host dependency factors of several clinically relevant viruses other than Flaviviruses [20]. These include Influenza A [21,22], Coronaviruses [23,24], picornaviruses [25], and HIV [26]. Moreover, understanding these interactions has paved the way for developing efficient therapeutics and preventive strategies against these infections [27,28].
This review gives a brief overview of GeCKO Technology and summarizes various studies that have utilized it to identify host dependency and restriction factors for Flavivirus Infection.
2. CRISPR-Cas Biology: An Overview
The clustered regularly interspaced short palindromic repeats (CRISPR)-Cas system was initially discovered as a sophisticated acquired immune system in Prokaryotes and Archaea, where it acts to protect against invading bacteriophages and conjugative plasmids by degrading their genetic elements [29,30]. At the molecular level, there are two key players in this process: the CRISPR RNA (crRNA) and its associated endonuclease (Cas) [31]. CRISPR systems have been rapidly implemented to perform eukaryotic genome manipulation. As seen in their natural prokaryotic counterparts, the engineered CRISPR systems also have two effector molecules: a single guide RNA (sgRNA) chimera, consisting of a fusion of a crRNA and tracrRNA, and a Cas protein. The sgRNA comprises a customizable spacer sequence of ∼20 nucleotides and a scaffold sequence that binds to the Cas protein. The spacer can be designed according to the genomic target to be modified. Therefore, by merely altering the spacer sequence, the sgRNA-Cas Ribonucleoprotein complex can be directed to target any genetic loci [32,33]. For knocking out a gene, the sgRNA designed specific to the targeted gene and a Cas enzyme are made to co-express within the chosen cell type or organism. Upon binding to the target, Cas9 undergoes a conformational change and cleaves the opposite strands of the target DNA, creating a blunt-ended Double-strand break (DSB). When cellular repair mechanisms try to ligate the broken ends, they end up introducing small nucleotide insertions or deletions (indels) or frameshift mutations, leading to premature stop codons within the ORF of the target. This ultimately leads to a loss-of-function mutation within the targeted gene.
One can use the same sgRNA library combined with certain derivatives of Cas9 for programmable genetic manipulations [34].
- (1)
- A catalytically dead Cas9 enzyme (dCas9) for CRISPR interference (CRISPRi) studies: This dCas9 binds to the target DNA sequence guided by the gRNA. Instead of cleaving the bound DNA, the dCas9 enzyme remains bound to the target DNA sequence, disrupting RNA polymerase or transcription factor binding to the promoter. Other than steric hindrance, CRISPRi can also repress transcription via a repressor domain, such as the Krüppel associated box (KRAB), fused to dCas9 [35,36] (Figure 1).
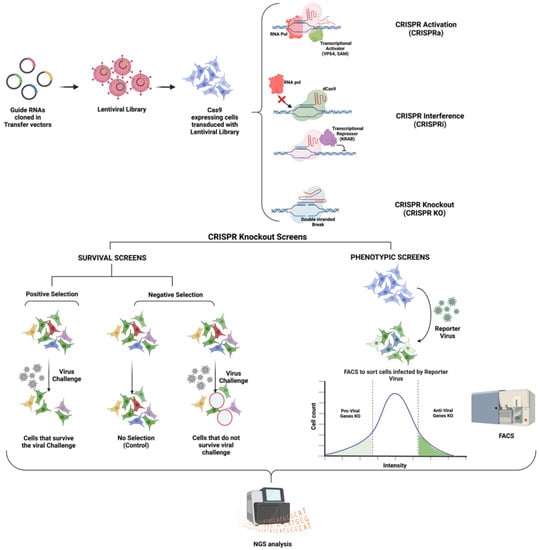 Figure 1. Schematic of different types of CRISPR Screens to identify virus-host interactions (Created with BioRender.com).
Figure 1. Schematic of different types of CRISPR Screens to identify virus-host interactions (Created with BioRender.com). - (2)
- Cas9 tethered with a transcriptional activator such as SunTAG [33], Synergistic Activation Mediator (SAM) [37], VP64 [38], etc., for CRISPR activation (CRISPRa) studies: Such Cas9 leads to the recruitment of transcriptional machinery to the targeted promoter. CRISPRa studies are employed to perform gain-of-function studies [35,39].
In general, the idea of a genome-wide loss-of-function screening is to generate a large population of cells with a wide variety of genes knocked out or knocked down and then identify the genetic perturbations that lead to the desired phenotype, such as the survival of cells despite a lethal viral infection [20]. Presently, the most popular method for conducting GeCKO screens involves using pooled lentiviral CRISPR libraries, which is briefly elucidated in the next section and illustrated in Figure 1.
3. CRISPR Screens for Studying Flavivirus Infections
Viruses, being obligate intracellular parasites, are intimately associated with their host cells [52]. While dual-host flaviviruses can cause disease in vertebrates as well as arthropods, insect-specific flaviviruses are restricted to their competent arthropods. How flaviviruses establish persistent infection in their insect vectors and humans relies upon an intricate interplay between flavivirus-encoded immune antagonists and the host antiviral innate immune effectors. In general, flaviviruses have a conserved replication cycle, which includes the following steps: viral entry via receptor-mediated endocytosis, fusion with the endosomal membrane and release of viral RNA, genome replication and translation into proteins in the ER membrane structures, virion packaging and processing through the trans-Golgi secretory pathway, and release of viruses via exocytosis. At every step, flaviviruses rely on the host machinery to facilitate replication, dampen host immune response, or disrupt cellular processes to aid pathogenesis. These host factors include proteins, RNAs, lipids, carbohydrates, or small molecules. They can be recognized by making use of techniques that probe direct or indirect physical interactions with viral RNA or proteins or through genetic interactions by perturbing the host, as in CRISPR and RNAi screens.
As mentioned before, CRISPR screens provide a tremendous advantage for high-throughput analysis of viral and host factors. Moreover, CRISPR-Cas technology is reliable for validating candidate genes. In contrast to gene knockdown approaches, such as RNAi, gene knockouts are absolute. They are associated with lesser variations when virus replication is quantified using assays such as qPCR, plaque assays, or immunostaining. Various genetic screens have been attempted over the last two decades to determine the host dependency and antiviral factors for multiple flaviviruses (Table 1).

Table 1.
Summary of Genome-Wide Genetic Screens to uncover host dependency and restriction factors for flaviviruses.
