
HBx 128–133 Deletion Affecting HBV Mother-to-Child Transmission Weakens HBV Replication via Reducing HBx Level and CP/ENII Transcriptional Activity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Amplification, Sequencing, and Sequence Analysis
2.3. Plasmids
2.4. Antibodies and Reagents
2.5. Cell Cultures and Transfection
2.6. Detections of HBsAg and HBeAg in Supernatants
2.7. HBV DNA and HBV RNA Quantification
2.8. Western Blot
2.9. Northern Blot
2.10. Luciferase Reporter Assay
2.11. Chromatin Immunoprecipitation (ChIP)-Polymerase Chain Reaction (PCR)
2.12. Statistical Analysis
3. Results
3.1. Baseline Characteristics between the Immunoprophylaxis Failure and Success Group
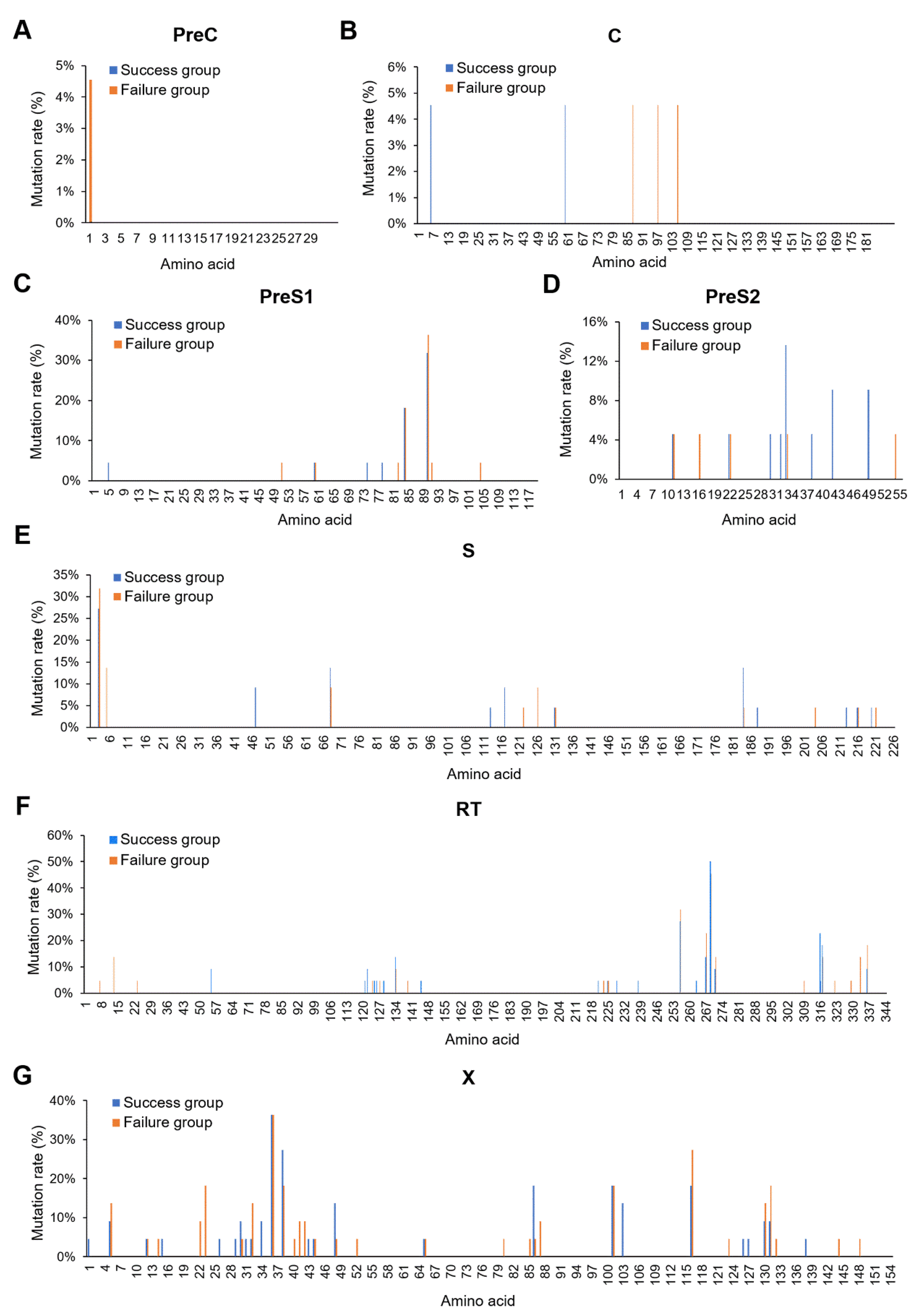
3.2. X128-133del May Affect HBV MTCT
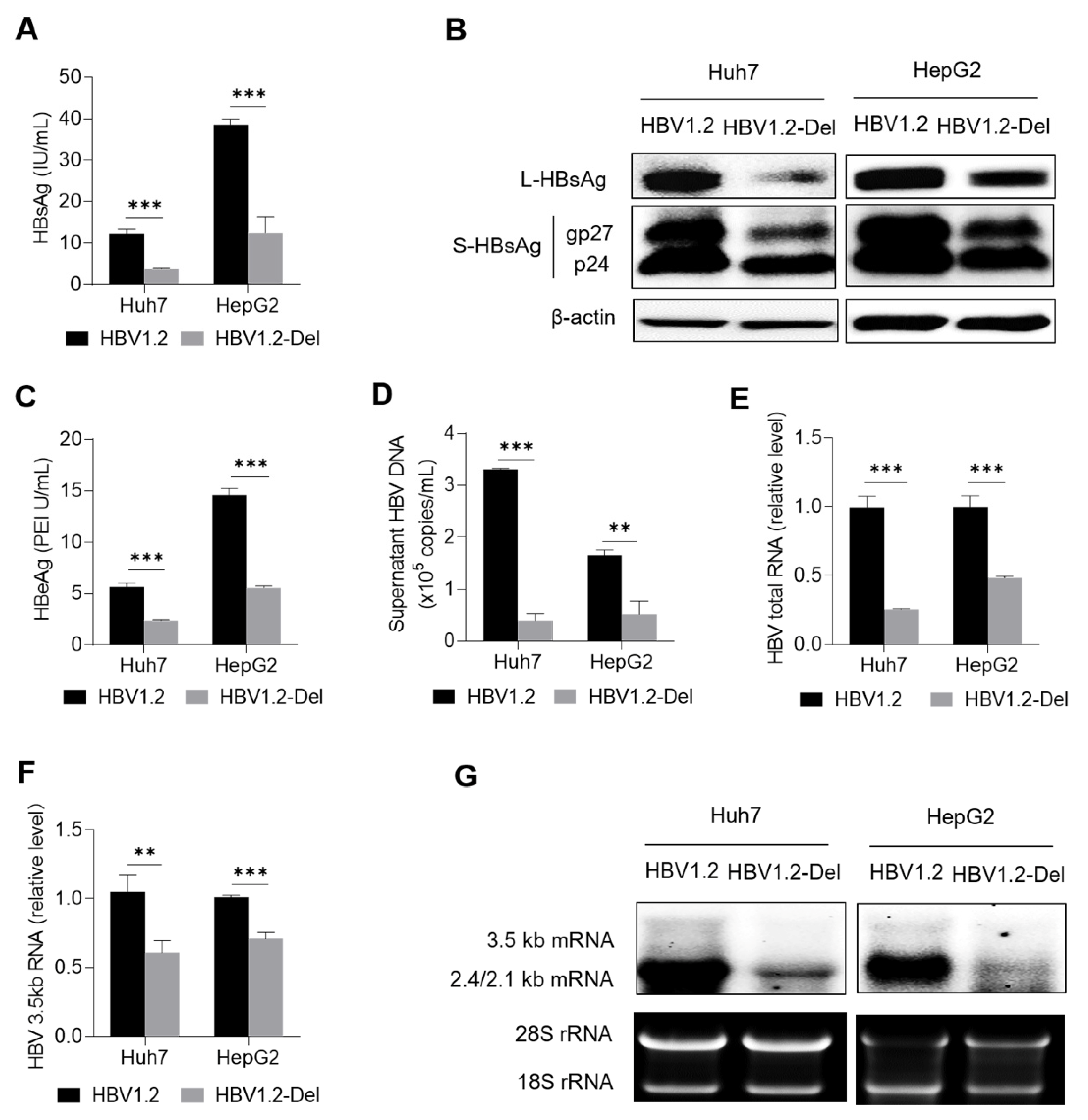
3.3. X128–133del Weakens HBV Replication
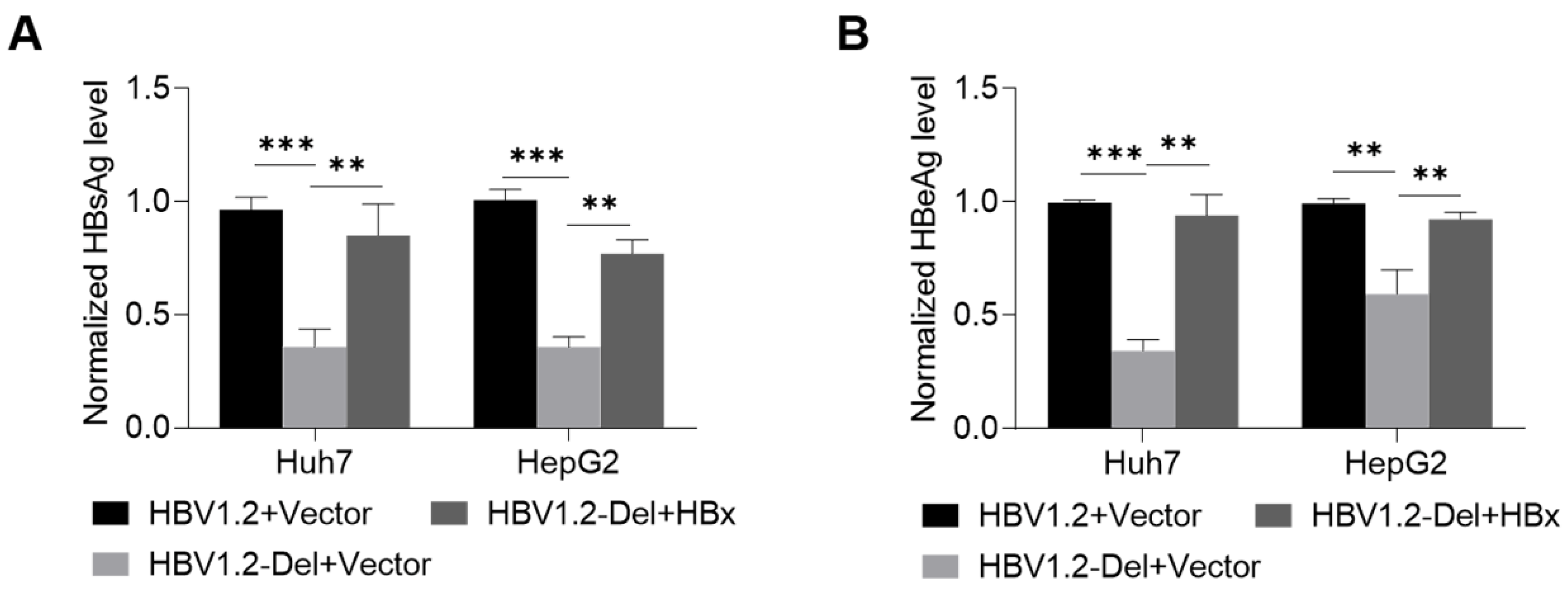
3.4. Wild-Type HBx Can Rescue the x128–133del-Weakened HBV Replication
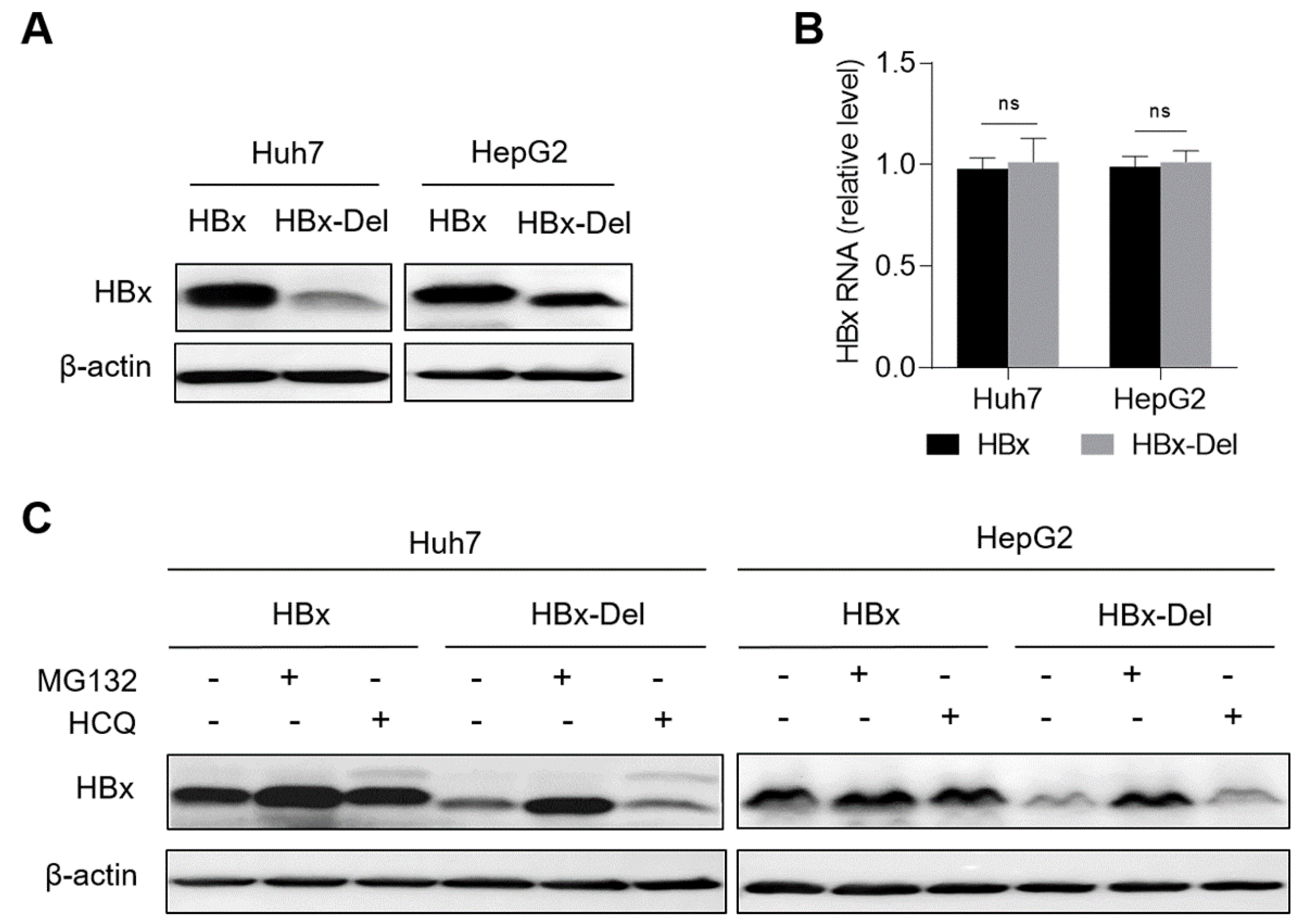
3.5. X128-133del Reduces the Level of HBx Protein through Promoting the Proteasome-Dependent Degradation
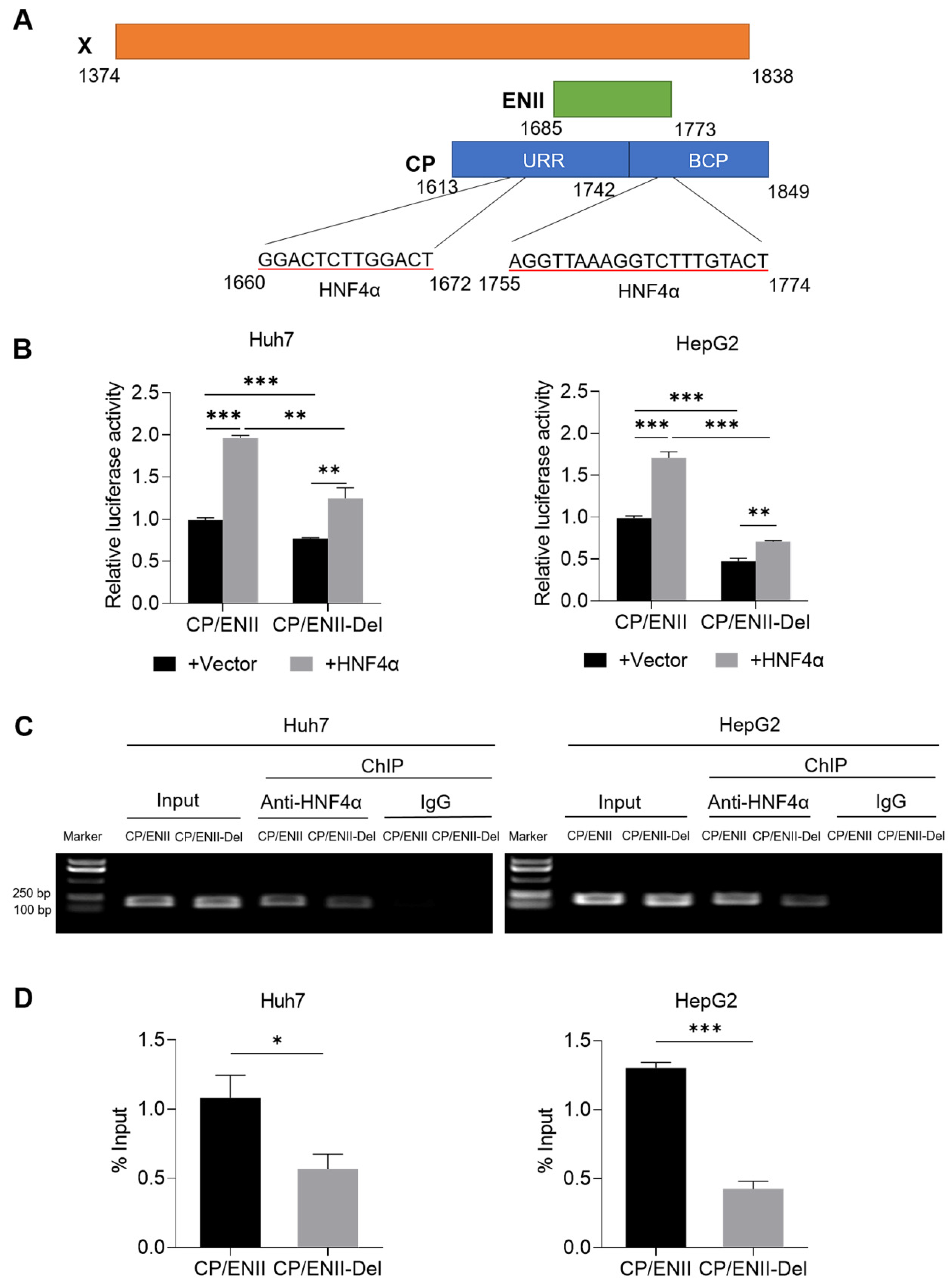
3.6. X128–133del Downregulates the Transcriptional Activity of CP/ENII through Attenuating the Binding Capacity of HNF4α to HBV CP/ENII
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hepatitis, B. World Health Organization Fact Sheets. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 27 July 2021).
