
The Virus-Induced Transcription Factor SHE1 Interacts with and Regulates Expression of the Inhibitor of Virus Replication (IVR) in N Gene Tobacco

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Virus Materials
2.2. Plant Culture and Virus Inoculation
2.3. Plasmid Construction and Plant Transformation
2.4. RNA Extraction, RT-PCR, RT-qPCR and Northern Blot Hybridization Analyses
2.5. Characterization of Transgenic Plants
2.6. Protein Extraction, Co-Immunoprecipitation and Western Blot Analysis
2.7. Yeast Two-Hybrid and X-Gal Assays
2.8. BiFC and Duolink In Situ PLA
3. Results
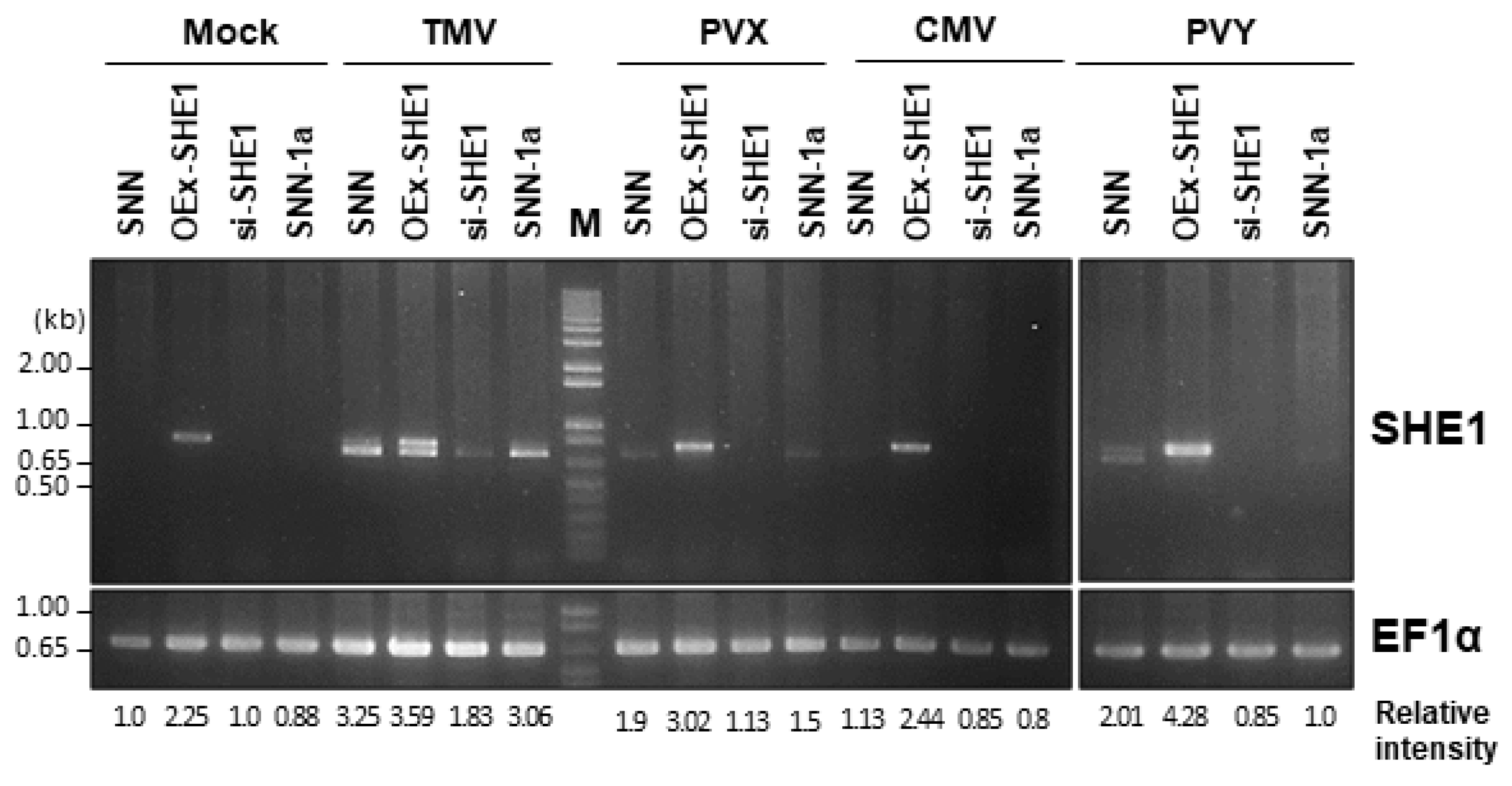
3.1. Expression of SHE1 in Various Transgenic Lines: Effect of Virus Infection
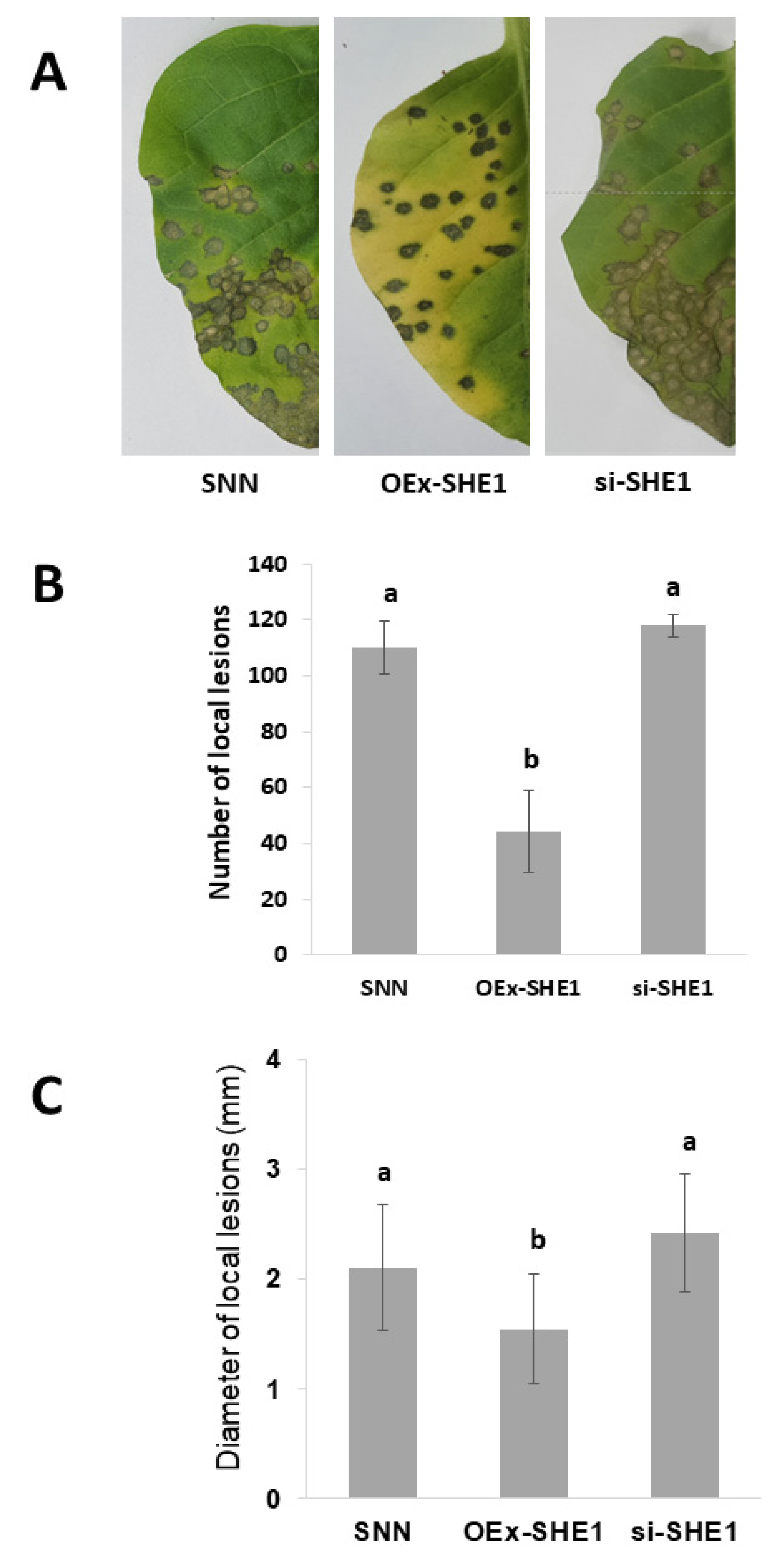
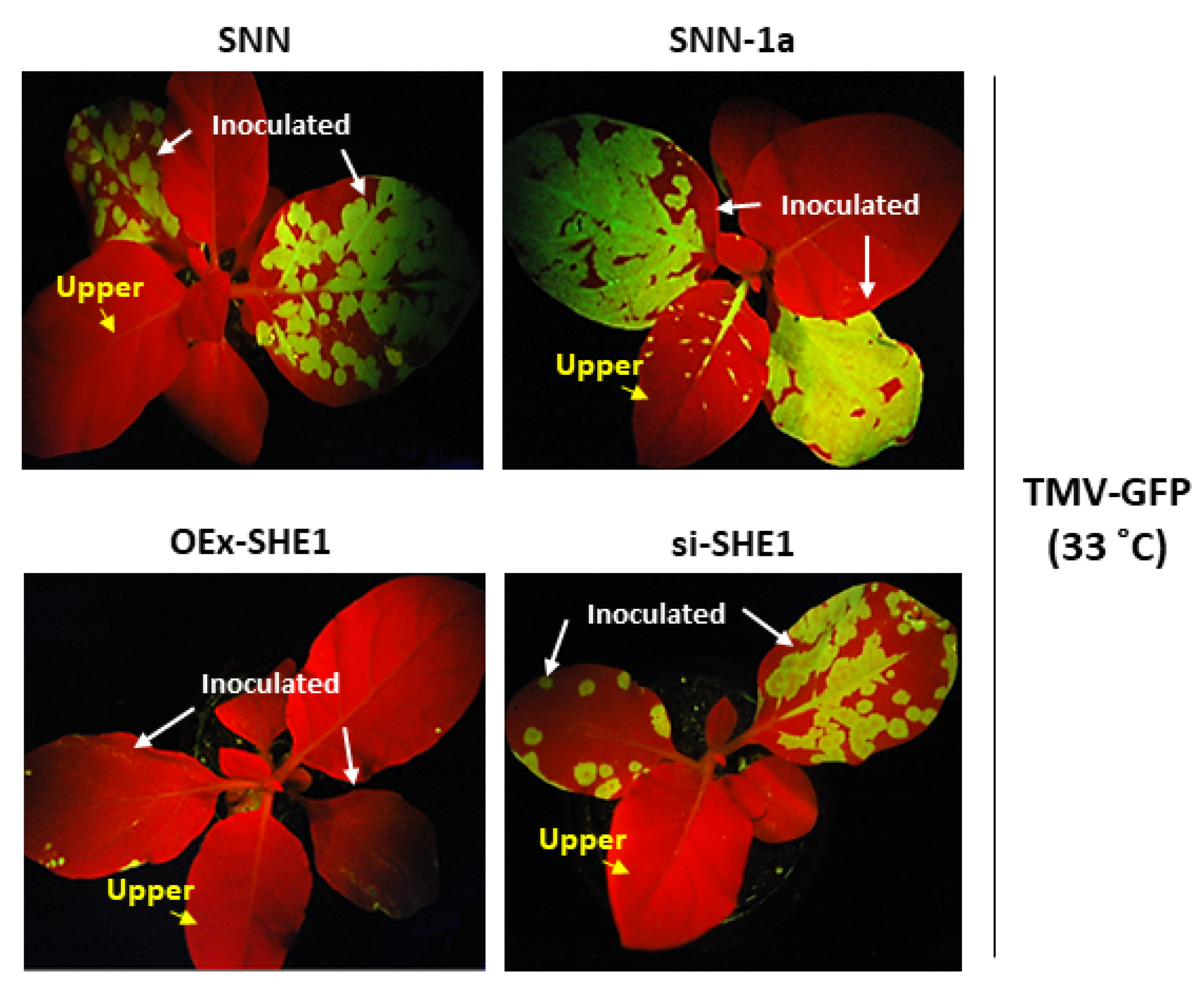
3.2. Effect of Silencing or Over-Expressing SHE1 on Movement of TMV
3.3. Effect of Silencing or Over-Expressing SHE1 on IVR mRNA Accumulation
3.4. Interaction between SHE1 and IVR
3.5. Co-Localization of SHE1 and IVR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carr, J.P.; Lewsey, M.G.; Palukaitis, P. Signaling in induced resistance. Adv. Virus Res. 2010, 76, 57–121. [Google Scholar] [PubMed]
- Carr, J.P.; Murphy, A.M.; Tungadi, T.; Yoon, J.-Y. Plant defense signals: Players and pawns in plant-virus-vector interactions. Plant Sci. 2019, 279, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, A.; Carrington, J.C. Antiviral roles of plant ARGONAUTES. Curr. Opin. Plant Biol. 2015, 27, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Palukaitis, P.; Yoon, J.-Y. R gene mediated defense against viruses. Curr. Opin. Virol. 2020, 45, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Leibman, D.; Kravchik, M.; Wolf, D.; Haviv, S.; Weissberg, M.; Ophir, R.; Paris, H.S.; Palukaitis, P.; Ding, S.