To probe for common genetic factors enriched among various CRISPR KO screens, we analyzed the overlapping genes from the top 10 hits from nine different screens (carried out for DENV, ZIKV, WNV, and JEV) and checked for intersections (Figure 2).
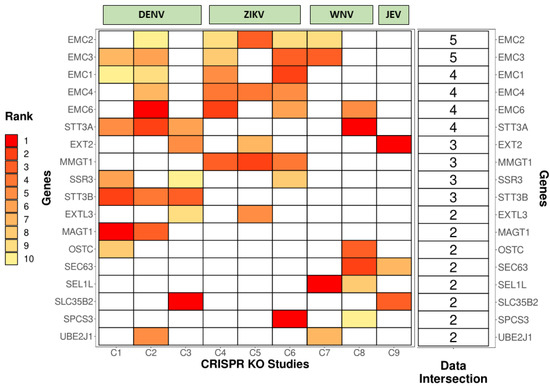
Figure 2.
Intersections among different CRISPR KO studies for Dengue Virus (DENV), Zika Virus (ZIKV), West Nile Virus (WNV), and Japanese Encephalitis Virus (JEV). The heat map shows the overlapping genes among the top 10 hits from 9 different CRISPR studies (C1 to C9) and their ranks based on the phenotypic significance (with 1 (Red) being the best to 10 (Yellow) being the last). Boxes colored in white denote that the gene is not present in the filtered dataset. The last column represents the number of studies in which the gene is present. Studies used in the analysis: C1 [53], C2 [54], C3 [55], C4 [56], C5 [57], C6 [58], C7 [59], C8 [60], C9 [61].
Several ER-associated proteins involved in membrane remodeling, protein stabilization, folding, and degradation emerged as common genetic factors for various viruses. Some of the important viral–host interactions uncovered by these studies are discussed in detail below:
3.1. Virus Receptors and Attachment Factors
Independent CRISPR Screens have revealed various cell surface molecules utilized by flaviviruses such as Zika, Dengue, and West Nile virus to enter the cell. These include heparan sulfate proteoglycans (HSPG) and TAM (e.g., Tyro3, Axl, and Mer) family receptor tyrosine kinases that interact with the envelope protein of the virus [58]. Genes associated with heparin sulfation (NDST1 and EXT1) have also been identified [58]. A recent CRISPR KO study performed in glioblastoma stem cells revealed integrin αvβ5 as an internalization factor for ZIKV. The authors further demonstrated that using an αvβ5 blocking antibody or two chemical inhibitors (SB273005 and cilengitide) brings down the ZIKV infection and alleviates ZIKV-induced pathology in human neural stem cells and mouse brains [67]. Another study by Marceau et al. in 2016 unraveled some receptors cardinal for Hepatitis C Virus (HCV) to enter hepatocytes. These include CD81, occludin (OCLN), and claudin 1 (CLDN1) [53]. Zhao et al. carried out a CRISPR KO screen on porcine kidney cells challenged with JEV and recognized many genes associated with heparan sulfate proteoglycans (HSPGs) and their metabolism. The study highlighted many potentially vulnerable targets for developing breeding technologies to combat and prevent JEV disease in pigs [61].
3.2. Viral Translation and Insertion into ER Membrane
Post uncoating, the viral RNA is translated by host ribosomes. Several proteins have been shown to interact with the nascent polypeptide for its proper folding, insertion into the ER membrane, and further processing (Figure 3):
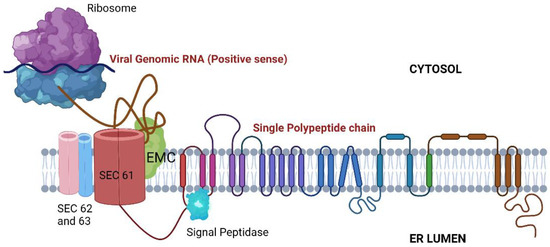
Figure 3.
ER-associated proteins are involved in viral RNA translation, stabilization, and folding. SEC61: Translocon; EMC: Endoplasmic reticulum membrane protein complex.
- Signal Peptidase Complex (SPCS): After being translated, the flavivirus polyprotein is inserted into the ER membrane as a single multipass protein and cleaved by viral and host proteases, including the host signal peptidase complex (SPCS). Knocking out SPCS1, a significant component of the SPCS, ablated the replication of all flaviviruses but not that of the unrelated RNA viruses, suggesting that it is needed for flavivirus replication specifically. Mechanistic studies revealed that the SPCS1 is involved in the cleavage of the polyprotein’s structural proteins prM and E [60].
- Translocon-associated protein complex (TRAP): The SRP ribonucleotide complex recognizes and binds to a hydrophobic transmembrane region of the nascent polypeptide, arrests translation, and brings the ribosome to a translocon where translation continues. Since the translated polyprotein contains several transmembrane domains that need to be appropriately integrated into the ER membrane, the host SRP-translocon pathway proteins such as SEC61A1 and SEC63 also showed up in several CRISPR Screens [53,60,64,68] and in an RNAi Screen [66]. Additionally, several protein-protein interaction studies have revealed interactions between ZIKV/DENV NS4A and SEC62, SEC61γ, and SRPR; NS4A/2B and SEC61β; and NS4B with SEC61α [69,70]. Interestingly, pharmacological modulation of this complex has been shown to inhibit DENV and ZIKV replication [70,71].
- Endoplasmic-reticulum-associated protein degradation (ERAD) Pathway: ERAD is a protein quality control mechanism that recognizes incorrectly folded proteins in the ER lumen. These proteins are then retro-translocated through the ER membrane to the cytosol to be targeted for proteasomal degradation. Certain components of the classical ERAD machinery, especially the ones that form the retro-translocation complex, were shown to be essential for infectious virus particle formation and virus-induced cell death for DENV, ZIKV, JEV, and WNV. These include proteins such as SEL1L, derlin 2 (DERL2), and ubiquitin-conjugating enzyme E2 J1 (UBE2J1). Knocking out these genes conferred robust protection against WNV-induced cell death. Remarkably, WNV replication was unaffected. Thus, these factors have been speculated to be the chief drivers of WNV-induced cell death [59].
