
Fighting Death for Living: Recent Advances in Molecular and Genetic Mechanisms Underlying Maize Lethal Necrosis Disease Resistance

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Occurrence and Threats of MLN Disease in SSA
3. MLN Causative Agents and Disease Management
3.1. Maize Chlorotic Mottle Virus (MCMV)
3.2. Potyviruses
3.3. Other MLN Associated Viruses
4. MLN Phenotyping and Management
4.1. Phenotyping and Diagnosis of MLN Disease
4.2. MLN Disease Management
5. Genetic Dissection of MLN Resistance
5.1. QTL Mapping of MLN Resistance
5.2. Genome Wide Association Studies (GWAS)
5.3. Cloning of Candidate Genesand Molecular Mechanisms Underlying MLN Resistance
6. Germplasm Identification and Molecular Breeding for MLN Resistance
6.1. Identification of Resistance Sources and Marker-Assisted Breeding
6.2. Application of Genomic Selection in MLN Resistance Breeding
7. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schaffnit-Chatterjee, C.; Lanzeni, M.L.; AG, D.B.; Hoffmann, R. Agricultural Value Chains in Sub-Saharan Africa. In From a Development Challenge to a Business Opportunity; Deutsche Bank Research: Frankfurt, Germany, 2014; pp. 1–28. [Google Scholar]
- Marenya, P.P.; Erenstein, O.; Prasanna, B.; Makumbi, D.; Jumbo, M.; Beyene, Y. Maize Lethal Necrosis Disease: Evaluating Agronomic and Genetic Control Strategies for Ethiopia and Kenya. Agric. Syst. 2018, 162, 220–228. [Google Scholar] [CrossRef]
- Mahuku, G.; Lockhart, B.E.; Wanjala, B.; Jones, M.W.; Kimunye, J.N.; Stewart, L.R.; Cassone, B.J.; Sevgan, S.; Nyasani, J.O.; Kusia, E.; et al. Maize Lethal Necrosis (MLN), an Emerging Threat to Maize-Based Food Security in Sub-Saharan Africa. Phytopathology 2015, 105, 956–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redinbaugh, M.G.; Stewart, L.R. Maize Lethal Necrosis: An Emerging, Synergistic Viral Disease. Annu. Rev. Virol. 2018, 5, 301–322. [Google Scholar] [CrossRef]
- E. Isabirye, B.; Rwomushana, I. Current and Future Potential Distribution of Maize Chlorotic Mottle Virus and Risk of Maize Lethal Necrosis Disease in Africa. J. Crop Prot. 2016, 5, 215–228. [Google Scholar] [CrossRef]
- Mahuku, G.; Wangai, A.; Sadessa, K.; Teklewold, A.; Wegary, D.; Ayalneh, D.; Adams, I.; Smith, J.; Bottomley, E.; Bryce, S. First Report of Maize Chlorotic Mottle Virus and Maize Lethal Necrosis on Maize in Ethiopia. Plant Dis. 2015, 99, 1870. [Google Scholar] [CrossRef]
- Boddupalli, P.; Suresh, L.M.; Mwatuni, F.; Beyene, Y.; Makumbi, D.; Gowda, M.; Olsen, M.; Hodson, D.; Worku, M.; Mezzalama, M.; et al. Maize Lethal Necrosis (MLN): Efforts toward Containing the Spread and Impact of a Devastating Transboundary Disease in Sub-Saharan Africa. Virus Res. 2020, 282, 197943. [Google Scholar] [CrossRef]
- Jiao, Z.; Tian, Y.; Wang, J.; Ismail, R.G.; Bondok, A.; Fan, Z. Advances in Research on Maize Lethal Necrosis, a Devastating Viral Disease. Phytopathol. Res. 2022, 4, 14. [Google Scholar] [CrossRef]
- Kwemoi, D.B. Genetic Analysis of Resistance to Maize Lethal Necrosis with Emphasis on Strategies for Improvement of Host Resistance. Ph.D. Thesis, University of the Free State, Bloemfontein, South Africa, 2021. [Google Scholar]
- Wangai, A.W.; Redinbaugh, M.G.; Kinyua, Z.M.; Miano, D.W.; Leley, P.K.; Kasina, M.; Mahuku, G.; Scheets, K.; Jeffers, D. First Report of Maize Chlorotic Mottle Virus and Maize Lethal Necrosis in Kenya. Plant Dis. 2012, 96, 1582. [Google Scholar] [CrossRef] [PubMed]
- Adams, I.; Harju, V.; Hodges, T.; Hany, U.; Skelton, A.; Rai, S.; Deka, M.; Smith, J.; Fox, A.; Uzayisenga, B.; et al. First Report of Maize Lethal Necrosis Disease in Rwanda. New Dis. Rep. 2014, 29, 22. [Google Scholar] [CrossRef] [Green Version]
- Asiimwe, T.; Stewart, L.R.; Willie, K.; Massawe, D.P.; Kamatenesi, J.; Redinbaugh, M.G. Maize Lethal Necrosis Viruses and Other Maize Viruses in Rwanda. Plant Pathol. 2020, 69, 585–597. [Google Scholar] [CrossRef]
- Lukanda, M.; Owati, A.; Ogunsanya, P.; Valimunzigha, K.; Katsongo, K.; Ndemere, H.; Kumar, P.L. First Report of Maize Chlorotic Mottle Virus Infecting Maize in the Democratic Republic of the Congo. Plant Dis. 2014, 98, 1448. [Google Scholar] [CrossRef] [PubMed]
- Frank, K.; Robert, G.; Brian, E.I. Status of Maize Lethal Necrosis in Eastern Uganda. Afr. J. Agric. Res. 2016, 11, 652–660. [Google Scholar] [CrossRef] [Green Version]
- Eunice, J.; Miano, D.; W.M, M.; Mutitu, E.; Macharia, I. Status of Maize Lethal Necrosis Disease in Seed Production System in Kenya. Cogent Food Agric. 2021, 7, 1918406. [Google Scholar] [CrossRef]
- De Groote, H.; Oloo, F.; Tongruksawattana, S.; Das, B. Community-Survey Based Assessment of the Geographic Distribution and Impact of Maize Lethal Necrosis (MLN) Disease in Kenya. Crop Prot. 2016, 82, 30–35. [Google Scholar] [CrossRef]
- Bockelman, D.L.; Claflin, L.E.; Uyemoto, J.K. Host Range and Seed-Transmission Studies of Maize Chlorotic Mottle Virus in Grasses and Corn. Plant Dis. 1982, 66, 216–218. [Google Scholar] [CrossRef]
- Niblett, C.L.; Claflin, L.E. Corn Lethal Necrosis-a New Virus Disease of Corn in Kansas. Plant Dis. Report. 1978, 62, 15–19. [Google Scholar]
- Goldberg, K.-B.; Brakke, M.K. Concentration of Maize Chlorotic Mottle Virus Increased in Mixed Infections with Maize Dwarf Mosaic Virus, Strain, B. Pap. Plant Pathol. 1987, 177, 162–167. [Google Scholar] [CrossRef]
- Leitich, R.K.; Korir, J.C.; Muoma, J.O.; Omayio, D.O. Viral Synergism and Its Role in Management of Maize Lethal Necrosis Disease. Afr. J. Plant Sci. 2021, 15, 151–157. [Google Scholar]