- Lu, Y.; Zhu, F.C.; Liu, J.X.; Zhai, X.J.; Chang, Z.J.; Yan, L.; Wei, K.P.; Zhang, X.; Zhuang, H.; Li, J. The maternal viral threshold for antiviral prophylaxis of perinatal hepatitis B virus transmission in settings with limited resources: A large prospective cohort study in China. Vaccine 2017, 35, 6627–6633. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wang, G.; Wang, F.; Cheng, J.; Ren, H.; Zhuang, H.; Sun, J.; Li, L.; Li, J.; Meng, Q.; et al. Guideline of prevention and treatment for chronic hepatitis B (2015 Update). J. Clin. Transl. Hepatol. 2017, 5, 297–318. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Health Sector Strategy on Viral Hepatitis 2016–2021; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- SMFM; Dionne-Odom, J.; Tita, A.T.; Silverman, N.S. #38: Hepatitis B in pregnancy screening, treatment, and prevention of vertical transmission. Am. J. Obstet. Gynecol. 2016, 214, 6–14. [Google Scholar]
- Caligiuri, P.; Cerruti, R.; Icardi, G.; Bruzzone, B. Overview of hepatitis B virus mutations and their implications in the management of infection. World J. Gastroenterol. 2016, 22, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Sa-Nguanmoo, P.; Tangkijvanich, P.; Tharmaphornpilas, P.; Rasdjarmrearnsook, A.O.; Plianpanich, S.; Thawornsuk, N.; Theamboonlers, A.; Poovorawan, Y. Molecular analysis of hepatitis B virus associated with vaccine failure in infants and mothers: A case-control study in Thailand. J. Med. Virol. 2012, 84, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, B.; Peng, Z.; Tang, N.; Chen, W. Hepatitis B virus surface gene mutants in immunoprophylaxis-failed infants from Southern China. J. Med. Virol. 2019, 91, 1069–1075. [Google Scholar] [CrossRef]