- The Endoplasmic reticulum membrane protein complex (EMC): EMC is an evolutionarily conserved complex responsible for stabilizing and helping in the insertion of multipass membrane proteins in the ER. Several genetic screens have independently shown the EMC proteins to be essential for correct viral protein insertion into the ER membrane [53,58,59,68]. A 2019 study suggested that biogenesis and co-translational stabilization of DENV and ZIKV multipass proteins NS4A and NS4B rely on the interaction with EMC components [72]. The authors used a dual-fluorescence reporter system to map the hydrophobic transmembrane regions of NS4B utilized for the interaction with the EMC complex. An independent study showed a very prominent loss of replication of DENV, ZIKV, and YFV upon knocking out protein complex EMC4. Interestingly, there was no effect on the replication of WNV. The authors speculated that this difference could be because Culex mosquitoes, rather than Aedes, primarily transmit WNV. To support this vector-specific hypothesis, they also interrogated the DENV titer in Aedes mosquito midguts, which was found to be depleted post-siRNA-mediated targeting of EMC2/3/4. All in all, the study suggested that the EMC is a critical host factor utilized by Aedes-transmitted flaviviruses [73].
- Additionally, two subunits of the endoplasmic reticulum (ER) resident dolichol-phosphate mannose synthase (DPMS) complex were identified as host dependency factors for DENV and ZIKV. The DPMS complex catalyzes the synthesis of dolichol-phosphate mannose (DPM), which serves as a mannosyl donor in pathways leading to N-glycosylation, glycosylphosphatidylinositol (GPI) anchor biosynthesis, and C- or O-mannosylation of proteins in the ER lumen. This DPMS complex was shown to be required for optimal viral RNA amplification and proper glycosylation and folding of viral structural proteins prM and E [55].
3.3. Formation of Replication Complexes (RCs) and Viral RNA Synthesis
The translated viral proteins assemble in the form of a replication complex in close association with several ER-resident host protein complexes. Many CRISPR screens mapping host factors for DENV and ZIKV uncovered proteins involved in the formation of the oligosaccharyltransferase (OST) complex, the endoplasmic reticulum membrane protein complex (EMC), and components of the ER-associated protein degradation (ERAD) pathway (Figure 4).
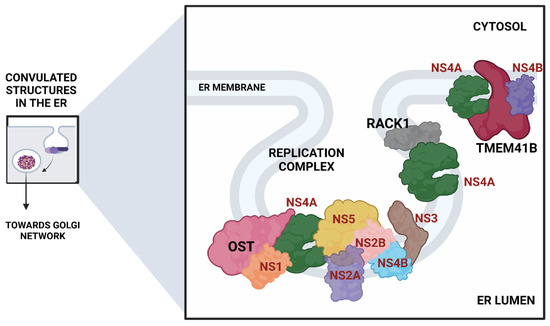
Figure 4.
ER remodeling proteins associated with the formation of the viral replication complex. (Viral proteins are shown in red).
- The Oligosaccharyltransferase (OST) complex: The OST complex is associated with N-linked glycosylation of host proteins in mammalian cells. Interestingly, different flaviviruses exhibit different dependencies on the two OST complex catalytic subunits: STT3A and STT3B. While the STT3A complex is needed for the co-translational N-linked glycosylation of the majority of the glycoproteins, the STT3B complex is essential for the co-translational or post-translational glycosylation of acceptor sites that have been skipped by the STT3A complex [74]. The OST complex was shown to be necessary for the viral RNA synthesis but not for the entry and translation. Both complexes were individually required for the replication of DENV. However, ZIKV replication was shown to be exclusively dependent on the STT3A complex, pointing out divergent virus-host interactions. Knocking out OST complex component STT3A abrogated the replication of YFV, WNV, and JEV as well. However, these replication defects were rescued by the expression of catalytically dead STT3A mutants, suggesting that the ability of OST complex to glycosylate proteins is not required for flavivirus replication. Additionally, physical interactions between flavivirus replication complex members NS1, NS2B, NS3, and NS4B and OST Complex in the ER suggest that the OST complex might act as a scaffold to orchestrate the assembly of the viral replication complex [53]. Lin et al. employed the same genome-wide CRISPR KO approach and extended this work to show that the oxidoreductase activity of the OST complex subunit MAGT1 was essential for DENV propagation. They further showed that the expression of MAGT1 depends on the presence of STT3B but not on its catalytic activity. MAGT1 was also associated with DENV NS1 and NS4B proteins during viral infection [54]. Collectively, these two studies suggested that the OST complex not only interacts physically with the replication complexes but is also engaged in post-translationally modifying and stabilizing the viral non-structural proteins associated with the complex.
- In another interesting genome-wide CRISPR KO study, Transmembrane Protein 41B (TMEM41B) was shown to be required for infection and replication of several mosquito-borne and tick-borne flaviviruses, making it a pan-flavivirus host factor. Based on mechanistic studies, the authors proposed a model whereupon flavivirus entry and subsequent translation of the viral polyprotein; this protein, TMEM41B, is recruited to the ER membrane together with viral proteins NS4A and NS4B, which are involved in inducing membrane curvature so that replication complexes (RCs) can form and make a protected environment for viral genome replication. The study also showed how the absence of TMEM41B leads to the formation of poor RCs, which ultimately causes the dsRNA replication intermediates to become exposed to innate immunity pattern recognition receptors (PRRs) in the host cell. This recognition and activation of innate immune responses lead to the abortion of the infectious replication cycle [68].
- Another significant flavivirus host factor is the Receptor for Activated C Kinase 1 (RACK1) protein. This protein has functions correlated with protein shuttling, anchoring, stabilization, and mediating specific cellular pathways through protein interactions. A recent CRISPR KO screen in Huh7 cells found that silencing of RACK1 affected the replication of several flaviviruses, including ZIKV, DENV, and WNV but not YFV. They utilized a Renilla luciferase DENV replicon to proclaim that RACK1 specifically played a role in viral genome replication rather than viral entry or translation. The authors used a replication-independent expression system to delineate the mechanism that induces the formation of RCs in the ER without virus infection. RACK1 silencing was shown to limit the organization of these structures in the ER membrane [56].