- Scheets, K. Maize Chlorotic Mottle Machlomovirus and Wheat Streak Mosaic Rymovirus Concentrations Increase in the Synergistic Disease Corn Lethal Necrosis. Virology 1998, 242, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Stenger, D.C.; French, R. Complete Nucleotide Sequence of a Maize Chlorotic Mottle Virus Isolate from Nebraska. Arch. Virol. 2008, 153, 995–997. [Google Scholar] [CrossRef]
- Castillo, J.; Hebert, T. A New Virus Disease of Maize in Peru. Fitopatologia 1974, 9, 79–84. [Google Scholar]
- Xie, L.; Zhang, J.; Wang, Q.; Meng, C.; Hong, J.; Zhou, X. Characterization of Maize Chlorotic Mottle Virus Associated with Maize Lethal Necrosis Disease in China. J. Phytopathol. 2011, 159, 191–193. [Google Scholar] [CrossRef]
- Uyemoto, J.; Claflin, L.; Wilson, D.; Raney, R. Maize Chlorotic Mottle and Maize Dwarf Mosaic Viruses; Effect of Single and Double Inoculations on Symptomatology and Yield. Plant Dis. 1981, 65, 39–41. [Google Scholar] [CrossRef]
- Scheets, K. Maize Chlorotic Mottle. In Viruses and Virus Diseases of Poaceae (Gramineae); Institut National de la Recherche Agronomique: Paris, France, 2004. [Google Scholar]
- Nutter, R.; Scheets, K.; Panganiban, L.; Lommel, S. The Complete Nucleotide Sequence of the Maize Chlorotic Mottle Virus Genome. Nucleic Acids Res. 1989, 17, 3163–3177. [Google Scholar] [CrossRef] [Green Version]
- Scheets, K. Analysis of Gene Functions in Maize Chlorotic Mottle Virus. Virus Res. 2016, 222, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carino, E.J.; Scheets, K.; Miller, W.A. The RNA of Maize Chlorotic Mottle Virus, an Obligatory Component of Maize Lethal Necrosis Disease, Is Translated via a Variant Panicum Mosaic Virus-like Cap-Independent Translation Element. J. Virol. 2020, 94, e01005–e01020. [Google Scholar] [CrossRef]
- Jiao, Z.; Tian, Y.; Cao, Y.; Wang, J.; Zhan, B.; Zhao, Z.; Sun, B.; Guo, C.; Ma, W.; Liao, Z.; et al. A Novel Pathogenicity Determinant Hijacks Maize Catalase 1 to Enhance Viral Multiplication and Infection. New Phytol. 2021, 230, 1126–1141. [Google Scholar] [CrossRef]
- Scheets, K. Maize Chlorotic Mottle Machlomovirus Expresses Its Coat Protein from a 1.47-Kb Subgenomic RNA and Makes a 0.34-Kb Subgenomic RNA. Virology 2000, 267, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Nault, L.R.; Styer, W.E.; Coffey, M.E.; Gordon, D.T.; Negi, L.S.; Niblett, C.L. Transmission of Maize Chlorotic Mottle Virus by Chrysomelid Beetles. Phytopathology 1978, 68, 1071–1074. [Google Scholar] [CrossRef] [Green Version]
- Cabanas, D.; Watanabe, S.; Higashi, C.; Bressan, A. Dissecting the Mode of Maize Chlorotic Mottle Virus Transmission (Tombusviridae: Machlomovirus) by Frankliniella Williamsi (Thysanoptera: Thripidae). J. Econ. Entomol. 2013, 106, 16–24. [Google Scholar] [CrossRef]
- Nyasani, J.O.; Meyhöfer, R.; Subramanian, S.; Poehling, H.-M. Effect of Intercrops on Thrips Species Composition and Population Abundance on French Beans in Kenya. Entomol. Exp. Appl. 2012, 142, 236–246. [Google Scholar] [CrossRef]
- Jensen, S.G.; Wysong, D.S.; Ball, E.M.; Higley, P.M. Seed Transmission of Maize Chlorotic Mottle Virus. Plant Dis. 1991, 75, 497–498. [Google Scholar] [CrossRef]
- Regassa, B.; Wegary, D.; Fininsa, C.; Abraham, A. Screening Maize Genotypes for Resistance to Maize Lethal Necrosis Disease in Ethiopia. Trop. Plant Pathol. 2021, 46, 583–595. [Google Scholar] [CrossRef]
- Mwando, N.L.; Tamiru, A.; Nyasani, J.O.; Obonyo, M.A.O.; Caulfield, J.C.; Bruce, T.J.A.; Subramanian, S. Maize Chlorotic Mottle Virus Induces Changes in Host Plant Volatiles That Attract Vector Thrips Species. J. Chem. Ecol. 2018, 44, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Lain, S.; García, J.A. Highlights and Prospects of Potyvirus Molecular Biology. J. Gen. Virol. 1992, 73, 1–16. [Google Scholar] [CrossRef]
- Keizerweerd, A.T.; Chandra, A.; Grisham, M.P. Development of a Reverse Transcription Loop-Mediated Isothermal Amplification (RT-LAMP) Assay for the Detection of Sugarcane Mosaic Virus and Sorghum Mosaic Virus in Sugarcane. J. Virol. Methods 2015, 212, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Louie, R.; Knoke, J. Strains of Maize Dwarf Mosaic Virus. Plant Dis. Report. 1975, 59, 518–522. [Google Scholar]
- Williams, L.E.; Alexander, L. Maize Dwarf Mosaic, a New Corn Disease. Phytopathology 1965, 55, 802–804. [Google Scholar]
- Stewart, L.R.; Willie, K.; Wijeratne, S.; Redinbaugh, M.G.; Massawe, D.; Niblett, C.L.; Kiggundu, A.; Asiimwe, T. Johnsongrass Mosaic Virus Contributes to Maize Lethal Necrosis in East Africa. Plant Dis. 2017, 101, 1455–1462. [Google Scholar] [CrossRef] [Green Version]
- Shukla, D.T.; Ward, C. Structure of Potyvirus Coat Proteins and Its Application in the Taxonomy of the Potyvirus Group. Adv. Virus Res. 1989, 36, 273–314. [Google Scholar]
- Louie, R. Sugarcane Mosaic Virus in Kenya. Plant Dis. 1980, 64, 944–947. [Google Scholar] [CrossRef] [Green Version]
- Urcuqui-Inchima, S.; Haenni, A.-L.; Bernardi, F. Potyvirus Proteins: A Wealth of Functions. Virus Res. 2001, 74, 157–175. [Google Scholar] [CrossRef] [PubMed]
- Revers, F.; García, J.A. Molecular Biology of Potyviruses. Adv. Virus Res. 2015, 92, 101–199. [Google Scholar] [PubMed]
- Vijayapalani, P.; Maeshima, M.; Nagasaki-Takekuchi, N.; Miller, W.A. Interaction of the Trans-Frame Potyvirus Protein P3N-PIPO with Host Protein PCaP1 Facilitates Potyvirus Movement. PLoS Pathog. 2012, 8, e1002639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valli, A.; Gallo, A.; Calvo, M.; Pérez, J.d.J.; García, J.A. A Novel Role of the Potyviral Helper Component Proteinase Contributes to Enhance the Yield of Viral Particles. J. Virol. 2014, 88, 9808–9818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valli, A.A.; Gallo, A.; Rodamilans, B.; López-Moya, J.J.; García, J.A. The HCPro from the Potyviridae Family: An Enviable Multitasking Helper Component That Every Virus Would like to Have. Mol. Plant Pathol. 2018, 19, 744–763. [Google Scholar] [CrossRef]