- Xiao, Y.; Sun, K.; Duan, Z.; Liu, Z.; Li, Y.; Yan, L.; Song, Y.; Zou, H.; Zhuang, H.; Wang, J.; et al. Quasispecies characteristic in "a" determinant region is a potential predictor for the risk of immunoprophylaxis failure of mother-to-child-transmission of sub-genotype C2 hepatitis B virus: A prospective nested case-control study. Gut 2020, 69, 933–941. [Google Scholar] [CrossRef]
- Liu, S.L.; Dong, Y.; Zhang, L.; Li, M.W.; Wo, J.E.; Lu, L.W.; Chen, Z.J.; Wang, Y.Z.; Ruan, B. Influence of HBV gene heterogeneity on the failure of immunization with HBV vaccines in eastern China. Arch. Virol. 2009, 154, 437–443. [Google Scholar] [CrossRef]
- Wang, X.; Deng, W.; Qian, K.; Deng, H.; Huang, Y.; Tu, Z.; Huang, A.; Long, Q. Quasispecies characters of hepatitis B virus in immunoprophylaxis failure infants. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1153–1162. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, Y.; Li, L.; Song, Y.; Zhai, X.; Liu, J.; Duan, Z.; Yan, L.; Ding, F.; Liu, J.; et al. The dynamic changes of HBV quasispecies diversity in infancy after immunoprophylaxis failure: A prospective cohort study. Virol. J. 2021, 18, 236. [Google Scholar] [CrossRef]
- Wang, X.L.; Ren, J.P.; Wang, X.Q.; Wang, X.H.; Yang, S.F.; Xiong, Y. Mutations in pre-core and basic core promoter regions of hepatitis B virus in chronic hepatitis B patients. World J. Gastroenterol. 2016, 22, 3268–3274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ma, S.; Zhang, X.; Zhao, H.; Ding, H.; Zeng, C. Prevalent HBV point mutations and mutation combinations at BCP/preC region and their association with liver disease progression. BMC Infect. Dis. 2010, 10, 271. [Google Scholar] [CrossRef]