- Apart from these pathways and complexes, FAD biosynthesis, catalyzed by riboflavin (vitamin B2), kinase (RFK), and FAD synthase (FLAD1), was shown to be critical for the synthesis of HCV RNA. ELAVL1, an RNA-binding protein that binds to host mRNAs and increases their stability [75], was shown to attach to the 3′ UTR of HCV RNA to aid its replication via circularization [53]. Significantly, a protein called Cyclophilin A (CYPA) that has been shown previously to interact with HCV replication protein NS5A was also enriched (3). Some host cyclophilin inhibitors have shown promising effects in curing HCV infection in both in vitro and in vivo settings and have advanced to phase II/III clinical trials [76]. This study on cyclophilin inhibitors also highlights how targeting the host factors instead of viral factors is associated with the reduced emergence of resistance [76]. This is important because HCV exhibits a brisk mutation rate as an RNA virus, and a single mutation in the viral target can render the antiviral ineffective.
4. CRISPR Screens to Identify Anti-Flavivirus Host Factors
In addition to CRISPR KO screens, some studies have tried decoding antiviral genes using genome-wide CRISPR activation screens. Most of the high-ranking hits from one such study included interferon-stimulated genes (ISGs) such as interferon lambda 2 (IFN-λ2) and interferon alpha-inducible protein 6 (IFI6). They were shown to provide high levels of protection from the early stages of ZIKV infection. Furthermore, the identified hits were very significantly induced in ZIKV-infected placenta explants. This study is an exciting example substantiating CRISPR activation screens as a tool to decode antiviral host factors [62]. Richardson et al. used CRISPR KO screening to identify genes that regulate interferon (IFN) response to flavivirus infection [77]. In this study, the cells were treated with a high dose of IFN-α before performing a CRISPR screen to decode host antiviral factors that make cells susceptible to infection when knocked out. Several members of the IFN-α signaling pathway, namely, IFNAR1, IFNAR2, IRF9, and ISG effector gene IFN-α-inducible protein 6 (IFI6), were identified as factors with the highest antiviral activity. Further experiments showed that IFI6 prophylactically protects the uninfected cells and prevents the formation of virus-induced invaginations in the ER membrane, and impairs viral replication. Interestingly, this protein IFI6 had a faint effect on other mammalian RNA viruses, including HCV, which replicate in double membrane RCs that protrude outwards instead of inwards from the ER membrane [77].
5. Conclusions and Prospective
This review describes the main types, principles, and steps of pooled CRISPR KO screens. Since their application at the genome-scale, CRISPR tools have contributed tremendously to fundamental and translational studies. Various Cas proteins and sgRNAs have been used to develop gain-of-function CRISPRa and loss-of-function CRISPRi tools. Studying emerging, re-emerging, and persistent viruses such as those belonging to the Flaviviridae family and their crosstalk with host cells is crucial for discerning their biology and host response. In addition, this information can be exploited to formulate therapeutic molecules or improve existing strategies against flavivirus disease and transmission. It is noteworthy that there are subtle differences in data sets from independent CRISPR screens for the same viral infections. These can arise due to differences in experimental setups, virus strains, the target cell line used, or types of CRISPR screening. However, there are studies with typically overlapping gene hits for the same virus challenge. This reproducibility is remarkable and is the principal advantage of CRISPR-Cas9 technology. Moreover, with the advent of combinatorial CRISPR screens, one can easily define the most effective combinations of host factors or cellular signaling pathways that can be targeted to impede viral infections [78]. Although not discussed here, it is worth noting that additional virus-host interactions, such as RNA–RNA, protein-protein, and RNA–protein interactions, also contribute remarkably to our conception of flavivirus replication and pathogenesis [70,79,80]. In the recent past, there has been significant development in the CRISPR field, with the development of more precise Cas enzymes with single base editing abilities [81] or others with RNA targeting activities [82]. These tools are yet to be used extensively to examine flavivirus–host interactions.
Furthermore, since flaviviruses persist in arthropod vectors, it becomes vital to comprehend how host factor interactions overlap or differ between the insect vector and human hosts. This information could uncover fundamental interactions shared across diverse hosts and certain cellular factors that can be used as targets of drug/chemical or genetic manipulation in vector species to reduce virus dissemination. To date, there has been no genome-wide CRISPR screen to decode the mosquito-flavivirus–host factors. Some studies, however, have tried to systematically uncover insect host factors required for DENV-2 propagation by employing a genome-wide RNA interference screen in Drosophila melanogaster cells [64]. Such studies highlight the conservation of host dependency factors between dipteran and human hosts and reveal genetic factors that can be targeted to control infection in both the insect vector and the mammalian host [64]. As sequencing and gene annotations of Aedes mosquito and tick species improve, carrying out genome-wide screens to uncover flavivirus–host factors for these species becomes more feasible.
In conclusion, genome-wide CRISPR screens provide a powerful tool to decode the flavivirus–host interactions. We believe that future advances in applying this technology to other hosts and cell types will shed light on how flaviviruses evolve to exploit and subvert host functions. Such understanding will go a long way in uncovering the novel aspects of flavivirus biology and help develop better therapeutics and strategies for medical interventions and vector control.
Author Contributions
A.K.: original draft preparation, A.K., M.S. and S.T.: writing—review and editing, A.K., R.S.: visualization, S.T.: supervision, S.T.: funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the APC were funded by DBT-Wellcome Trust, grant number IA/I/18/1/503613 to ST.