- Govier, D.; Kassanis, B. A Virus-Induced Component of Plant Sap Needed When Aphids Acquire Potato Virus Y from Purified Preparations. Virology 1974, 61, 420–426. [Google Scholar] [CrossRef]
- Maia, I.G.; Haenni, A.-L.; Bernardi, F. Potyviral HC-Pro: A Multifunctional Protein. J. Gen. Virol. 1996, 77, 1335–1341. [Google Scholar] [CrossRef]
- Syller, J. The Roles and Mechanisms of Helper Component Proteins Encoded by Potyviruses and Caulimoviruses. Physiol. Mol. Plant Pathol. 2005, 67, 119–130. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Gautam, S.; Rasmussen, D.A.; Srinivasan, R. Aphid transmission of Potyvirus: The largest plant-infecting RNA virus genus. Viruses 2020, 12, 773. [Google Scholar] [CrossRef]
- Wamaitha, M.J.; Nigam, D.; Maina, S.; Stomeo, F.; Wangai, A.; Njuguna, J.N.; Holton, T.A.; Wanjala, B.W.; Wamalwa, M.; Lucas, T.; et al. Metagenomic analysis of viruses associated with maize lethal necrosis in Kenya. Virol. J. 2018, 15, 90. [Google Scholar] [CrossRef] [PubMed]
- Yahaya, A.; Al Rwahnih, M.; Dangora, D.; Gregg, L.; Alegbejo, M.; Lava Kumar, P.; Alabi, O. First Report of Maize Yellow Mosaic Virus Infecting Sugarcane (Saccharum Spp.) and Itch Grass (Rottboelliacochinchinensis) in Nigeria. Plant Dis. 2017, 101, 1335. [Google Scholar] [CrossRef]
- Chepkorir, T.; Sylvester, A.; Jane, W.; Bramwel, W.W. Occurrence of Maize Yellow Mosaic Virus and Evidence of Co-Infection with Maize Lethal Necrosis Viruses in Bomet County, Kenya. Afr. J. Plant Sci. 2021, 15, 299–308. [Google Scholar]
- Maize. Org. Leading the Fight against Maize Lethal Necrosis. CRP Annu. Rep. 2013. Available online: https://annualreport2013.maize.org/leading-the-fight-against-maize-lethal-necrosis/ (accessed on 12 September 2022).
- Fatma, H.K.; Tileye, F.; Patrick, A.N. Insights of Maize Lethal Necrotic Disease: A Major Constraint to Maize Production in East Africa. Afr. J. Microbiol. Res. 2016, 10, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Boonham, N.; Kreuze, J.; Winter, S.; van der Vlugt, R.; Bergervoet, J.; Tomlinson, J.; Mumford, R. Methods in Virus Diagnostics: From ELISA to next Generation Sequencing. Virus Res. 2014, 186, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Mezzalama, M.; Das, B.; Prasanna, B. MLN Pathogen Diagnosis, MLN-Free Seed Production and Safe Exchange to Non-Endemic Countries; CIMMYT: Texcoco, Mexico, 2015. [Google Scholar]
- Zhan, B.-H.; Yang, X.-L.; Lommel, S.A.; Zhou, X.-P. Recent Progress in Maize Lethal Necrosis Disease: From Pathogens to Integrated Pest Management. J. Integr. Agric. 2022, 21, 3445–3455. [Google Scholar] [CrossRef]
- Phillips, N.; Uyemoto, J.; Wilson, D. Maize Chlorotic Mottle Virus and Crop Rotation: Effect of Sorghum on Virus Incidence. Plant Dis. 1982, 66, 376–379. [Google Scholar] [CrossRef]
- Jones, M.W.; Redinbaugh, M.G.; Louie, R. The Mdm1 Locus and Maize Resistance to Maize Dwarf Mosaic Virus. Plant Dis. 2007, 91, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Zambrano, J.L.; Jones, M.W.; Brenner, E.; Francis, D.M.; Tomas, A.; Redinbaugh, M.G. Genetic Analysis of Resistance to Six Virus Diseases in a Multiple Virus-Resistant Maize Inbred Line. Theor. Appl. Genet. 2014, 127, 867–880. [Google Scholar] [CrossRef]
- Murithi, A.; Olsen, M.S.; Kwemoi, D.B.; Veronica, O.; Ertiro, B.T.; M., S.L.; Beyene, Y.; Das, B.; Prasanna, B.M.; Gowda, M. Discovery and Validation of a Recessively Inherited Major-Effect QTL Conferring Resistance to Maize Lethal Necrosis (MLN) Disease. Front. Genet. 2021, 12, 767883. [Google Scholar] [CrossRef]
- Jones, M.W.; Penning, B.W.; Jamann, T.M.; Glaubitz, J.C.; Romay, C.; Buckler, E.S.; Redinbaugh, M.G. Diverse Chromosomal Locations of Quantitative Trait Loci for Tolerance to Maize Chlorotic Mottle Virus in Five Maize Populations. Phytopathology 2018, 108, 748–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitonik, C.; Suresh, L.; Beyene, Y.; Olsen, M.S.; Makumbi, D.; Oliver, K.; Das, B.; Bright, J.M.; Mugo, S.; Crossa, J. Genetic Architecture of Maize Chlorotic Mottle Virus and Maize Lethal Necrosis through GWAS, Linkage Analysis and Genomic Prediction in Tropical Maize Germplasm. Theor. Appl. Genet. 2019, 132, 2381–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlson, E.W.; Redinbaugh, M.G.; Jones, M.W. Mapping Maize Chlorotic Mottle Virus Tolerance Loci in the Maize 282 Association Panel. Crop Sci. 2022, 62, 1497–1510. [Google Scholar] [CrossRef]
- Beyene, Y.; Gowda, M.; Suresh, L.M.; Mugo, S.; Olsen, M.; Oikeh, S.O.; Juma, C.; Tarekegne, A.; Prasanna, B.M. Genetic Analysis of Tropical Maize Inbred Lines for Resistance to Maize Lethal Necrosis Disease. Euphytica 2017, 213, 224. [Google Scholar] [CrossRef] [Green Version]
- Nyaga, C.; Gowda, M.; Beyene, Y.; Murithi, W.T.; Burgueno, J.; Toledo, F.; Makumbi, D.; Olsen, M.S.; Das, B.; M., S.L.; et al. Hybrid Breeding for MLN Resistance: Heterosis, Combining Ability, and Hybrid Prediction. Plants 2020, 9, 468. [Google Scholar] [CrossRef] [Green Version]
- Redinbaugh, M.G.; Zambrano, J.L. Control of Virus Diseases in Maize. Adv. Virus Res. 2014, 90, 391–429. [Google Scholar]
