
Huaier Polysaccharide Interrupts PRV Infection via Reducing Virus Adsorption and Entry

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Reagents and Antibodies
2.3. Cell-Based Infectivity Assays
2.3.1. Cells Viability Assay
2.3.2. General Effect of Huaier Polysaccharide on PRV Infection
2.3.3. Exploration of Huaier Polysaccharide’s Effect on PRV Adsorption and Entry
2.3.4. General Effect of Huaier Polysaccharide on PRV Adsorption
2.3.5. Exploration of Huaier Polysaccharide’s Effect on PRV Entry
2.3.6. Effect of Huaier Polysaccharide on PRV Replication
2.3.7. Effect of Huaier Polysaccharide on PRV XJ5
2.3.8. Western Blotting
2.3.9. Virus Titer Assays
2.3.10. Indirect Immunofluorescent Assay
2.3.11. DNA Extraction and qRT-PCR
2.3.12. Electron Microscopy
2.3.13. Flow Cytometry Assay
2.3.14. Statistical Analysis
3. Results
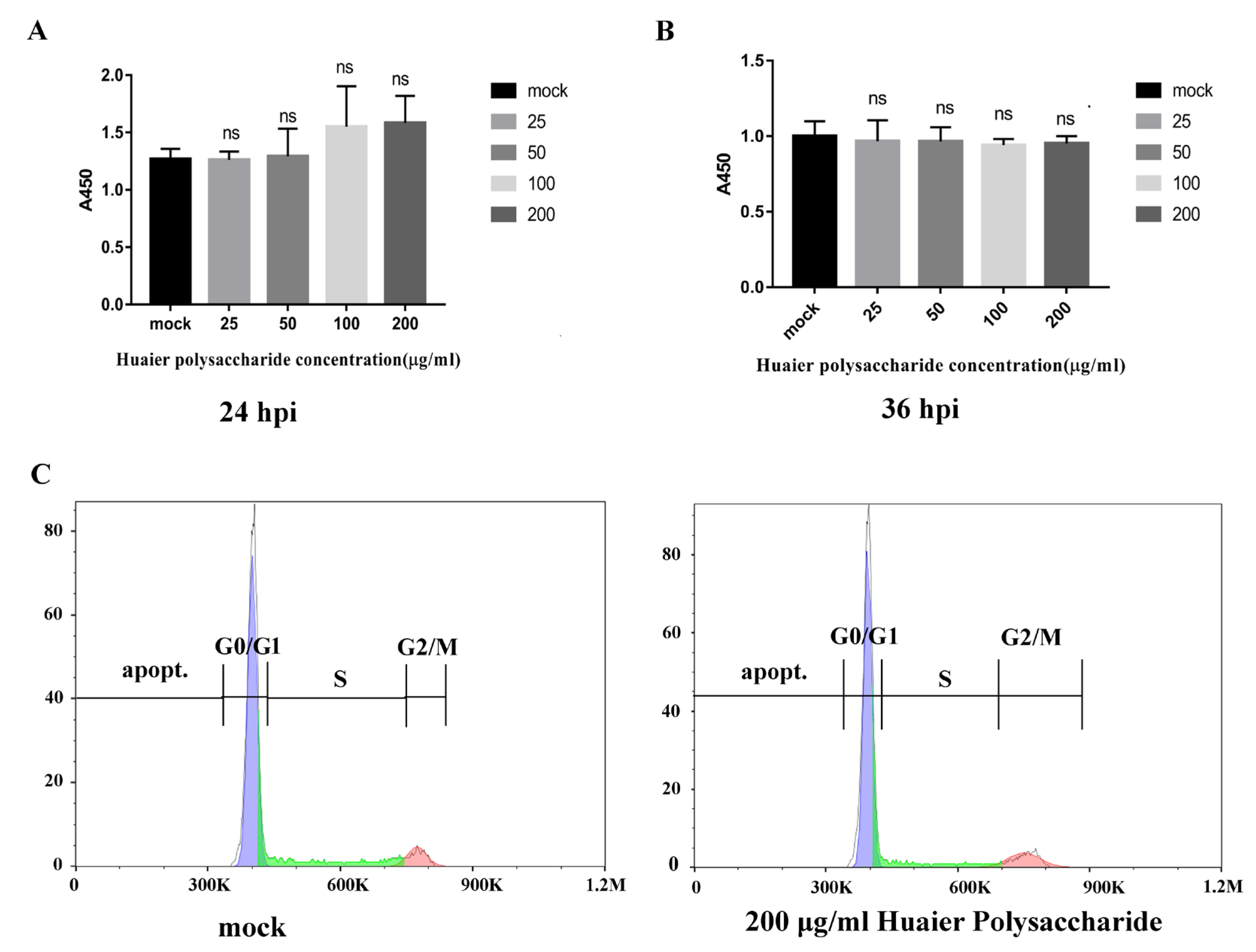
3.1. Huaier Polysaccharide Has No Inhibitory Effect on PK15 Cells’ Growth and Apoptosis
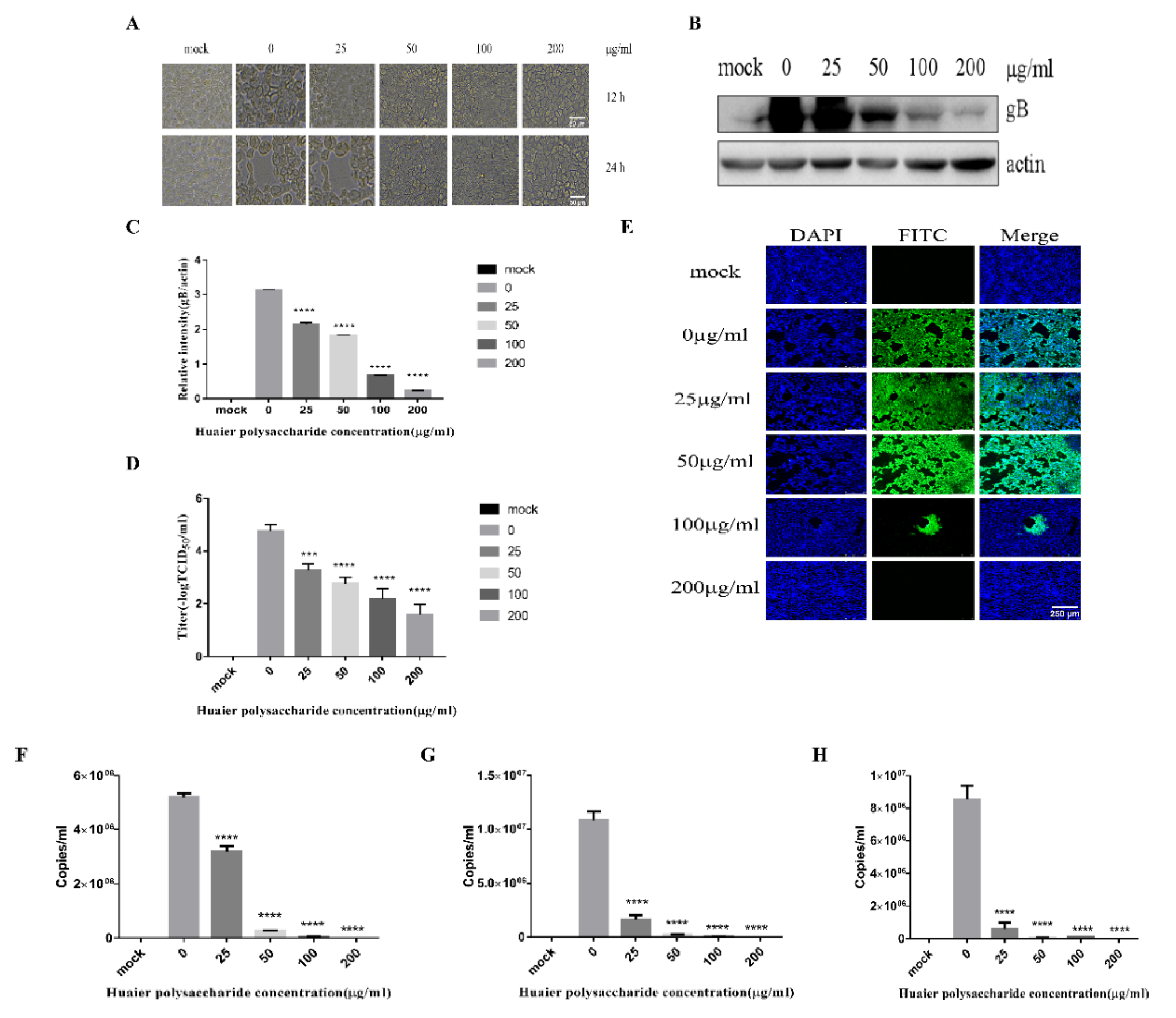
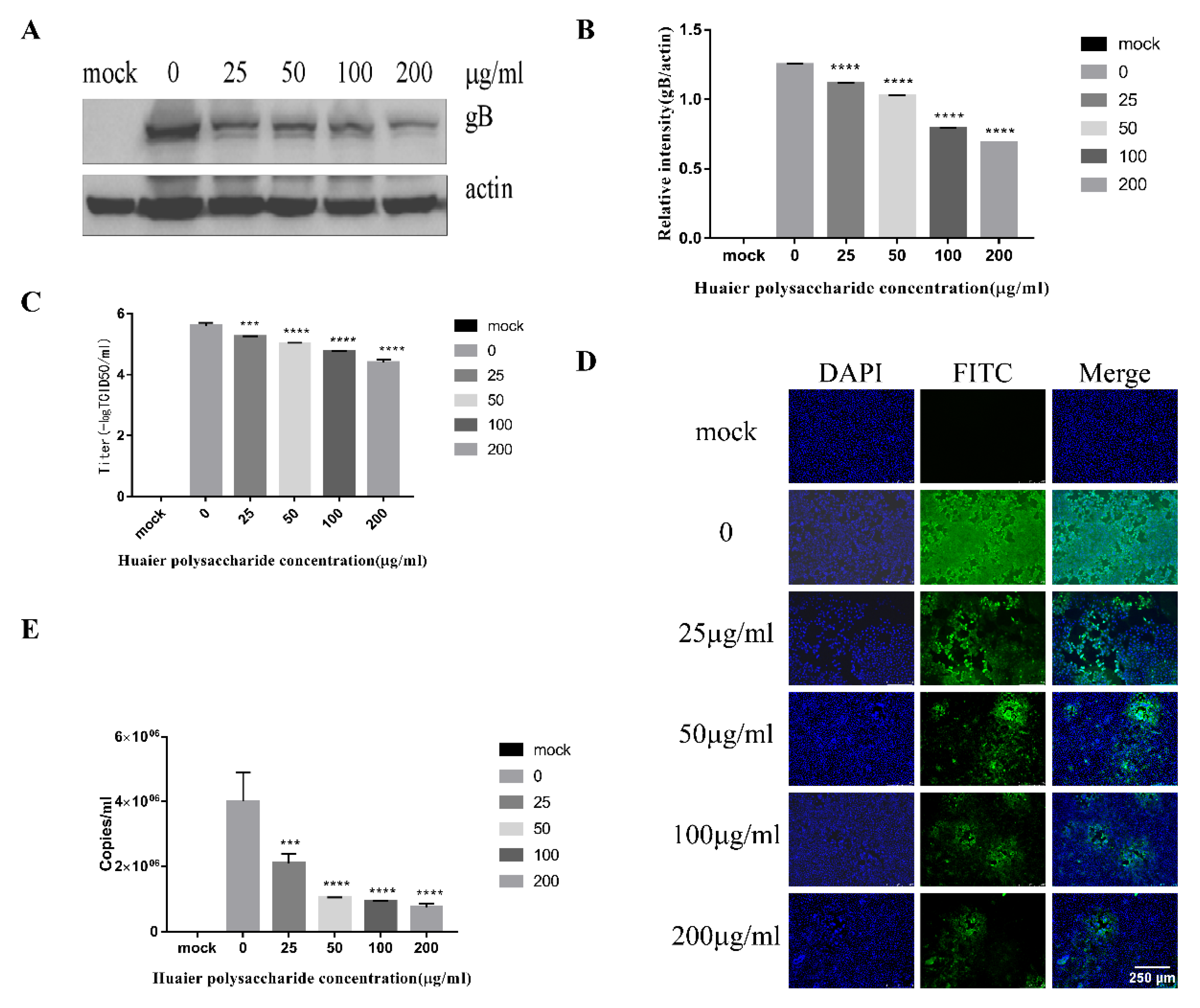
3.2. Huaier Polysaccharide Inhibis PRV XJ5 Infection in PK15 Cells
3.3. Huaier Polysaccharide Decreased the Adsorption and Entry of PRV XJ5
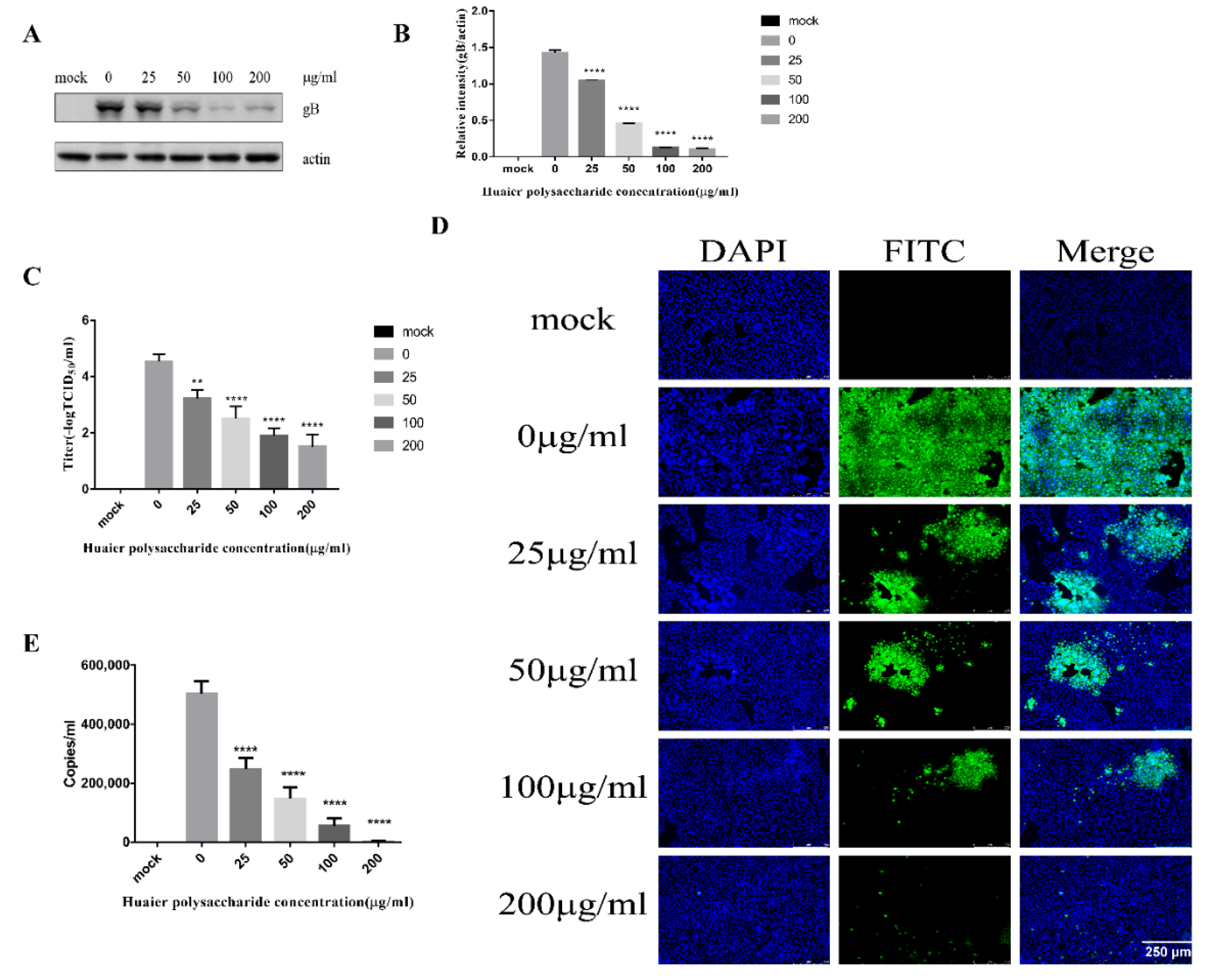
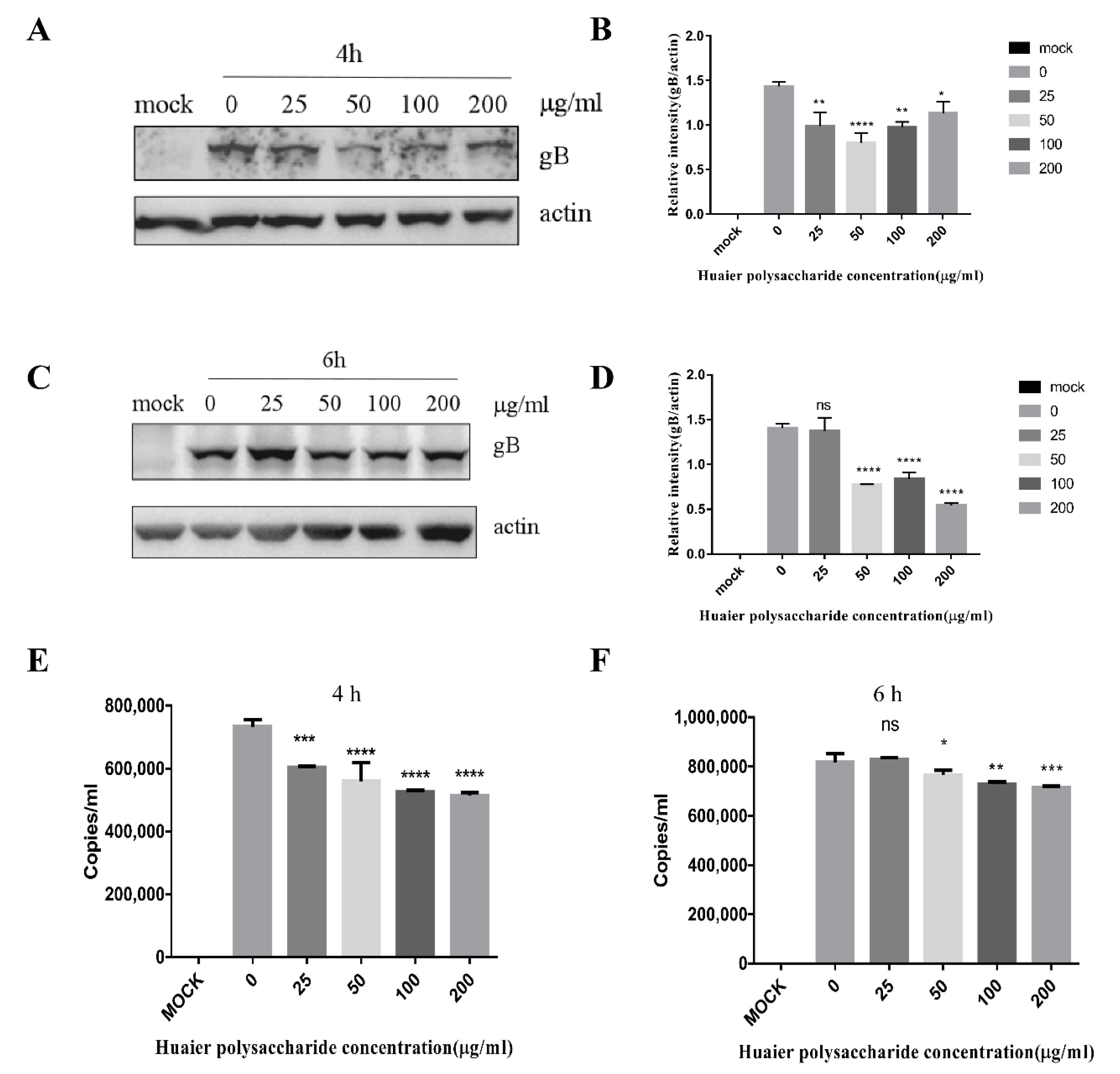
3.3.1. Huaier Polysaccharide Attenuates PRV Adsorption
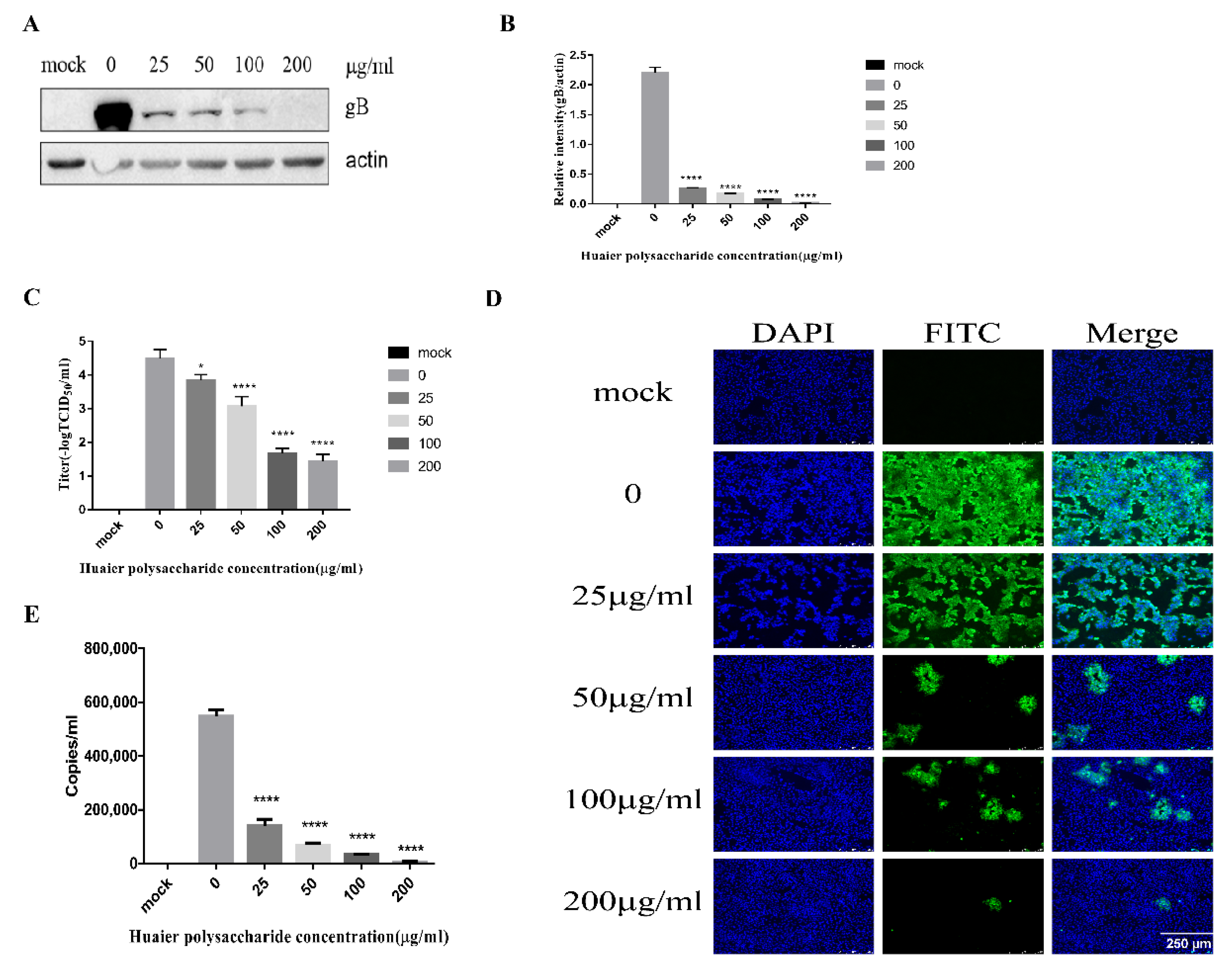
3.3.2. Huaier Polysaccharide Influences PRV Entry into PK15 Cells
3.4. Huaier Polysaccharide Mildly Reduces PRV XJ5 Replication in PK15 Cells
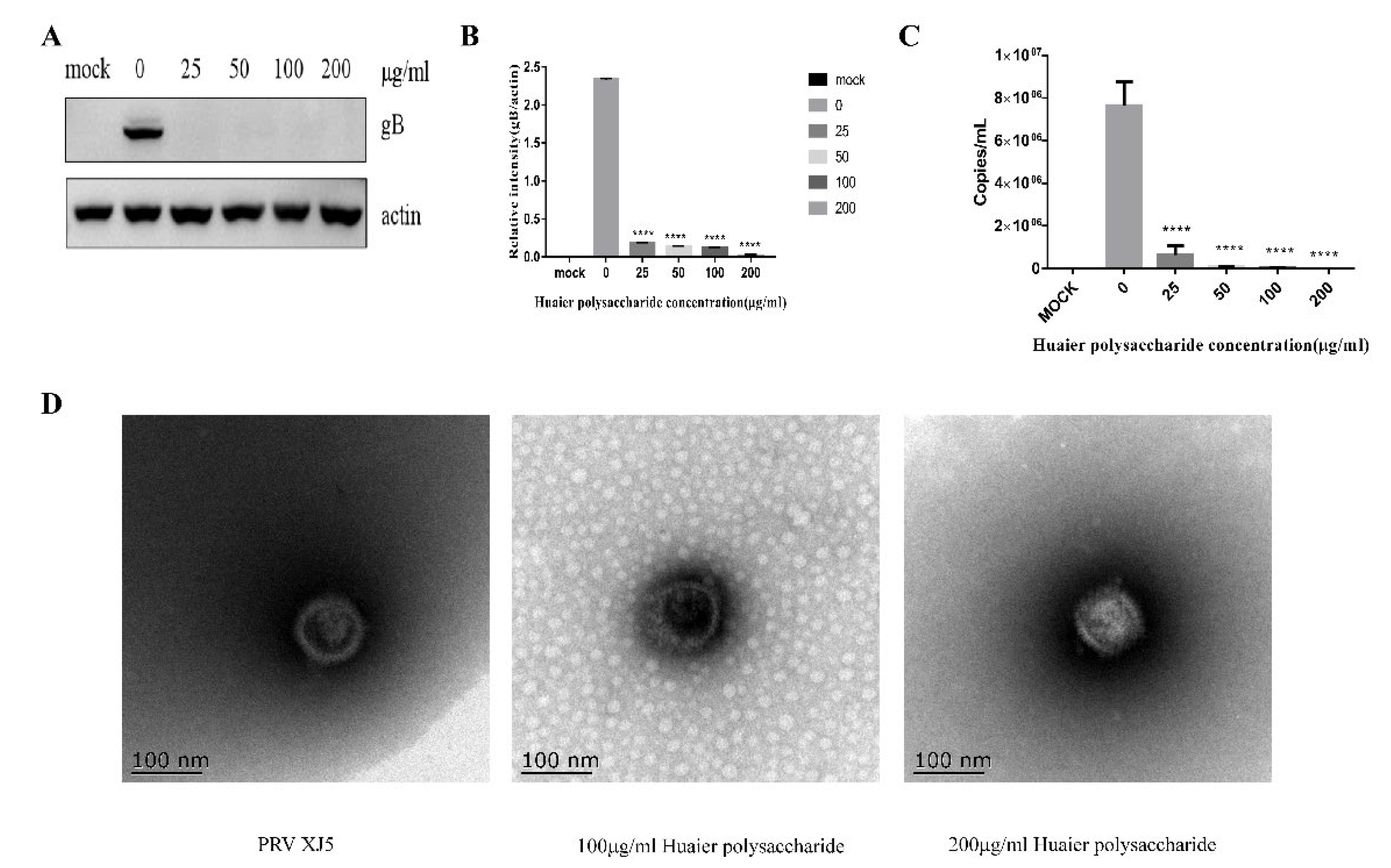