Institutional Review Board Statement
The study did not require ethical approval.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
A.K. and M.S. are supported by the Graduate Fellowship from the Ministry of Education, Government of India. R.S. is supported by the SERB-FICCI PMRF fellowship. Illustrations were created using Biorender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fan, J.; Liu, Y.; Yuan, Z. Critical Role of Dengue Virus NS1 Protein in Viral Replication. Virol. Sin. 2014, 29, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.R.; de Sessions, P.F.; Leon, M.A.; Scholle, F. West Nile Virus Nonstructural Protein 1 Inhibits TLR3 Signal Transduction. J. Virol. 2008, 82, 8262–8271. [Google Scholar] [CrossRef] [PubMed]
- Falgout, B.; Pethel, M.; Zhang, Y.M.; Lai, C.J. Both Nonstructural Proteins NS2B and NS3 Are Required for the Proteolytic Processing of Dengue Virus Nonstructural Proteins. J. Virol. 1991, 65, 2467–2475. [Google Scholar] [CrossRef] [PubMed]
- Steinkühler, C.; Tomei, L.; De Francesco, R. In Vitro Activity of Hepatitis C Virus Protease NS3 Purified from Recombinant Baculovirus-Infected Sf9 Cells. J. Biol. Chem. 1996, 271, 6367–6373. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Jordan, J.L.; Sánchez-Burgos, G.G.; Laurent-Rolle, M.; García-Sastre, A. Inhibition of Interferon Signaling by Dengue Virus. Proc. Natl. Acad. Sci. USA 2003, 100, 14333–14338. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.H.; Fu, J.; Sugrue, R.J.; Yap, E.H.; Chan, Y.C.; Tan, Y.H. Recombinant Dengue Type 1 Virus NS5 Protein Expressed in Escherichia coli Exhibits RNA-Dependent RNA Polymerase Activity. Virology 1996, 216, 317–325. [Google Scholar] [CrossRef]
- Behrens, S.E.; Tomei, L.; De Francesco, R. Identification and Properties of the RNA-Dependent RNA Polymerase of Hepatitis C Virus. EMBO J. 1996, 15, 12–22. [Google Scholar] [CrossRef]
- Alvarez, D.E.; De Lella Ezcurra, A.L.; Fucito, S.; Gamarnik, A.V. Role of RNA Structures Present at the 3′UTR of Dengue Virus on Translation, RNA Synthesis, and Viral Replication. Virology 2005, 339, 200–212. [Google Scholar] [CrossRef]
- Gillespie, L.K.; Hoenen, A.; Morgan, G.; Mackenzie, J.M. The Endoplasmic Reticulum Provides the Membrane Platform for Biogenesis of the Flavivirus Replication Complex. J. Virol. 2010, 84, 10438–10447. [Google Scholar] [CrossRef]
- Klaitong, P.; Smith, D.R. Roles of Non-Structural Protein 4A in Flavivirus Infection. Viruses 2021, 13, 2077. [Google Scholar] [CrossRef]
- Welsch, S.; Miller, S.; Romero-Brey, I.; Merz, A.; Bleck, C.K.E.; Walther, P.; Fuller, S.D.; Antony, C.; Krijnse-Locker, J.; Bartenschlager, R. Composition and Three-Dimensional Architecture of the Dengue Virus Replication and Assembly Sites. Cell Host Microbe 2009, 5, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.E.; Gorbalenya, A.E.; Rice, C.M. The NS5A/NS5 Proteins of Viruses from Three Genera of the Family Flaviviridae Are Phosphorylated by Associated Serine/Threonine Kinases. J. Virol. 1998, 72, 6199–6206. [Google Scholar] [CrossRef] [PubMed]
- Dengue and Severe Dengue. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 22 August 2022).
- Fitzgerald, B.; Boyle, C.; Honein, M.A. Birth Defects Potentially Related to Zika Virus Infection During Pregnancy in the United States. JAMA 2018, 319, 1195–1196. [Google Scholar] [CrossRef] [PubMed]
- Factsheet about Japanese Encephalitis. Available online: https://www.ecdc.europa.eu/en/japanese-encephalitis/facts (accessed on 10 September 2022).
- Reed, K.D.; Meece, J.K.; Henkel, J.S.; Shukla, S.K. Birds, Migration and Emerging Zoonoses: West Nile Virus, Lyme Disease, Influenza A and Enteropathogens. Clin. Med. Res. 2003, 1, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Valentine, M.J.; Murdock, C.C.; Kelly, P.J. Sylvatic Cycles of Arboviruses in Non-Human Primates. Parasit. Vectors 2019, 12, 463. [Google Scholar] [CrossRef] [PubMed]
- Germain, M.; Francy, D.B.; Monath, T.P.; Ferrara, L.; Bryan, J.; Salaun, J.J.; Heme, G.; Renaudet, J.; Adam, C.; Digoutte, J.P. Yellow Fever in the Gambia, 1978–1979: Entomological Aspects and Epidemiological Correlations. Am. J. Trop. Med. Hyg. 1980, 29, 929–940. [Google Scholar] [CrossRef]
- Hanley, K.A.; Monath, T.P.; Weaver, S.C.; Rossi, S.L.; Richman, R.L.; Vasilakis, N. Fever versus Fever: The Role of Host and Vector Susceptibility and Interspecific Competition in Shaping the Current and Future Distributions of the Sylvatic Cycles of Dengue Virus and Yellow Fever Virus. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2013, 19, 292–311. [Google Scholar] [CrossRef]
- Puschnik, A.S.; Majzoub, K.; Ooi, Y.S.; Carette, J.E. A CRISPR Toolbox to Study Virus-Host Interactions. Nat. Rev. Microbiol. 2017, 15, 351–364. [Google Scholar] [CrossRef]
- Li, B.; Clohisey, S.M.; Chia, B.S.; Wang, B.; Cui, A.; Eisenhaure, T.; Schweitzer, L.D.; Hoover, P.; Parkinson, N.J.; Nachshon, A.; et al. Genome-Wide CRISPR Screen Identifies Host Dependency Factors for Influenza A Virus Infection. Nat. Commun. 2020, 11, 164. [Google Scholar] [CrossRef]