- Lu, J.F.; Li, J.E.; Liu, Y.L.; Jin, Y.; Ma, L.N.; Hu, Z.J.; Chen, X.Y. Relationship between HBV pre-C/BCP mutation/quasispecies and HBeAg and HBV DNA in HBeAg positive patients with chronic hepatitis B. Chin. J. Exp. Clin. Infect. Dis. 2020, 14, 278–283. [Google Scholar]
- Boucheron, P.; Lu, Y.; Yoshida, K.; Zhao, T.; Funk, A.L.; Lunel-Fabiani, F.; Guingané, A.; Tuaillon, E.; van Holten, J.; Chou, R.; et al. Accuracy of HBeAg to identify pregnant women at risk of transmitting hepatitis B virus to their neonates: A systematic review and meta-analysis. Lancet Infect. Dis. 2021, 21, 85–96. [Google Scholar] [CrossRef]
- Kay, A.; Zoulim, F. Hepatitis B virus genetic variability and evolution. Virus Res. 2007, 127, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Buckwold, V.E.; Hon, M.W.; Ou, J.H. Mechanism of suppression of hepatitis B virus precore RNA transcription by a frequent double mutation. J. Virol. 1999, 73, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- Candotti, D.; Danso, K.; Allain, J.P. Maternofetal transmission of hepatitis B virus genotype E in Ghana, west Africa. J. Gen. Virol. 2007, 88, 2686–2695. [Google Scholar] [CrossRef]
- Cheng, H.; Su, H.; Wang, S.; Shao, Z.; Men, K.; Li, M.; Li, S.; Zhang, J.; Xu, J.; Zhang, H.; et al. Association between genomic heterogeneity of hepatitis B virus and intrauterine infection. Virology 2009, 387, 168–175. [Google Scholar] [CrossRef]
- Wu, J.X.; Yang, Z.Q.; Zhang, R.J.; Li, Y.D.; Zhao, T.J.; Yi, L.Z.; Feng, Y.L.; Feng, S.Y.; Wang, B.; Wang, S.P. Relationship between mutations of HBV basal core promoter region in HBsAg-positive mothers and intrauterine transmission. Chin. J. Epidemiol. 2020, 41, 902–907. [Google Scholar]