3.5. Huaier Polysaccharide May Directly Inactivate PRV XJ5
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lee, J.Y.; Wilson, M.R. A review of pseudorabies (Aujeszky’s disease) in pigs. Can. Vet. J. 1979, 20, 65–69. [Google Scholar] [PubMed]
- Xia, L.M.; Sun, Q.Y.; Wang, J.J.; Chen, Q.; Liu, P.H.; Shen, C.J.; Sun, J.H.; Tu, Y.P.; Shen, S.F.; Zhu, J.C.; et al. Epidemiology of pseudorabies in intensive pig farms in Shanghai, China: Herd-level prevalence and risk factors. Prev. Vet. Med. 2018, 159, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Luo, Y.Z.; Wang, C.-H.; Yuan, J.; Li, N.; Song, K.; Qiu, H.-J. Control of swine pseudorabies in China: Opportunities and limitations. Vet. Microbiol. 2016, 183, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Klupp, B.G. Pseudorabies Virus Infections. Pathogens 2021, 10, 719. [Google Scholar] [CrossRef]
- Minamiguchi, K.; Kojima, S.; Sakumoto, K.; Kirisawa, R. Isolation and molecular characterization of a variant of Chinese gC-genotype II pseudorabies virus from a hunting dog infected by biting a wild boar in Japan and its pathogenicity in a mouse model. Virus Genes 2019, 55, 322–331. [Google Scholar] [CrossRef]
- Cheng, Z.; Kong, Z.; Liu, P.; Fu, Z.; Zhang, J.; Liu, M.; Shang, Y. Natural infection of a variant pseudorabies virus leads to bovine death in China. Transbound. Emerg. Dis. 2020, 67, 518–522. [Google Scholar] [CrossRef]
- Kong, H.; Zhang, K.; Liu, Y.; Shang, Y.; Wu, B.; Liu, X. Attenuated live vaccine (Bartha-K16) caused pseudorabies (Aujeszky’s disease) in sheep. Vet. Res. Commun. 2013, 37, 329–332. [Google Scholar] [CrossRef]