- Awata, L.A.O.; Beyene, Y.; Gowda, M.; M., S.L.; Jumbo, M.B.; Tongoona, P.; Danquah, E.; Ifie, B.E.; Marchelo-Dragga, P.W.; Olsen, M.; et al. Genetic Analysis of QTL for Resistance to Maize Lethal Necrosis in Multiple Mapping Populations. Genes 2020, 11, 32. [Google Scholar] [CrossRef] [Green Version]
- Awata, L.A.O.; Ifie, B.E.; Danquah, E.; Jumbo, M.B.; Suresh, L.M.; Gowda, M.; Marchelo-Dragga, P.W.; Olsen, M.S.; Shorinola, O.; Yao, N.K.; et al. Introgression of Maize Lethal Necrosis Resistance Quantitative Trait Loci into Susceptible Maize Populations and Validation of the Resistance under Field Conditions in Naivasha, Kenya. Front. Plant Sci. 2021, 12, 649308. [Google Scholar] [CrossRef]
- Jones, M.W.; Redinbaugh, M.G.; Anderson, R.J.; Louie, R. Identification of Quantitative Trait Loci Controlling Resistance to Maize Chlorotic Dwarf Virus. Theor. Appl. Genet. 2004, 110, 48–57. [Google Scholar] [CrossRef]
- Ming, R.; Brewbaker, J.L.; Pratt, R.C.; Musket, T.A.; McMullen, M.D. Molecular Mapping of a Major Gene Conferring Resistance to Maize Mosaic Virus. Theor. Appl. Genet. 1997, 95, 271–275. [Google Scholar] [CrossRef]
- Redinbaugh, M.G.; Pratt, R.C. Virus Resistance. In Handbook of Maize: Its Biology; Springer: Berlin/Heidelberg, Germany, 2009; pp. 251–270. [Google Scholar]
- Zhang, Z.-Y.; Wang, Y.-G.; Shen, X.-J.; Li, L.; Zhou, S.-F.; Li, W.-C.; Fu, F.-L. RNA Interference-Mediated Resistance to Maize Dwarf Mosaic Virus. Plant Cell Tissue Organ Cult. 2013, 113, 571–578. [Google Scholar] [CrossRef]
- Gowda, M.; Beyene, Y.; Makumbi, D.; Semagn, K.; Olsen, M.S.; Bright, J.M.; Das, B.; Mugo, S.; Suresh, L.M.; Prasanna, B.M. Discovery and Validation of Genomic Regions Associated with Resistance to Maize Lethal Necrosis in Four Biparental Populations. Mol. Breed. 2018, 38, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza, I.R.P.; Schuelter, A.R.; Guimarães, C.T.; Schuster, I.; De Oliveira, E.; Redinbaugh, M. Mapping QTL Contributing to SCMV Resistance in Tropical Maize. Hereditas 2008, 145, 167–173. [Google Scholar] [CrossRef]
- Gowda, M.; Das, B.; Makumbi, D.; Babu, R.; Semagn, K.; Mahuku, G.; Olsen, M.S.; Bright, J.M.; Beyene, Y.; Prasanna, B.M. Genome-Wide Association and Genomic Prediction of Resistance to Maize Lethal Necrosis Disease in Tropical Maize Germplasm. Theor. Appl. Genet. 2015, 128, 1957–1968. [Google Scholar] [CrossRef] [Green Version]
- Sadessa, K.; Beyene, Y.; Ifie, B.E.; Suresh, L.; Olsen, M.S.; Ogugo, V.; Wegary, D.; Tongoona, P.; Danquah, E.; Offei, S.K. Identification of Genomic Regions Associated with Agronomic and Disease Resistance Traits in a Large Set of Multiple DH Populations. Genes 2022, 13, 351. [Google Scholar] [CrossRef]
- Tao, Y.; Jiang, L.; Liu, Q.; Zhang, Y.; Zhang, R.; Ingvardsen, C.R.; Frei, U.K.; Wang, B.; Lai, J.; Lübberstedt, T.; et al. Combined Linkage and Association Mapping Reveals Candidates for Scmv1, a Major Locus Involved in Resistance to Sugarcane Mosaic Virus (SCMV) in Maize. BMC Plant Biol. 2013, 13, 162. [Google Scholar] [CrossRef] [Green Version]
- Ingvardsen, C.R.; Xing, Y.; Frei, U.K.; Lübberstedt, T. Genetic and Physical Fine Mapping of Scmv2, a Potyvirus Resistance Gene in Maize. Theor. Appl. Genet. 2010, 120, 1621–1634. [Google Scholar] [CrossRef]
- Leng, P.; Ji, Q.; Asp, T.; Frei, U.K.; Ingvardsen, C.R.; Xing, Y.; Studer, B.; Redinbaugh, M.; Jones, M.; Gajjar, P.; et al. Auxin Binding Protein 1 Reinforces Resistance to Sugarcane Mosaic Virus in Maize. Mol. Plant 2017, 10, 1357–1360. [Google Scholar] [CrossRef]
- Leng, P.; Ji, Q.; Tao, Y.; Ibrahim, R.; Pan, G.; Xu, M.; Lübberstedt, T. Characterization of Sugarcane Mosaic Virus Scmv1 and Scmv2 Resistance Regions by Regional Association Analysis in Maize. PLoS ONE 2015, 10, e0140617. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, H.; Gong, Y.; Tao, Y.; Jiang, L.; Zuo, W.; Yang, Q.; Ye, J.; Lai, J.; Wu, J.; et al. An Atypical Thioredoxin Imparts Early Resistance to Sugarcane Mosaic Virus in Maize. Mol. Plant 2017, 10, 483–497. [Google Scholar] [CrossRef] [Green Version]
- Semagn, K.; Beyene, Y.; Babu, R.; Nair, S.; Gowda, M.; Das, B.; Tarekegne, A.; Mugo, S.; Mahuku, G.; Worku, M.; et al. Quantitative Trait Loci Mapping and Molecular Breeding for Developing Stress Resilient Maize for Sub-Saharan Africa. Crop Sci. 2015, 55, 1449–1459. [Google Scholar] [CrossRef]
- Wen, W.; Araus, J.L.; Shah, T.; Cairns, J.; Mahuku, G.; Bänziger, M.; Torres, J.L.; Sánchez, C.; Yan, J. Molecular Characterization of a Diverse Maize Inbred Line Collection and Its Potential Utilization for Stress Tolerance Improvement. Crop Sci. 2011, 51, 2569–2581. [Google Scholar] [CrossRef]
- Yu, J.; Buckler, E.S. Genetic Association Mapping and Genome Organization of Maize. Curr. Opin. Biotechnol. 2006, 17, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Riedelsheimer, C.; Technow, F.; Melchinger, A.E. Comparison of Whole-Genome Prediction Models for Traits with Contrasting Genetic Architecture in a Diversity Panel of Maize Inbred Lines. BMC Genom. 2012, 13, 452. [Google Scholar] [CrossRef] [Green Version]
- Nyaga, C.; Gowda, M.; Beyene, Y.; Muriithi, W.T.; Makumbi, D.; Olsen, M.S.; Suresh, L.M.; Bright, J.M.; Das, B.; Prasanna, B.M. Genome-Wide Analyses and Prediction of Resistance to MLN in Large Tropical Maize Germplasm. Genes 2019, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, T.J.; de Leon, N.; Kaeppler, S.M.; Tracy, W.F. Genetic Analysis of Sugarcane Mosaic Virus Resistance in the Wisconsin Diversity Panel of Maize. Crop Sci. 2018, 58, 1853–1865. [Google Scholar] [CrossRef]