-W.; Gaba, V.; et al. Differential expression of cucumber RNA-dependent RNA polymerase 1 genes during antiviral defence and resistance. Mol. Plant Pathol. 2018, 19, 300–312. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C.; Van Strien, E.S. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Loebenstein, G. Local lesions and induced resistance. Adv. Virus Res. 2009, 75, 73–117. [Google Scholar]
- Garcia-Ruiz, H. Host factors against plant viruses. Mol. Plant Pathol. 2019, 20, 1588–1601. [Google Scholar] [CrossRef]
- Alazem, M.; Lin, N.-S. Roles of plant hormones in the regulation of host–virus interactions. Mol. Plant Pathol. 2015, 16, 529–540. [Google Scholar] [CrossRef]
- Alazem, M.; Lin, N.-S. Antiviral roles of abscisic acid in plants. Front. Plant Sci. 2017, 8, 1760. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The crosstalks between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef] [PubMed]
- Hunter, L.J.R.; Westwood, J.H.; Heath, G.; Macaulay, K.; Smith, A.G.; MacFarlane, S.A.; Palukaitis, P.; Carr, J.P. Regulation of RNA-dependent RNA polymerase 1 and Isochorismate Synthase gene expression in Arabidopsis. PLoS ONE 2013, 8, e66530. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandehroo, F.; Takeshita, M.; Squires, J.; Palukaitis, P. The influence of RNA-dependent RNA polymerase 1 on Potato virus Y infection and on other antiviral response genes. Mol. Plant-Microbe Interact. 2009, 22, 1312–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilliland, A.; Singh, D.P.; Hayward, J.M.; Moore, C.A.; Murphy, A.M.; York, C.J.; Slator, J.; Carr, J.P. Genetic modification of alternative respiration has differential effects on antimycin A-induced versus salicylic acid-induced resistance to Tobacco mosaic virus. Plant Physiol. 