- Kaneko, C.; Kaneko, Y.; Sudaryatma, P.E.; Mekata, H.; Kirino, Y.; Yamaguchi, R.; Okabayashi, T. Pseudorabies virus infection in hunting dogs in Oita, Japan: Report from a prefecture free from Aujeszky’s disease in domestic pigs. J. Vet. Med. Sci. 2021, 83, 680–684. [Google Scholar] [CrossRef]
- Jin, H.-L.; Gao, S.-M.; Liu, Y.; Zhang, S.-F.; Hu, R.-L. Pseudorabies in farmed foxes fed pig offal in Shandong province, China. Arch. Virol. 2016, 161, 445–448. [Google Scholar] [CrossRef]
- Freuling, C.M.; Mueller, T.F.; Mettenleiter, T.C. Vaccines against pseudorabies virus (PrV). Vet. Microbiol. 2017, 206, 3–9. [Google Scholar] [CrossRef]
- Mettenleiter, T.C. Aujeszky’s disease (pseudorabies) virus: The virus and molecular pathogenesis—State of the art, June 1999. Vet. Res. 2000, 31, 99–115. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Yao, J.; Yang, Y.; Luo, W.; Yuan, X.; Yang, L.; Wang, A. Current Status and Challenge of Pseudorabies Virus Infection in China. Virol. Sin. 2021, 36, 588–607. [Google Scholar] [CrossRef]
- Szpara, M.L.; Tafuri, Y.R.; Parsons, L.; Shamim, S.R.; Verstrepen, K.J.; Legendre, M.; Enquist, L.W. A Wide Extent of Inter-Strain Diversity in Virulent and Vaccine Strains of Alphaherpesviruses. PLoS Pathog. 2011, 7, e1002282. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Guo, J.-C.; Gao, J.-C.; Wang, T.-Y.; Zhao, K.; Chang, X.-B.; Wang, Q.; Peng, J.-M.; Tian, Z.-J.; Cai, X.-H.; et al. Genomic analyses reveal that partial sequence of an earlier pseudorabies virus in China is originated from a Bartha-vaccine-like strain. Virology 2016, 491, 56–63. [Google Scholar] [CrossRef]