- Li, Y.; Shen, C.; Yang, L.; Yang, Y.; Wang, M.; Li, S.; Chen, F.; Yang, M.; Peng, L.; Ma, J.; et al. Intra-host diversity of hepatitis B virus during mother-to-child transmission: The X gene may play a key role in virus survival in children after transmission. Arch. Virol. 2020, 165, 1279–1288. [Google Scholar] [CrossRef]
- Zhao, T.J.; Yang, Z.Q.; Li, Y.D.; Yi, L.Z.; Feng, S.Y.; Wang, B.; Feng, Y.L.; Wang, S.P. Relationship between C genotype HBV mutation and intrauterine transmission. Chin. J. Epidemiol. 2021, 42, 716–722. [Google Scholar]
- Slagle, B.L.; Bouchard, M.J. Hepatitis B virus X and regulation of viral gene expression. Cold Spring Harb. Perspect. Med. 2016, 6, a021402. [Google Scholar] [CrossRef] [PubMed]
- Lucifora, J.; Arzberger, S.; Durantel, D.; Belloni, L.; Strubin, M.; Levrero, M.; Zoulim, F.; Hantz, O.; Protzer, U. Hepatitis B virus X protein is essential to initiate and maintain virus replication after infection. J. Hepatol. 2011, 55, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.Y.; Chen, E.Q.; Huang, F.J.; Leng, X.H.; Cheng, X.; Tang, H. Role and functional domain of hepatitis B virus X protein in regulating HBV transcription and replication in vitro and in vivo. Viruses 2013, 5, 1261–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keasler, V.V.; Hodgson, A.J.; Madden, C.R.; Slagle, B.L. Enhancement of hepatitis B virus replication by the regulatory X protein in vitro and in vivo. J. Virol. 2007, 81, 2656–2662. [Google Scholar] [CrossRef]
- Tsuge, M.; Hiraga, N.; Akiyama, R.; Tanaka, S.; Matsushita, M.; Mitsui, F.; Abe, H.; Kitamura, S.; Hatakeyama, T.; Kimura, T.; et al. HBx protein is indispensable for development of viraemia in human hepatocyte chimeric mice. J. Gen. Virol. 2010, 91, 1854–1864. [Google Scholar] [CrossRef]