2003, 132, 1518–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, U.; Dröge-Laser, W. Overexpression of NtERF5, a new member of the tobacco ethylene response transcription factor family enhances resistance to Tobacco mosaic virus. Mol. Plant-Microbe Interact. 2004, 17, 1162–1171. [Google Scholar] [CrossRef] [Green Version]
- Baek, E.; Yoon, J.-Y.; Palukaitis, P. Validation of reference genes for quantifying changes in gene expression in virus-infected tobacco. Virology 2017, 510, 29–39. [Google Scholar] [CrossRef]
- Yoon, J.-Y.; Palukaitis, P. Cucumber mosaic virus 1a protein interacts with the tobacco SHE1 transcription factor and partitions between the nucleus and the tonoplast membrane. Plant Pathol. J. 2021, 37, 182–193. [Google Scholar] [CrossRef]
- Akad, F.; Teverovsky, E.; David, A.; Czosnek, H.; Gidoni, D.; Gera, A.; Loebenstein, G. A cDNA from tobacco codes for an inhibitor of virus replication (IVR)-like protein. Plant Mol. Biol. 1999, 40, 969–976. [Google Scholar] [CrossRef]
- Loebenstein, G.; Gera, A. Inhibitor of virus replication released from tobacco mosaic virus-infected protoplasts of a local lesion-responding tobacco cultivar. Virology 1981, 114, 132–139. [Google Scholar] [CrossRef]
- Gera, A.; Loebenstein, G. Further studies of an inhibitor of virus replication from tobacco mosaic virus-infected protoplasts of a local lesion-responding tobacco cultivar. Phytopathology 1983, 73, 111–115. [Google Scholar] [CrossRef]
- Loebenstein, G.; Akad, F. The local lesion response. In Natural Resistance Mechanisms of Plants to Viruses; Loebenstein, G., Carr, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 99–124. [Google Scholar]