- Ai, J.-W.; Weng, S.-S.; Cheng, Q.; Cui, P.; Li, Y.-J.; Wu, H.-L.; Zhu, Y.-M.; Xu, B.; Zhang, W.-H. Human Endophthalmitis Caused by Pseudorabies Virus Infection, China, 2017. Emerg. Infect. Dis. 2018, 24, 1087–1090. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, X.; Xie, C.; Ding, S.; Yang, H.; Guo, S.; Li, J.; Qin, L.; Ban, F.; Wang, D.; et al. A novel human acute encephalitis caused by pseudorabies virus variant strain. Clin. Infect. Dis. 2020, 73, e3690–e3700. [Google Scholar] [CrossRef]
- Yang, X.; Guan, H.; Li, C.; Li, Y.; Wang, S.; Zhao, X.; Zhao, Y.; Liu, Y. Characteristics of human encephalitis caused by pseudorabies virus: A case series study. Int. J. Infect. Dis. 2019, 87, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.-H.; Chuang, W.-C.; Lam, W.; Jiang, Z.; Cheng, Y.-C. Safety Surveillance of Traditional Chinese Medicine: Current and Future. Drug Saf. 2015, 38, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Kannan, S.; Kolandaivel, P. Antiviral potential of natural compounds against influenza virus hemagglutinin. Comput. Biol. Chem. 2017, 71, 207–218. [Google Scholar] [CrossRef]
- Zhang, N.; Kong, X.; Yan, S.; Yuan, C.; Yang, Q. Huaier aqueous extract inhibits proliferation of breast cancer cells by inducing apoptosis. Cancer Sci. 2010, 101, 2375–2383. [Google Scholar] [CrossRef]