- Jones, M.W.; Boyd, E.C.; Redinbaugh, M.G. Responses of Maize (Zea Mays L.) near Isogenic Lines Carrying Wsm1, Wsm2, and Wsm3 to Three Viruses in the Potyviridae. Theor. Appl. Genet. 2011, 123, 729–740. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, Y.; Ding, X.S.; Webb, S.L.; Zhou, T.; Nelson, R.S.; Fan, Z. Maize Elongin C Interacts with the Viral Genome-linked Protein, VP g, of Sugarcane Mosaic Virus and Facilitates Virus Infection. New Phytol. 2014, 203, 1291–1304. [Google Scholar] [CrossRef]
- Yang, Z.; Dong, M.; Cheng, G.; Liu, S.; Zhang, H.; Shang, H.; Zhou, Y.; Huang, G.; Zhang, M.; Wang, F.; et al. Selective Interaction of Sugarcane EIF4E with VPgs from Sugarcane Mosaic Pathogens. Viruses 2021, 13, 518. [Google Scholar] [CrossRef]
- Chen, H.; Cao, Y.; Li, Y.; Xia, Z.; Xie, J.; Carr, J.P.; Wu, B.; Fan, Z.; Zhou, T. Identification of Differentially Regulated Maize Proteins Conditioning Sugarcane Mosaic Virus Systemic Infection. New Phytol. 2017, 215, 1156–1172. [Google Scholar] [CrossRef] [Green Version]
- Dang, M.; Cheng, Q.; Hu, Y.; Wu, J.; Zhou, X.; Qian, Y. Proteomic Changes during MCMV Infection Revealed by ITRAQ Quantitative Proteomic Analysis in Maize. Int. J. Mol. Sci. 2019, 21, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Liu, S.; Chen, X.; Prasanna, B.M.; Ni, Z.; Li, X.; He, Y.; Fan, Z.; Zhou, T. Maize MiR167-ARF3/30-Polyamine Oxidase 1 Module-Regulated H2O2 Production Confers Resistance to Maize Chlorotic Mottle Virus. Plant Physiol. 2022, 189, 1065–1082. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Ingvardsen, C.; Salomon, R.; Lübberstedt, T. Analysis of Sugarcane Mosaic Virus Resistance in Maize in an Isogenic Dihybrid Crossing Scheme and Implications for Breeding Potyvirus-Resistant Maize Hybrids. Genome 2006, 49, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.R.; Haque, M.A.; Jones, M.W.; Redinbaugh, M.G. Response of Maize (Zea Mays L.) Lines Carrying Wsm1, Wsm2, and Wsm3 to the Potyviruses Johnsongrass Mosaic Virus and Sorghum Mosaic Virus. Mol. Breed. 2013, 31, 289–297. [Google Scholar] [CrossRef]
- Użarowska, A.; Dionisio, G.; Sarholz, B.; Piepho, H.-P.; Xu, M.; Ingvardsen, C.R.; Wenzel, G.; Lübberstedt, T. Validation of Candidate Genes Putatively Associated with Resistance to SCMV and MDMV in Maize (Zea Mays L.) by Expression Profiling. BMC Plant Biol. 2009, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Han, Z.; Wang, S.; Wang, X.; Sun, A.; Zu, X.; Chen, Y. Comparative Proteomic Analysis of the Plant–Virus Interaction in Resistant and Susceptible Ecotypes of Maize Infected with Sugarcane Mosaic Virus. J. Proteom. 2013, 89, 124–140. [Google Scholar] [CrossRef]
- Xu, X.-J.; Geng, C.; Jiang, S.-Y.; Zhu, Q.; Yan, Z.-Y.; Tian, Y.-P.; Li, X.-D. A Maize Triacylglycerol Lipase Inhibits Sugarcane Mosaic Virus Infection. Plant Physiol. 2022, 189, 754–771. [Google Scholar] [CrossRef]
- Yuan, W.; Chen, H.; Xie, J.; Li, M.; Fan, Z.; Zhou, T. Maize Phenylalanine Ammonia Lyases Contribute to Resistance to Sugarcane Mosaic Virus Infection; APSNET: Saint Paul, MN, USA, 2018. [Google Scholar]
- Cao, N.; Zhan, B.; Zhou, X. Nitric Oxide as a Downstream Signaling Molecule in Brassinosteroid-Mediated Virus Susceptibility to Maize Chlorotic Mottle Virus in Maize. Viruses 2019, 11, 368. [Google Scholar] [CrossRef]
- Iqbal, M.S.; Jabbar, B.; Sharif, M.N.; Ali, Q.; Husnain, T.; Nasir, I.A. In Silico MCMV Silencing Concludes Potential Host-Derived MiRNAs in Maize. Front. Plant Sci. 2017, 8, 372. [Google Scholar] [CrossRef] [Green Version]
- Uyemoto, J.; Bockelman, D.; Claflin, L. Severe Outbreak of Corn Lethal Necrosis Disease in Kansas. Plant Dis. 1980, 64, 99–100. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Kuntze, L.; Gumber, R.K.; Lübberstedt, T.; Fuchs, E. Genetic Basis of Resistance to Sugarcane Mosaic Virus in European Maize Germplasm. Theor. Appl. Genet. 1998, 96, 1151–1161. [Google Scholar] [CrossRef]
- Prasanna, B. Maize Lethal Necrosis (MLN) in Eastern Africa: Tackling a Major Challenge. Afr. Seed 2015, 2015, 18–21. [Google Scholar]
- Romay, M.C.; Millard, M.J.; Glaubitz, J.C.; Peiffer, J.A.; Swarts, K.L.; Casstevens, T.M.; Elshire, R.J.; Acharya, C.B.; Mitchell, S.E.; Flint-Garcia, S.A. Comprehensive Genotyping of the USA National Maize Inbred Seed Bank. Genome Biol. 2013, 14, R55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathias, T.; Kabamba, M.; Kayombo, K.; Victoria, N.; Dickson, N.; Lubasi, C.; Cosmos, M.; Lingadahali, M.S. Screening of Maize Germplasm for Resistance to Maize Lethal Necrosis Disease in Zambia. Afr. J. Biotechnol. 2021, 20, 25–32. [Google Scholar] [CrossRef]
- Xia, X.; Melchinger, A.E.; Kuntze, L.; Lübberstedt, T. Quantitative Trait Loci Mapping of Resistance to Sugarcane Mosaic Virus in Maize. Phytopatholog 1999, 89, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Lübberstedt, T.; Xia, X.; Xu, M.; Kuntze, L.; Melchinger, A. Inheritance of Resistance to SCMV and MDMV in European Maize. In Genetics and Breeding for Crop Quality and Resistance; Springer: Berlin/Heidelberg, Germany, 1999; pp. 241–250. [Google Scholar]
- Das, B.; Atlin, G.N.; Olsen, M.; Burgueño, J.; Tarekegne, A.; Babu, R.; Ndou, E.N.; Mashingaidze, K.; Moremoholo, L.; Ligeyo, D.; et al. Identification of Donors for Low-Nitrogen Stress with Maize Lethal Necrosis (MLN) Tolerance for Maize Breeding in Sub-Saharan Africa. Euphytica 2019, 215, 80. [Google Scholar] [CrossRef] [Green Version]
- CIMMYT. Maize Lethal Necrosis (MLN): A Technical Manual for Disease Management; Prasanna, B.M., Ed.; CIMMYT: Texcoco, Mexico, 2021; Available online: https://mln.cimmyt.org/wp-content/uploads/sites/39/2021/10/MLN-Disease-Management_Technical-Manual-CIMMYT.pdf (accessed on 12 September 2022).