- Spiegel, S.; Gera, A.; Salomon, R.; Ahl, P.; Harlap, S.; Loebenstein, G. Recovery of an inhibitor of virus replication from the intercellular fluid of hypersensitive tobacco infected with tobacco mosaic virus and from uninfected induced-resistant tissue. Phytopathology 1989, 79, 258–262. [Google Scholar] [CrossRef]
- Akad, A.; Teverovsky, E.; Gidoni Elad, D.Y.; Kirschner, B.; Rav-David, D.; Czosnek, H.; Loebenstein, G. Resistance to Tobacco mosaic virus and Botrytis cinerea in tobacco transformed with complementary DNA encoding an inhibitor of viral replication-like protein. Ann. Appl. Biol. 2005, 147, 89–100. [Google Scholar] [CrossRef]
- Canto, T.; Palukaitis, P. Transgenically expressed cucumber mosaic virus RNA 1 simultaneously complements replication of cucumber mosaic virus RNAs 2 and 3 and confers resistance to systemic infection. Virology 1998, 250, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canto, T.; Palukaitis, P. Novel N gene-associated, temperature-independent resistance to the movement of Tobacco mosaic virus vectors neutralized by a Cucumber mosaic virus RNA1 transgene. J. Virol. 2002, 76, 12908–12916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerschen, A.; Napoli, C.A.; Jorgensen, R.A.; Müller, A.E. Effectiveness of RNA interference in transgenic plants. FEBS Lett. 2004, 566, 223–228. [Google Scholar] [CrossRef]
- Gehl, C.; Waadt, R.; Kudla, J.; Mendel, R.R.; Hänsch, R. New GATEWAY vectors for high throughput analysis of protein-protein interactions by bimolecular fluorescence complementation. Mol. Plant 2009, 2, 1051–1058. [Google Scholar] [CrossRef] [Green Version]
- Waadt, R.; Schmidt, L.K.; Lohse, M.; Hashimoto, K.; Bock, R.; Kudla, J. Multicolor bimolecular fluorescence complementation reveals simultaneous formation of alternative CBL/CIPK complexes in planta. Plant J. 2008, 56, 505–516. [Google Scholar] [CrossRef]