- Han, J.; Perez, J.T.; Chen, C.; Li, Y.; Benitez, A.; Kandasamy, M.; Lee, Y.; Andrade, J.; tenOever, B.; Manicassamy, B. Genome-Wide CRISPR/Cas9 Screen Identifies Host Factors Essential for Influenza Virus Replication. Cell Rep. 2018, 23, 596–607. [Google Scholar] [CrossRef]
- Grodzki, M.; Bluhm, A.P.; Schaefer, M.; Tagmount, A.; Russo, M.; Sobh, A.; Rafiee, R.; Vulpe, C.D.; Karst, S.M.; Norris, M.H. Genome-Scale CRISPR Screens Identify Host Factors That Promote Human Coronavirus Infection. Genome Med. 2022, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Baggen, J.; Persoons, L.; Vanstreels, E.; Jansen, S.; Van Looveren, D.; Boeckx, B.; Geudens, V.; De Man, J.; Jochmans, D.; Wauters, J.; et al. Genome-Wide CRISPR Screening Identifies TMEM106B as a Proviral Host Factor for SARS-CoV-2. Nat. Genet. 2021, 53, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, K.; Bae, S.; Park, J.; Lee, C.-K.; Kim, M.; Kim, E.; Kim, M.; Kim, S.; Kim, C.; et al. CRISPR/Cas9-Mediated Gene Knockout Screens and Target Identification via Whole-Genome Sequencing Uncover Host Genes Required for Picornavirus Infection. J. Biol. Chem. 2017, 292, 10664–10671. [Google Scholar] [CrossRef] [PubMed]
- Park, R.J.; Wang, T.; Koundakjian, D.; Hultquist, J.F.; Lamothe-Molina, P.; Monel, B.; Schumann, K.; Yu, H.; Krupzcak, K.M.; Garcia-Beltran, W.; et al. A Genome-Wide CRISPR Screen Identifies a Restricted Set of HIV Host Dependency Factors. Nat. Genet. 2017, 49, 193–203. [Google Scholar] [CrossRef]
- Yi, C.; Cai, C.; Cheng, Z.; Zhao, Y.; Yang, X.; Wu, Y.; Wang, X.; Jin, Z.; Xiang, Y.; Jin, M.; et al. Genome-Wide CRISPR-Cas9 Screening Identifies the CYTH2 Host Gene as a Potential Therapeutic Target of Influenza Viral Infection. Cell Rep. 2022, 38, 110559. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, S.; Li, W.; Yau, E.; Hui, H.; Singh, P.K.; Achuthan, V.; Young Karris, M.A.; Engelman, A.N.; Rana, T.M. Genome-Wide CRISPR/Cas9 Transcriptional Activation Screen Identifies a Histone Acetyltransferase Inhibitor Complex as a Regulator of HIV-1 Integration. Nucleic Acids Res. 2022, 50, 6687–6701. [Google Scholar] [CrossRef]
- Wiedenheft, B.; Sternberg, S.H.; Doudna, J.A. RNA-Guided Genetic Silencing Systems in Bacteria and Archaea. Nature 2012, 482, 331–338. [Google Scholar] [CrossRef]
- Terns, M.P.; Terns, R.M. CRISPR-Based Adaptive Immune Systems. Curr. Opin. Microbiol. 2011, 14, 321–327. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Soria, E. Intervening Sequences of Regularly Spaced Prokaryotic Repeats Derive from Foreign Genetic Elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Gasiunas, G.; Barrangou, R.; Horvath, P.; Siksnys, V. Cas9-CrRNA Ribonucleoprotein Complex Mediates Specific DNA Cleavage for Adaptive Immunity in Bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, E2579–E2586. [Google Scholar] [CrossRef] [PubMed]
- Shalem, O.; Sanjana, N.E.; Zhang, F. High-Throughput Functional Genomics Using CRISPR-Cas9. Nat. Rev. Genet. 2015, 16, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.A.; Horlbeck, M.A.; Adamson, B.; Villalta, J.E.; Chen, Y.; Whitehead, E.H.; Guimaraes, C.; Panning, B.; Ploegh, H.L.; Bassik, M.C.; et al. Genome-Scale CRISPR-Mediated Control of Gene Repression and Activation. Cell 2014, 159, 647–661. [Google Scholar] [CrossRef]
- Vigouroux, A.; Oldewurtel, E.; Cui, L.; Bikard, D.; van Teeffelen, S. Tuning DCas9’s Ability to Block Transcription Enables Robust, Noiseless Knockdown of Bacterial Genes. Mol. Syst. Biol. 2018, 14, e7899. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yin, C.; Zhang, T.; Li, F.; Yang, W.; Kaminski, R.; Fagan, P.R.; Putatunda, R.; Young, W.-B.; Khalili, K.; et al. CRISPR/GRNA-Directed Synergistic Activation Mediator (SAM) Induces Specific, Persistent and Robust Reactivation of the HIV-1 Latent Reservoirs. Sci. Rep. 2015, 5, 16277. [Google Scholar] [CrossRef] [PubMed]
- Bikard, D.; Jiang, W.; Samai, P.; Hochschild, A.; Zhang, F.; Marraffini, L.A. Programmable Repression and Activation of Bacterial Gene Expression Using an Engineered CRISPR-Cas System. Nucleic Acids Res. 2013, 41, 7429–7437. [Google Scholar] [CrossRef]
- Kampmann, M. CRISPRi and CRISPRa Screens in Mammalian Cells for Precision Biology and Medicine. ACS Chem. Biol. 2018, 13, 406–416. [Google Scholar] [CrossRef]
- Roesch, F.; OhAinle, M. HIV-CRISPR: A CRISPR/Cas9 Screening Method to Identify Genes Affecting HIV Replication. Bio Protoc. 2020, 10, e3614. [Google Scholar] [CrossRef]
- Heaton, B.E.; Kennedy, E.M.; Dumm, R.E.; Harding, A.T.; Sacco, M.T.; Sachs, D.; Heaton, N.S. A CRISPR Activation Screen Identifies a Pan-Avian Influenza Virus Inhibitory Host Factor. Cell Rep. 2017, 20, 1503–1512. [Google Scholar] [CrossRef]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef]