- Kong, X.; Ding, X.; Yang, Q. Identification of multi-target effects of Huaier aqueous extract via microarray profiling in triple-negative breast cancer cells. Int. J. Oncol. 2015, 46, 2047–2056. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Hu, X.; Xiong, H.; Qiu, H.; Yuan, X.; Zhu, F.; Wang, Y.; Zou, Y. A polysaccharide from Huaier induced apoptosis in MCF-7 breast cancer cells via down-regulation of MTDH protein. Carbohydr. Polym. 2016, 151, 1027–1033. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Shu, C.; Laurence, A.D.; Chen, Y.; Peng, B.G.; Zhen, Z.J.; Cai, J.Q.; Ding, Y.T.; Li, L.Q.; Zhang, Y.B.; et al. Effect of Huaier granule on recurrence after curative resection of HCC: A multicentre, randomised clinical trial. Gut 2018, 67, 2006–2016. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Wu, W.; Qu, K.; Xi, W. Traditional Chinese biomedical preparation (Huaier Granule) for breast cancer: A PRISMA-compliant meta-analysis. Biosci. Rep. 2020, 40, BSR20202509. [Google Scholar] [CrossRef]
- Wang, M.; Hu, Y.; Hou, L.; Pan, Q.; Tang, P.; Jiang, J. A clinical study on the use of Huaier granules in post-surgical treatment of triple-negative breast cancer. Gland Surg. 2019, 8, 758–765. [Google Scholar] [CrossRef]
- Enquist, L.W.; Husak, P.J.; Banfield, B.W.; Smith, G.A. Infection and spread of alphaherpesviruses in the nervous system. Adv. Virus Res. 1998, 51, 237–347. [Google Scholar]
- Mettenleiter, T.C. Budding events in herpesvirus morphogenesis. Virus Res. 2004, 106, 167–180. [Google Scholar] [CrossRef]