- Kim, B.M.; Suehiro, N.; Natsuaki, T.; Inukai, T.; Masuta, C. The P3 protein of Turnip mosaic virus can alone induce hypersensitive response-like cell death in Arabidopsis thaliana carring TuNI. Mol. Plant-Microbe Interact. 2010, 23, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Inaba, J.; Kim, B.M.; Shimura, H.; Masuta, C. Virus-induced necrosis is a consequence of direct protein-protein interaction between a viral RNA-silencing suppressor and a host catalase. Plant Physiol. 2011, 156, 2026–2036. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol. 2008, 26, 101–106. [Google Scholar] [CrossRef]
- Kassanis, B. Some effects of high temperature on the susceptibility of plants to infection with viruses. Ann. Appl. Biol. 1952, 39, 358–369. [Google Scholar] [CrossRef]
- Takahashi, T. Studies on viral pathogenesis in plant hosts. VIII. Systemic viral invasion and localization of infection in “Samsun NN” tobacco plants resulting from tobacco mosaic virus infection. J. Phytopathol. 1975, 84, 75–87. [Google Scholar] [CrossRef]
- Kobayashi, M.; Seo, S.; Hirai, K.; Yamamoto-Katou, A.; Katou, S.; Seto, H.; Meshi, T.; Mitsuhara, I.; Ohashi, Y. Silencing of WIPK and SIPK mitogen-activated protein kinases reduces Tobacco mosaic virus accumulation but permits systemic viral movement in tobacco possessing the N resistance gene. Mol. Plant-Microbe Interact. 2010, 23, 1032–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, A.; Zaitlin, M.; Sehgal, O.P. The isolation of defective tobacco mosaic virus strains. Proc. Natl. Acad. Sci. USA 1962, 48, 1845–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mur, L.A.J.; Bi, Y.-M.; Darby, R.M.; Firek, S.; Draper, J. Compromising early salicylic acid accumulation delays the hypersensitive response and increases viral dispersal during lesion establishment in TMV-infected tobacco. Plant J. 1997, 12, 1113–1126. [Google Scholar] [CrossRef]