- Murithi, A.K. Mapping and Validation of Major Quantitative Trait Loci (Qtl) for Maize Lethal Necrosis (Mln) Resistance in Maize. Master Degree Thesis, University of Nairobi, Nairobi, Kenya, 2020. [Google Scholar]
- Karanja, J.; Derera, J.; Gubba, A.; Mugo, S.; Wangai, A. Response of Selected Maize Inbred Germplasm to Maize Lethal Necrosis Disease and Its Causative Viruses (Sugarcane Mosaic Virus and Maize Chlorotic Mottle Virus) in Kenya. Open Agric. J. 2018, 12, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Jannink, J.-L.; Lorenz, A.J.; Iwata, H. Genomic Selection in Plant Breeding: From Theory to Practice. Brief. Funct. Genom. 2010, 9, 166–177. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, A.J.; Chao, S.; Asoro, F.G.; Heffner, E.L.; Hayashi, T.; Iwata, H.; Smith, K.P.; Sorrells, M.E.; Jannink, J.-L. Genomic Selection in Plant Breeding: Knowledge and Prospects. Adv. Agron. 2011, 110, 77–123. [Google Scholar]
- Heslot, N.; Yang, H.; Sorrells, M.E.; Jannink, J. Genomic Selection in Plant Breeding: A Comparison of Models. Crop Sci. 2012, 52, 146–160. [Google Scholar] [CrossRef]
- Nakaya, A.; Isobe, S.N. Will Genomic Selection Be a Practical Method for Plant Breeding? Ann. Bot. 2012, 110, 1303–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of Total Genetic Value Using Genome-Wide Dense Marker Maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, Y.; Hu, Z.; Xu, C. Genomic Selection Methods for Crop Improvement: Current Status and Prospects. Crop J. 2018, 6, 330–340. [Google Scholar] [CrossRef]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; de los Campos, G.; Burgueño, J.; González-Camacho, J.M.; Pérez-Elizalde, S.; Beyene, Y.; et al. Genomic Selection in Plant Breeding: Methods, Models, and Perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef] [PubMed]
- Dreher, K.; Morris, M.; Khairallah, M. Cost-Effective Compared with Conventional Plant Breeding Methods? In Economic and Social Issues in Agricultural Biotechnology; CABI Publishing: New York, NY, USA, 2002; p. 203. [Google Scholar]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted Mutagenesis, Precise Gene Editing, and Site-Specific Gene Insertion in Maize Using Cas9 and Guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Li, P.; Zou, C.; Lu, Y.; Xie, C.; Zhang, X.; Prasanna, B.M.; Olsen, M.S. Enhancing Genetic Gain in the Era of Molecular Breeding. J. Exp. Bot. 2017, 68, 2641–2666. [Google Scholar] [CrossRef] [Green Version]
- Beyene, Y.; Dhugga, K.; Gowda, M.; Gichuki, S.; Mugo, S.N.; Karanja, J.; Murithi, A.; Ogugo, V.; Olsen, M.; Pixley, K.; et al. Genome Editing for Tolerance to Maize Lethal Necrosis; KALRO: Nairobi, Kenya, 2021. [Google Scholar]