- Shi, S.; Liu, M.; Xi, J.; Liu, H.; Guan, G.; Shen, C.; Guo, Z.; Zhang, T.; Xu, Q.; Kudereti, D.; et al. Sex-determining region Y box 4 (SOX4) suppresses Hepatitis B virus replication by inhibiting hepatocyte nuclear factor 4α expression. Antivir. Res. 2020, 176, 104745. [Google Scholar] [CrossRef]
- Zhang, Z.; Torii, N.; Furusaka, A.; Malayaman, N.; Hu, Z.; Liang, T.J. Structural and functional characterization of interaction between hepatitis B virus X protein and the proteasome complex. J. Biol. Chem. 2000, 275, 15157–15165. [Google Scholar] [CrossRef]
- Oropeza, C.E.; Tarnow, G.; Sridhar, A.; Taha, T.Y.; Shalaby, R.E.; McLachlan, A. The regulation of HBV transcription and replication. Adv. Exp. Med. Biol. 2020, 1179, 39–69. [Google Scholar]
- Hong, X.; Kim, E.S.; Guo, H. Epigenetic regulation of hepatitis B virus covalently closed circular DNA: Implications for epigenetic therapy against chronic hepatitis B. Hepatology 2017, 66, 2066–2077. [Google Scholar] [CrossRef]
- Belloni, L.; Pollicino, T.; Nicola, F.D.; Guerrieri, F.; Raffa, G.; Fanciulli, M.; Raimondo, G.; Levrero, M. Nuclear HBx binds the HBV minichromosome and modifies the epigenetic regulation of cccDNA function. Proc. Natl. Acad. Sci. USA 2009, 106, 19975–19979. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Cheong, J.H.; Kaneko, S. Human hepatitis virus X gene encodes a regulatory domain that represses transactivation of X protein. J. Biol. Chem. 1994, 269, 15118–15123. [Google Scholar] [CrossRef]