- Ren, J.; Wang, H.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. Glycoproteins C and D of PRV Strain HB1201 Contribute Individually to the Escape From Bartha-K61 Vaccine-Induced Immunity. Front. Microbiol. 2020, 11, 323. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Zhai, X.; Su, J.; Ye, R.; Zheng, Y.; Su, S. Antiviral Activity of Germacrone against Pseudorabies Virus in Vitro. Pathogens 2019, 8, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Xie, F.-j.; Liu, S.; Yao, C.-y.; Liu, W.-h.; Cai, G.-q.; Liao, G.-q. Huaier Granule Combined with Tegafur Gimeracil Oteracil Potassium Promotes Stage IIb Gastric Cancer Prognosis and Induces Gastric Cancer Cell Apoptosis by Regulating Livin. Biomed. Res. Int. 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Zhao, Y.; Wang, Y.; Zha, X.; Zhao, Y.; Tu, P.; Hu, Z. Huaier suppresses proliferative and metastatic potential of prostate cancer PC3 cells via downregulation of Lamin B1 and induction of autophagy. Oncol. Rep. 2018, 39, 3055–3063. [Google Scholar]
- Wang, Y.; Lv, H.; Xu, Z.; Sun, J.; Ni, Y.; Chen, Z.; Cheng, X. Huaier n-butanol extract suppresses proliferation and metastasis of gastric cancer via c-Myc-Bmi1 axis. Sci. Rep. 2019, 9, 447. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Liu, X.; Yin, A.; Wei, Y.; Yang, Q.; Kong, B. Huaier aqueous extract inhibits cervical cancer cell proliferation via JNK/p38 pathway. Int. J. Oncol. 2015, 47, 1054–1060. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Yang, A.; Su, G.; Zhao, Y.; Wang, Y.; Chai, X.; Tu, P. Huaier restrains proliferative and invasive potential of human hepatoma SKHEP-1 cells partially through decreased Lamin B1 and elevated NOV. Sci. Rep. 2016, 6, 31298. [Google Scholar] [CrossRef]