- Dhugga, K.S. Gene Editing for Accelerated Crop Breeding. Front. Plant Sci. 2022, 13, 1763. [Google Scholar] [CrossRef]
- Duan, X.; Ma, W.; Jiao, Z.; Tian, Y.; Ismail, R.G.; Zhou, T.; Fan, Z. Reverse Transcription-Recombinase-Aided Amplification and CRISPR/Cas12a-Based Visual Detection of Maize Chlorotic Mottle Virus. Phytopathol. Res. 2022, 4, 23. [Google Scholar] [CrossRef]
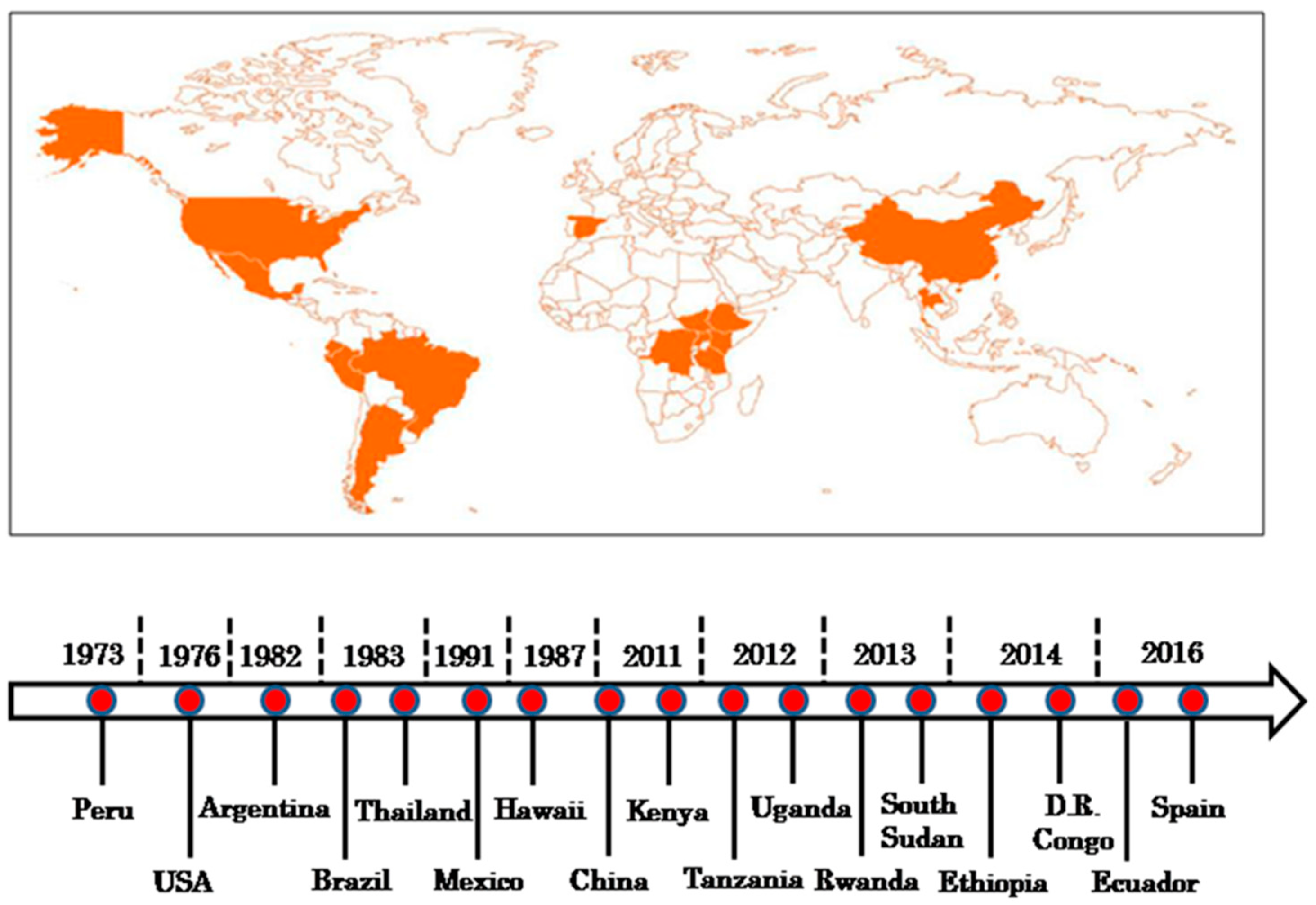


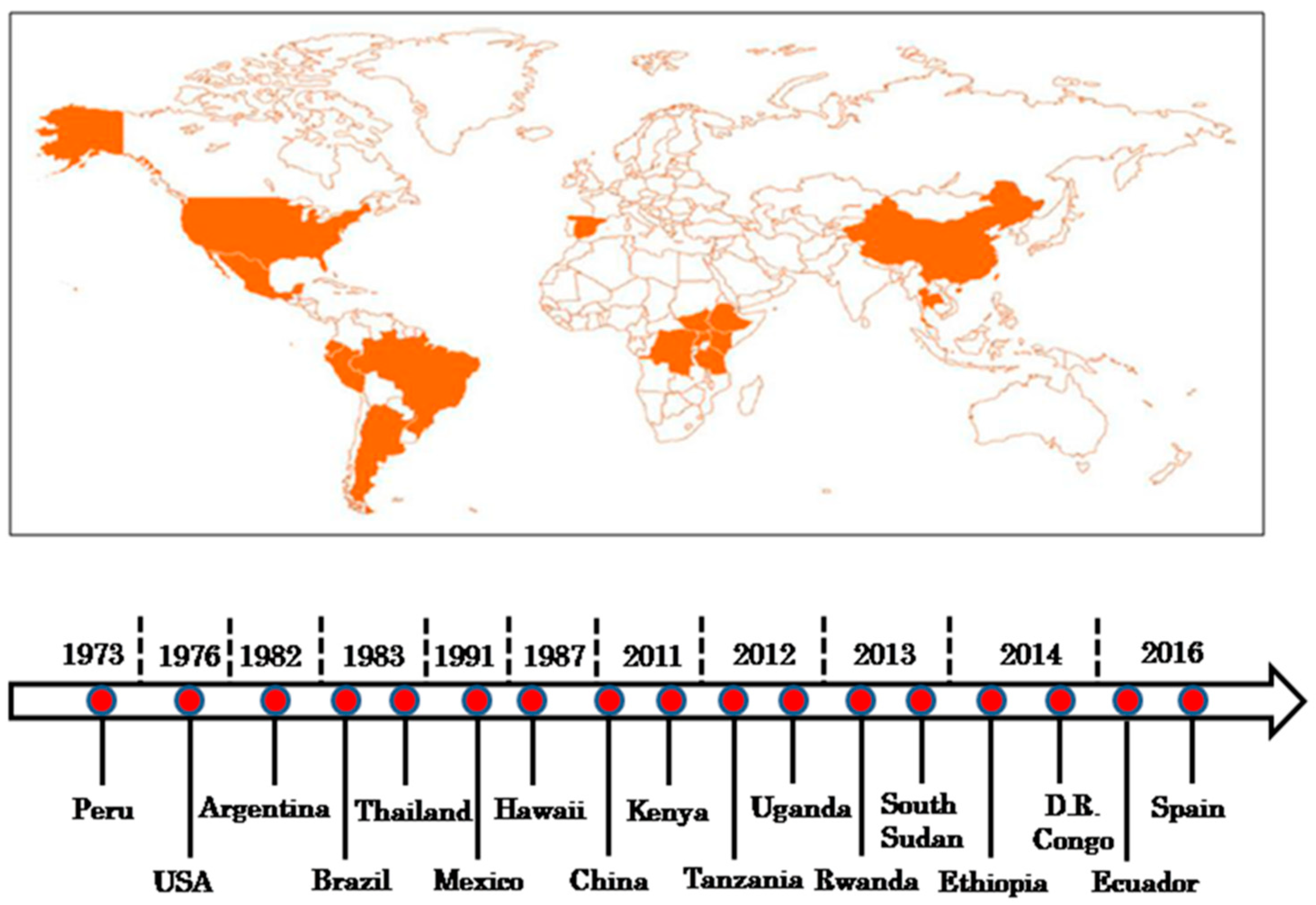{kind=link}
{kind=link}
| No. | Loci Names (QTLs or SNPs) | Genetic Approaches | Chromosomes | Positions (cM/Mb) | Lines/Populations | Virus | Genetic Effects | References |
|---|---|---|---|---|---|---|---|---|
| 1 | qMLN06_157 | Linkage mapping + association mapping | Chr 6 | /155.6 /156.5 | F2 population: SG1 (KS23-5 × CZL00025), SG2 (KS23-5 × CML545), SG3 (KS23-6 × CML539), SG4 (CML494 × CZL068), SG5 (DTP-F46 × CML442). | MCMV and SCMV | 55–70% | [65] |
| 2 | Joint mapping | Chr 1, 2, 3, 5, 6, 10 | F2 populations: (KS23-5, KS23-6, N211, DR, and Oh1VI) × Oh28 (S) RIL: Oh1VI × Oh28 (S) | MCMV | 9~78% | [66] | ||
| 3 4 5 6 | qMCMV3-108 qMLN3-108 qMCMV6-17 qMLN6-17 | GWAS+Linkage mapping + GP | Chr 3 Chr 3 Chr 6 Chr 6 | 39/ 39/ 38/ 38/ | 380 lines of Association Mapping Panel; Three DH populations | MCMV and MLN | 49.87% (DS) + 58.70% (AUDPC) 21.43% (DS) + 23.73% (AUDPC) 29.4% 17.6% (DS) and 22.9% (AUDPC) | [67] |
| 7 | GWAS Linkage mapping | Chr 3, 4, 5, 8 Chr 10 | 282 lines of Association Mapping Panel; RIL: B73 (S) × CML333 (R). | MCMV | 6.6–15.1% each SNP 16.3% for qMCMV | [68] | ||
| 8 9 10 11 12 13 14 | qMLN3_130 qMLN3_142 qMLN5_190 qMLN5_202 qMLN6_85 qMLN6_157 qMLN9_142 | Linkage mapping | Chr 3 Chr 3 Chr 5 Chr 5 Chr 6 Chr 6 Chr 9. | /125.1 /52.8;/68.5;/133.0 /190.6 /200.9 /85.2; /156.3 /132.7 | 306 F3 lines: pop1:CKDHL120918 × CML494, pop2: CML543 × CML494, pop3:CKDHL120918 × CML543, pop4:CKLTI0227 × CKDHL120918, pop5:CKDHL0089×CKDHL120918, pop6:CKDHL0221 × CKDHL120312, pop7:CKDHL0089 × CML494. | SCMV and MCMV | [72] | |
| 15 | 16 main effect QTL for MLN-early and 10 for MLN-late | Joint linkage association + Genome-wide prediction mapping | Chr 3 Chr 6 Chr 9 | F3 lines: Pop 1: 229 (CML543 × LaPostaSeqC7-F71-1-2-1-2-B-B-B-B) pop 2: 200 (CML543 × CML444) pop 3: 260 (CML444 × CML539) pop 4: 124 (Mo37 × CML144) | 3.9~43.8% | [78] | ||
| 16 17 18 | Scm2a, Scm2b, Scm1. | Linkage mapping | Chr 3 Chr 3 Chr6 | 23.5/ 46.6/ 27.5/ | 150 F2:3 lines from L520 (R) and L19 (S) | SCMV | 13.3%, 41.9% and 7.7% of PVE, respectively | [79] |
| 19 | GWAS | Across 10 Chrs | IMAS-AM: 380 lines; DTMA-AM: 235 lines. | MLND | 8~10% and 14–18% each, respectively. | [80] | ||
| 20 | 12 SNPs for disease severity; 10 SNPs for AUDPC | GWAS + GP | Chr 3, 4,6~10 (DS) Chr 1,2,4,6~10 (AUDPC) | 879 DH lines | MLND | [81] | ||