- Liu, L.P.; Hu, B.G.; Ye, C.; Ho, R.L.; Chen, G.G.; Lai, P.B. HBx mutants differentially affect the activation of hypoxia-inducible factor-1alpha in hepatocellular carcinoma. Br. J. Cancer 2014, 110, 1066–1073. [Google Scholar] [CrossRef] [PubMed]
- Moolla, N.; Kew, M.; Arbuthnot, P. Regulatory elements of hepatitis B virus transcription. J. Viral Hepat. 2002, 9, 323–331. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunoprophylaxis Success Group | Immunoprophylaxis Failure Group | p | ||
|---|---|---|---|---|
| Mothers | ||||
| Number | 22 | 22 | ||
| Age (years), median (range) | 25.00 (20.00–35.00) | 25.50 (19.00–34.00) | 0.906 | |
| HBsAg (log10 IU/mL), median (range) | 4.51 (3.78–4.91) | 4.49 (3.49–4.88) | 0.925 | |
| HBeAg (log10 S/CO), median (range) | 3.15 (3.03–3.28) | 3.16 (2.90–3.23) | 0.907 | |
| HBV DNA (log10 IU/mL), median (range) | 8.28 (7.18–8.72) | 8.23 (7.48–8.96) | 0.823 | |
| ALT (<40 U/L), n (%) | 22 (100) | 22 (100) | – | |
| Genotype C2, n (%) | 22 (100) | 22 (100) | – | |
| Infants | ||||
| Gender, male: female | 9:13 | 13:9 | 0.228 | |
| Birth weight (kg), median (range) | 3.30 (2.60–4.25) | 3.55 (2.60–4.00) | 0.204 | |
| Parturition manner, cesarean: vaginal | 11:11 | 12:10 | 0.763 | |
| Feeding pattern, breast a: artificial | 8:14 | 7:15 | 0.750 | |
| Infant’s age at first dose of HepB (hours), n (%) | <12 h | 22 (100) | 19 (86.36) | 0.232 |
| 12–24 h | 0 | 3 (13.64) | ||
| Nucleotide Mutation | Region | Amino Acid Mutation | Success Group, n (%) | Failure Group, n (%) | p | Qa |
|---|---|---|---|---|---|---|
| G1437A | X | xG22S | 0 | 2 (9.09%) | 0.023 | 0.052 |
| G1440A | X | xA23T | 4 (3.33%) | 4 (18.18%) | 0.023 | 0.052 |