- Gera, A.; Tam, Y.; Teverovsky, E.; Loebenstein, G. Enhanced tobacco mosaic virus production and suppressed synthesis of the inhibitor of virus replication in protoplasts and plants of local lesion responding cultivars exposed to 35 °C. Physiol. Mol. Plant Pathol. 1993, 43, 299–306. [Google Scholar] [CrossRef]
- Loebenstein, G.; Gera, A.; Gianinazzi, S. Constitutive production of an inhibitor of virus replication in the interspecific hybrid of Nicotiana glutinosa × Nicotiana debneyi. Physiol. Mol. Plant Pathol. 1990, 37, 145–151. [Google Scholar] [CrossRef]
- Palukaitis, P.; Akbarimotlagh, M.; Baek, E.; Yoon, J.-Y. The secret life of the inhibitor of virus replication. Viruses 2022, 14, 2782. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, J.-Y.; Baek, E.; Kim, M.; Palukaitis, P. The Virus-Induced Transcription Factor SHE1 Interacts with and Regulates Expression of the Inhibitor of Virus Replication (IVR) in N Gene Tobacco. Viruses 2023, 15, 59. https://doi.org/10.3390/v15010059
Yoon J-Y, Baek E, Kim M, Palukaitis P. The Virus-Induced Transcription Factor SHE1 Interacts with and Regulates Expression of the Inhibitor of Virus Replication (IVR) in N Gene Tobacco. Viruses. 2023; 15(1):59. https://doi.org/10.3390/v15010059
Chicago/Turabian StyleYoon, Ju-Yeon, Eseul Baek, Mira Kim, and Peter Palukaitis. 2023. "The Virus-Induced Transcription Factor SHE1 Interacts with and Regulates Expression of the Inhibitor of Virus Replication (IVR) in N Gene Tobacco" Viruses 15, no. 1: 59. https://doi.org/10.3390/v15010059