- Ikeda, A.; Fujii, W.; Sugiura, K.; Naito, K. High-Fidelity Endonuclease Variant HypaCas9 Facilitates Accurate Allele-Specific Gene Modification in Mouse Zygotes. Commun. Biol. 2019, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jung, M.-H.; Jeong, E.; Lee, J.K. Using Sniper-Cas9 to Minimize Off-Target Effects of CRISPR-Cas9 Without the Loss of On-Target Activity Via Directed Evolution. J. Vis. Exp. 2019, 144, e59202. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Jeong, E.; Lee, J.; Jung, M.; Shin, E.; Kim, Y.H.; Lee, K.; Jung, I.; Kim, D.; Kim, S.; et al. Directed Evolution of CRISPR-Cas9 to Increase Its Specificity. Nat. Commun. 2018, 9, 3048. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xu, J.; Ge, S.; Lai, L. CRISPR/Cas: Advances, Limitations, and Applications for Precision Cancer Research. Front. Med. 2021, 8, 649896. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.A.; Larson, M.H.; Morsut, L.; Liu, Z.; Brar, G.A.; Torres, S.E.; Stern-Ginossar, N.; Brandman, O.; Whitehead, E.H.; Doudna, J.A.; et al. CRISPR-Mediated Modular RNA-Guided Regulation of Transcription in Eukaryotes. Cell 2013, 154, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Aach, J.; Stranges, P.B.; Esvelt, K.M.; Moosburner, M.; Kosuri, S.; Yang, L.; Church, G.M. CAS9 Transcriptional Activators for Target Specificity Screening and Paired Nickases for Cooperative Genome Engineering. Nat. Biotechnol. 2013, 31, 833–838. [Google Scholar] [CrossRef]
- Burns, J.C.; Friedmann, T.; Driever, W.; Burrascano, M.; Yee, J.K. Vesicular Stomatitis Virus G Glycoprotein Pseudotyped Retroviral Vectors: Concentration to Very High Titer and Efficient Gene Transfer into Mammalian and Nonmammalian Cells. Proc. Natl. Acad. Sci. USA 1993, 90, 8033–8037. [Google Scholar] [CrossRef]
- Lohmann, V.; Körner, F.; Koch, J.; Herian, U.; Theilmann, L.; Bartenschlager, R. Replication of Subgenomic Hepatitis C Virus RNAs in a Hepatoma Cell Line. Science 1999, 285, 110–113. [Google Scholar] [CrossRef]
- Joung, J.; Konermann, S.; Gootenberg, J.S.; Abudayyeh, O.O.; Platt, R.J.; Brigham, M.D.; Sanjana, N.E.; Zhang, F. Genome-Scale CRISPR-Cas9 Knockout and Transcriptional Activation Screening. Nat. Protoc. 2017, 12, 828–863. [Google Scholar] [CrossRef]
- Hoenen, T.; Groseth, A. Virus–Host Cell Interactions. Cells 2022, 11, 804. [Google Scholar] [CrossRef]
- Marceau, C.D.; Puschnik, A.S.; Majzoub, K.; Ooi, Y.S.; Brewer, S.M.; Fuchs, G.; Swaminathan, K.; Mata, M.A.; Elias, J.E.; Sarnow, P.; et al. Genetic Dissection of Flaviviridae Host Factors through Genome-Scale CRISPR Screens. Nature 2016, 535, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.L.; Cherepanova, N.A.; Bozzacco, L.; MacDonald, M.R.; Gilmore, R.; Tai, A.W. Dengue Virus Hijacks a Noncanonical Oxidoreductase Function of a Cellular Oligosaccharyltransferase Complex. mBio 2017, 8, e00939-17. [Google Scholar] [CrossRef] [PubMed]
- Labeau, A.; Simon-Loriere, E.; Hafirassou, M.-L.; Bonnet-Madin, L.; Tessier, S.; Zamborlini, A.; Dupré, T.; Seta, N.; Schwartz, O.; Chaix, M.-L.; et al. A Genome-Wide CRISPR-Cas9 Screen Identifies the Dolichol-Phosphate Mannose Synthase Complex as a Host Dependency Factor for Dengue Virus Infection. J. Virol. 2020, 94, e01751-19. [Google Scholar] [CrossRef] [PubMed]
- Shue, B.; Chiramel, A.I.; Cerikan, B.; To, T.-H.; Frölich, S.; Pederson, S.M.; Kirby, E.N.; Eyre, N.S.; Bartenschlager, R.F.W.; Best, S.M.; et al. Genome-Wide CRISPR Screen Identifies RACK1 as a Critical Host Factor for Flavivirus Replication. J. Virol. 2021, 95, e00596-21. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Muffat, J.; Omer Javed, A.; Keys, H.R.; Lungjangwa, T.; Bosch, I.; Khan, M.; Virgilio, M.C.; Gehrke, L.; Sabatini, D.M.; et al. Genome-Wide CRISPR Screen for Zika Virus Resistance in Human Neural Cells. Proc. Natl. Acad. Sci. USA 2019, 116, 9527–9532. [Google Scholar] [CrossRef] [PubMed]
- Savidis, G.; McDougall, W.M.; Meraner, P.; Perreira, J.M.; Portmann, J.M.; Trincucci, G.; John, S.P.; Aker, A.M.; Renzette, N.; Robbins, D.R.; et al. Identification of Zika Virus and Dengue Virus Dependency Factors Using Functional Genomics. Cell Rep. 2016, 16, 232–246. [Google Scholar] [CrossRef]
- Ma, H.; Dang, Y.; Wu, Y.; Jia, G.; Anaya, E.; Zhang, J.; Abraham, S.; Choi, J.-G.; Shi, G.; Qi, L.; et al. A CRISPR-Based Screen Identifies Genes Essential for West-Nile-Virus-Induced Cell Death. Cell Rep. 2015, 12, 673–683. [Google Scholar] [CrossRef]
- Zhang, R.; Miner, J.J.; Gorman, M.J.; Rausch, K.; Ramage, H.; White, J.P.; Zuiani, A.; Zhang, P.; Fernandez, E.; Zhang, Q.; et al. A CRISPR Screen Defines a Signal Peptide Processing Pathway Required by Flaviviruses. Nature 2016, 535, 164–168. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, H.; Xiao, T.; Wang, Z.; Nie, X.; Li, X.; Qian, P.; Qin, L.; Han, X.; Zhang, J.; et al. CRISPR Screening of Porcine SgRNA Library Identifies Host Factors Associated with Japanese Encephalitis Virus Replication. Nat. Commun. 2020, 11, 5178. [Google Scholar] [CrossRef]
- Dukhovny, A.; Lamkiewicz, K.; Chen, Q.; Fricke, M.; Jabrane-Ferrat, N.; Marz, M.; Jung, J.U.; Sklan, E.H. A CRISPR Activation Screen Identifies Genes That Protect against Zika Virus Infection. J. Virol. 2019, 93, e00211-19. [Google Scholar] [CrossRef]