- Xie, H.-X.; Xu, Z.-Y.; Tang, J.-N.; Du, Y.-A.; Huang, L.; Yu, P.-F.; Cheng, X.-D. Effect of Huaier on the proliferation and apoptosis of human gastric cancer cells through modulation of the PI3K/AKT signaling pathway. Exp. Ther. Med. 2015, 10, 1212–1218. [Google Scholar] [CrossRef] [Green Version]
- Heller, M.J. DNA microarray technology: Devices, systems, and applications. Annu. Rev. Biomed. Eng. 2002, 4, 129–153. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Tang, Y.; Zhou, R.; Shi, X.; Zhang, H.; Liu, T.; Lian, Z.; Shi, X. Bi-directional solid fermentation products of Trametes robiniophila Murr with Radix Isatidis inhibit proliferation and metastasis of breast cancer cells. J. Chin. Med. Assoc. 2018, 81, 520–530. [Google Scholar] [CrossRef]
- Wu, T.; Chen, W.; Liu, S.; Lu, H.; Wang, H.; Kong, D.; Huang, X.; Kong, Q.; Ning, Y.; Lu, Z. Huaier suppresses proliferation and induces apoptosis in human pulmonary cancer cells via upregulation of miR-26b-5p. Febs Lett. 2014, 588, 2107–2114. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Yang, Q.; Kong, X.; Haffty, B.G.; Gao, S.; Moran, M.S. Radiosensitization effect of Huaier on breast cancer cells. Oncol. Rep. 2016, 35, 2843–2850. [Google Scholar] [CrossRef]
- Yu, T.; Chen, F.; Ku, X.; Fan, J.; Zhu, Y.; Ma, H.; Li, S.; Wu, B.; He, Q. Growth characteristics and complete genomic sequence analysis of a novel pseudorabies virus in China. Virus Genes 2016, 52, 474–483. [Google Scholar] [CrossRef]