| 21 | Scmv1 | Linkage mapping Association mapping | Chr 6 | 177 F2 lines: F7RR/RR × F7RR/SS RILs: Zheng58(S) × Chang7-2(R), X178 (R) × HuangC(S) | SCMV | [82] | ||
| 25 26 | Scmv1, Scmv2. | Linkage mapping Association mapping | Chr 6 Chr 3 | 30.7/ 18.4/ | 121 F3 lines: F7 (S) ×FAP1360A (R) | SCMV | 15~62% | [83,84,85,86] |
| No. | Name of Candidate Gene | Gene ID or Tag | Viruses Involved | References |
|---|---|---|---|---|
| 1 | ZmTrx-h | Zm00001d035390 | SCMV | [82,85,86] |
| 2 | ZmABP1 | Zm00001d041711 | SCMV | [83,84,85] |
| 3 | metallothionein-like protein | 605018B04.x1 (UP|Q5U7K6_9POAL (Q5U7K6)) | SCMV, MDMV | [101] |
| 4 | S-adenosylmethionine synthetase | 946126A02.y1 (UP|METK_ORYSA (P46611)) | SCMV, MDMV | [101] |
| 5 | germin-like protein 4 | za72g09.b50 (UP|O49000_ORYSA (O49000)) | SCMV, MDMV | [101] |
| 6 | large subunit 26S ribosomal RNA | 605018B03.x1 (gb|AF036494.1|AF036494) | SCMV, MDMV | [101] |
| 7 | 14-3-3- like protein GF14-6 | Zm06_09h07_R (UP|14331_MAIZE (P49106)) | SCMV, MDMV | [101] |
| 8 | ZmPDIL-1 | GRMZM2G091481_P01 | MCMV, SCMV (susceptible) | [96,97] |
| 9 | ZmPrx5 | GRMZM2G036921_P01 | MCMV | [97] |
| 10 | ZmElc | KJ811537/Zm00001d037277 | SCMV (susceptible), MCMV (resistant) | [94] |
| 11 | SceIF4Ea/SceIF4Eb/SceIF(iso)4E | MW547070/MW547071/MW547072 | MCMV (susceptible) | [94,95] |
| 12 | ScnCBP | KX757019 | MCMV (susceptible) | [94,95] |
| 13 | ZmTGL | NP 001149280 | SCMV | [103] |
| 14 | ZmPAL | Zm00001d017274 | SCMV | [104] |
| 15 | ZmDWF4 | Zm00001d028325 | MCMV | [105] |
| 16 | ZmNR | Zm00001d049995 | MCMV | [105] |
| 17 | ZmARF3/30 | Zm00001d001879/Zm00001d026590 | MCMV (susceptible) | [98] |
| 18 | ZmPAO1 | Zm00001d024281 | MCMV (susceptible) | [98] |
| 19 | ZmCAT1/2/3 | NM_001254879.2/NM_001111840.2/NM_001363892.1 | MCMV | [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnmark, O.; Indieka, S.; Liu, G.; Gowda, M.; Suresh, L.M.; Zhang, W.; Gao, X. Fighting Death for Living: Recent Advances in Molecular and Genetic Mechanisms Underlying Maize Lethal Necrosis Disease Resistance. Viruses 2022, 14, 2765. https://doi.org/10.3390/v14122765
Johnmark O, Indieka S, Liu G, Gowda M, Suresh LM, Zhang W, Gao X. Fighting Death for Living: Recent Advances in Molecular and Genetic Mechanisms Underlying Maize Lethal Necrosis Disease Resistance. Viruses. 2022; 14(12):2765. https://doi.org/10.3390/v14122765
Chicago/Turabian StyleJohnmark, Onyino, Stephen Indieka, Gaoqiong Liu, Manje Gowda, L. M. Suresh, Wenli Zhang, and Xiquan Gao. 2022. "Fighting Death for Living: Recent Advances in Molecular and Genetic Mechanisms Underlying Maize Lethal Necrosis Disease Resistance" Viruses 14, no. 12: 2765. https://doi.org/10.3390/v14122765