- Luu, A.P.; Yao, Z.; Ramachandran, S.; Azzopardi, S.A.; Miles, L.A.; Schneider, W.M.; Hoffmann, H.-H.; Bozzacco, L.; Garcia, G.; Gong, D.; et al. A CRISPR Activation Screen Identifies an Atypical Rho GTPase That Enhances Zika Viral Entry. Viruses 2021, 13, 2113. [Google Scholar] [CrossRef] [PubMed]
- Sessions, O.M.; Barrows, N.J.; Souza-Neto, J.A.; Robinson, T.J.; Hershey, C.L.; Rodgers, M.A.; Ramirez, J.L.; Dimopoulos, G.; Yang, P.L.; Pearson, J.L.; et al. Discovery of Insect and Human Dengue Virus Host Factors. Nature 2009, 458, 1047–1050. [Google Scholar] [CrossRef] [PubMed]
- Le Sommer, C.; Barrows, N.J.; Bradrick, S.S.; Pearson, J.L.; Garcia-Blanco, M.A. G Protein-Coupled Receptor Kinase 2 Promotes Flaviviridae Entry and Replication. PLoS Negl. Trop. Dis. 2012, 6, e1820. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, M.N.; Ng, A.; Sukumaran, B.; Gilfoy, F.D.; Uchil, P.D.; Sultana, H.; Brass, A.L.; Adametz, R.; Tsui, M.; Qian, F.; et al. RNA Interference Screen for Human Genes Associated with West Nile Virus Infection. Nature 2008, 455, 242–245. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Q.; Tiwari, S.K.; Lichinchi, G.; Yau, E.H.; Hui, H.; Li, W.; Furnari, F.; Rana, T.M. Integrin Avβ5 Internalizes Zika Virus during Neural Stem Cells Infection and Provides a Promising Target for Antiviral Therapy. Cell Rep. 2020, 30, 969–983.E4. [Google Scholar] [CrossRef]
- Hoffmann, H.-H.; Schneider, W.M.; Rozen-Gagnon, K.; Miles, L.A.; Schuster, F.; Razooky, B.; Jacobson, E.; Wu, X.; Yi, S.; Rudin, C.M.; et al. TMEM41B Is a Pan-Flavivirus Host Factor. Cell 2021, 184, 133–148.E20. [Google Scholar] [CrossRef]
- Scaturro, P.; Stukalov, A.; Haas, D.A.; Cortese, M.; Draganova, K.; Płaszczyca, A.; Bartenschlager, R.; Götz, M.; Pichlmair, A. An Orthogonal Proteomic Survey Uncovers Novel Zika Virus Host Factors. Nature 2018, 561, 253–257. [Google Scholar] [CrossRef]
- Shah, P.S.; Link, N.; Jang, G.M.; Sharp, P.P.; Zhu, T.; Swaney, D.L.; Johnson, J.R.; Von Dollen, J.; Ramage, H.R.; Satkamp, L.; et al. Comparative Flavivirus-Host Protein Interaction Mapping Reveals Mechanisms of Dengue and Zika Virus Pathogenesis. Cell 2018, 175, 1931–1945.E18. [Google Scholar] [CrossRef]
- Monel, B.; Compton, A.A.; Bruel, T.; Amraoui, S.; Burlaud-Gaillard, J.; Roy, N.; Guivel-Benhassine, F.; Porrot, F.; Génin, P.; Meertens, L.; et al. Zika Virus Induces Massive Cytoplasmic Vacuolization and Paraptosis-like Death in Infected Cells. EMBO J. 2017, 36, 1653–1668. [Google Scholar] [CrossRef]
- Lin, D.L.; Inoue, T.; Chen, Y.-J.; Chang, A.; Tsai, B.; Tai, A.W. The ER Membrane Protein Complex Promotes Biogenesis of Dengue and Zika Virus Non-Structural Multi-Pass Transmembrane Proteins to Support Infection. Cell Rep. 2019, 27, 1666–1674.E4. [Google Scholar] [CrossRef]
- Barrows, N.J.; Anglero-Rodriguez, Y.; Kim, B.; Jamison, S.F.; Le Sommer, C.; McGee, C.E.; Pearson, J.L.; Dimopoulos, G.; Ascano, M.; Bradrick, S.S.; et al. Dual Roles for the ER Membrane Protein Complex in Flavivirus Infection: Viral Entry and Protein Biogenesis. Sci. Rep. 2019, 9, 9711. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Canada, C.; Kelleher, D.J.; Gilmore, R. Cotranslational and Posttranslational N-Glycosylation of Polypeptides by Distinct Mammalian OST Isoforms. Cell 2009, 136, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.M.; Steitz, J.A. HuR and MRNA Stability. Cell. Mol. Life Sci. CMLS 2001, 58, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Gallay, P. Curing a Viral Infection by Targeting the Host: The Example of Cyclophilin Inhibitors. Antivir. Res. 2013, 99, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.B.; Ohlson, M.B.; Eitson, J.L.; Kumar, A.; McDougal, M.B.; Boys, I.N.; Mar, K.B.; De La Cruz-Rivera, P.C.; Douglas, C.; Konopka, G.; et al. A CRISPR Screen Identifies IFI6 as an ER-Resident Interferon Effector That Blocks Flavivirus Replication. Nat. Microbiol. 2018, 3, 1214–1223. [Google Scholar] [CrossRef]
- Zhou, P.; Chan, B.K.C.; Wan, Y.K.; Yuen, C.T.L.; Choi, G.C.G.; Li, X.; Tong, C.S.W.; Zhong, S.S.W.; Sun, J.; Bao, Y.; et al. A Three-Way Combinatorial CRISPR Screen for Analyzing Interactions among Druggable Targets. Cell Rep. 2020, 32, 108020. [Google Scholar] [CrossRef]
- Tham, H.-W.; Balasubramaniam, V.R.; Chew, M.-F.; Ahmad, H.; Hassan, S.S. Protein-Protein Interactions between A. Aegypti Midgut and Dengue Virus 2: Two-Hybrid Screens Using the Midgut CDNA Library. J. Infect. Dev. Ctries. 2015, 9, 1338–1349. [Google Scholar] [CrossRef]
- Mairiang, D.; Zhang, H.; Sodja, A.; Murali, T.; Suriyaphol, P.; Malasit, P.; Limjindaporn, T.; Finley, R.L. Identification of New Protein Interactions between Dengue Fever Virus and Its Hosts, Human and Mosquito. PLoS ONE 2013, 8, e53535. [Google Scholar] [CrossRef]
- Lee, H.K.; Oh, Y.; Hong, J.; Lee, S.H.; Hur, J.K. Development of CRISPR Technology for Precise Single-Base Genome Editing: A Brief Review. BMB Rep. 2021, 54, 98–105. [Google Scholar] [CrossRef]
- Burmistrz, M.; Krakowski, K.; Krawczyk-Balska, A. RNA-Targeting CRISPR–Cas Systems and Their Applications. Int. J. Mol. Sci. 2020, 21, 1122. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).