- Danthi, P. Viruses and the Diversity of Cell Death. Annu. Rev. Virol. 2016, 3, 533–553. [Google Scholar] [CrossRef]
- Zhou, X.; Jiang, W.; Liu, Z.; Liu, S.; Liang, X. Virus Infection and Death Receptor-Mediated Apoptosis. Viruses 2017, 9, 316. [Google Scholar] [CrossRef] [Green Version]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef]
- Wang, X.; Xie, F.; Zhou, X.; Chen, T.; Xue, Y.; Wang, W. 18β-Glycyrrhetinic acid inhibits the apoptosis of cells infected with rotavirus SA11 via the Fas/FasL pathway. Pharm. Biol. 2021, 59, 1098–1105. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huan, C.; Yao, J.; Xu, W.; Zhang, W.; Zhou, Z.; Pan, H.; Gao, S. Huaier Polysaccharide Interrupts PRV Infection via Reducing Virus Adsorption and Entry. Viruses 2022, 14, 745. https://doi.org/10.3390/v14040745
Huan C, Yao J, Xu W, Zhang W, Zhou Z, Pan H, Gao S. Huaier Polysaccharide Interrupts PRV Infection via Reducing Virus Adsorption and Entry. Viruses. 2022; 14(4):745. https://doi.org/10.3390/v14040745
Chicago/Turabian StyleHuan, Changchao, Jingting Yao, Weiyin Xu, Wei Zhang, Ziyan Zhou, Haochun Pan, and Song Gao. 2022. "Huaier Polysaccharide Interrupts PRV Infection via Reducing Virus Adsorption and Entry" Viruses 14, no. 4: 745. https://doi.org/10.3390/v14040745
APA StyleHuan, C., Yao, J., Xu, W., Zhang, W., Zhou, Z., Pan, H., & Gao, S. (2022). Huaier Polysaccharide Interrupts PRV Infection via Reducing Virus Adsorption and Entry. Viruses, 14(4), 745. https://doi.org/10.3390/v14040745