| T1464C | X | xS31P | 7 (5.83%) | 0 | 0.531 | 0.534 |
| G1467C | X | xG32R | 3 (2.50%) | 3 (13.64%) | 0.048 | 0.086 |
| C1498A/A1499C | X | xS42Y | 0 | 2 (9.09%) | 0.023 | 0.052 |
| G1515A | X | xD48N | 15 (12.50%) | 1 (4.55%) | 0.473 | 0.534 |
| A1630G | X/CP | xH86R | 14 (11.67%) | 1 (4.55%) | 0.534 | 0.534 |
| A1680C | X/CP | xM103L | 7 (5.83%) | 0 | 0.531 | 0.534 |
| 1755–1772del | X/CP/ENII | x128–133del | 33 (27.50%) | 0 | 0.005 | 0.045 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Lu, Y.; Li, Y.; Liu, M.; Zhuang, H.; Li, J.; Wang, J. HBx 128–133 Deletion Affecting HBV Mother-to-Child Transmission Weakens HBV Replication via Reducing HBx Level and CP/ENII Transcriptional Activity. Viruses 2022, 14, 1887. https://doi.org/10.3390/v14091887
Song Y, Lu Y, Li Y, Liu M, Zhuang H, Li J, Wang J. HBx 128–133 Deletion Affecting HBV Mother-to-Child Transmission Weakens HBV Replication via Reducing HBx Level and CP/ENII Transcriptional Activity. Viruses. 2022; 14(9):1887. https://doi.org/10.3390/v14091887
Chicago/Turabian StyleSong, Yarong, Ying Lu, Yi Li, Minmin Liu, Hui Zhuang, Jie Li, and Jie Wang. 2022. "HBx 128–133 Deletion Affecting HBV Mother-to-Child Transmission Weakens HBV Replication via Reducing HBx Level and CP/ENII Transcriptional Activity" Viruses 14, no. 9: 1887. https://doi.org/10.3390/v14091887
APA StyleSong, Y., Lu, Y., Li, Y., Liu, M., Zhuang, H., Li, J., & Wang, J. (2022). HBx 128–133 Deletion Affecting HBV Mother-to-Child Transmission Weakens HBV Replication via Reducing HBx Level and CP/ENII Transcriptional Activity. Viruses, 14(9), 1887. https://doi.org/10.3390/v14091887