
Development and Characterization of a Genetically Stable Infectious Clone for a Genotype I Isolate of Dengue Virus Serotype 1



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and DENV-Infected Serum
2.2. Determination of Viral Genome Sequence and Construction of Full-Length cDNA Clone
2.3. Transformation and Purification of Plasmids
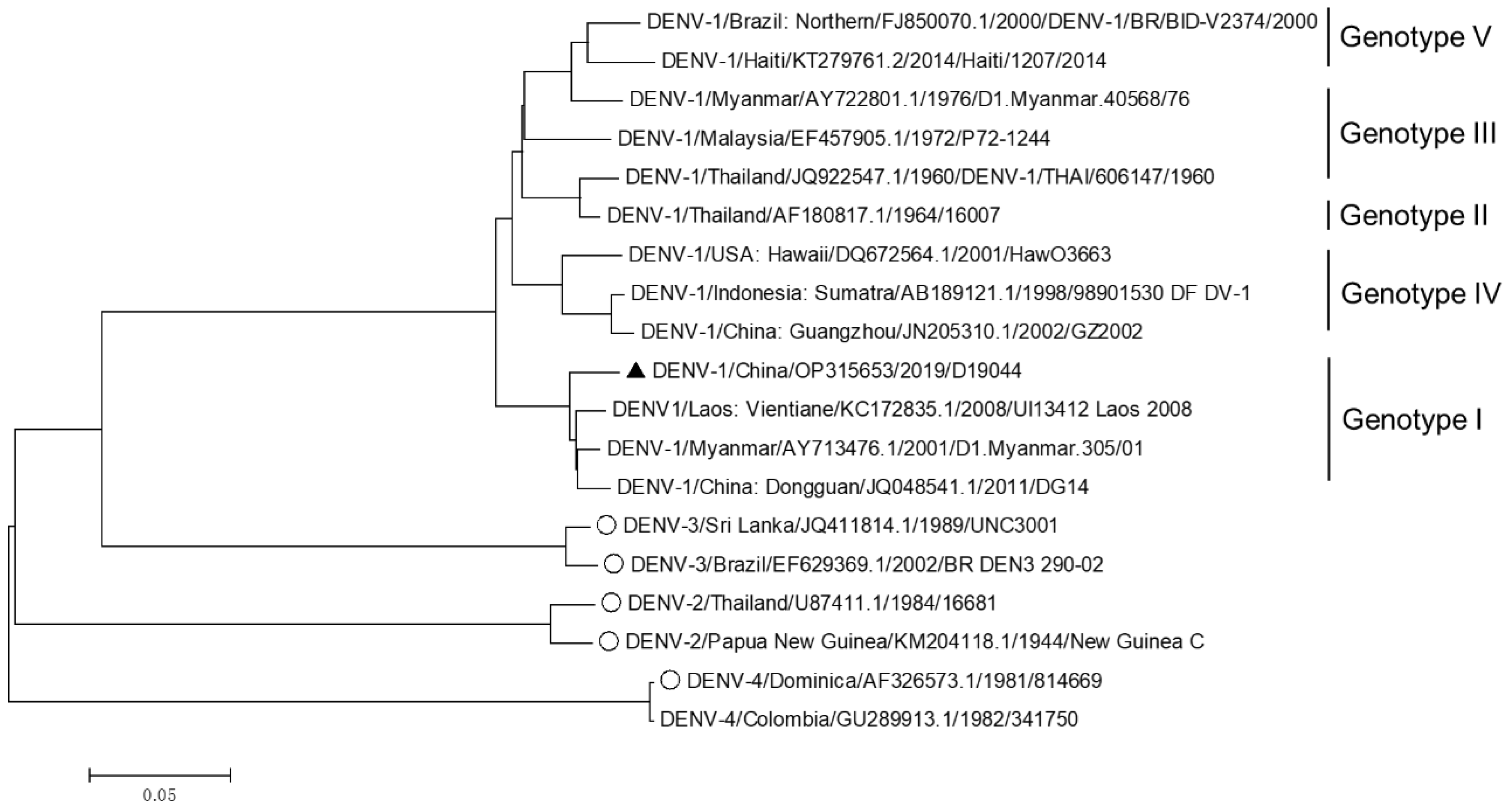
2.4. Phylogenetic Analysis of D19044 and Other DENV Sequences
2.5. Virus Stock Preparation, Infection Experiment, and Virus Passage
2.6. Virus Titration and Indirect Immunofluorescence Foci Assay (IFA)
2.7. Cytotoxicity Assay
2.8. Antiviral Drug Treatment
3. Results
3.1. Genome Sequence of DENV-1 Genotype I Isolate, D19044
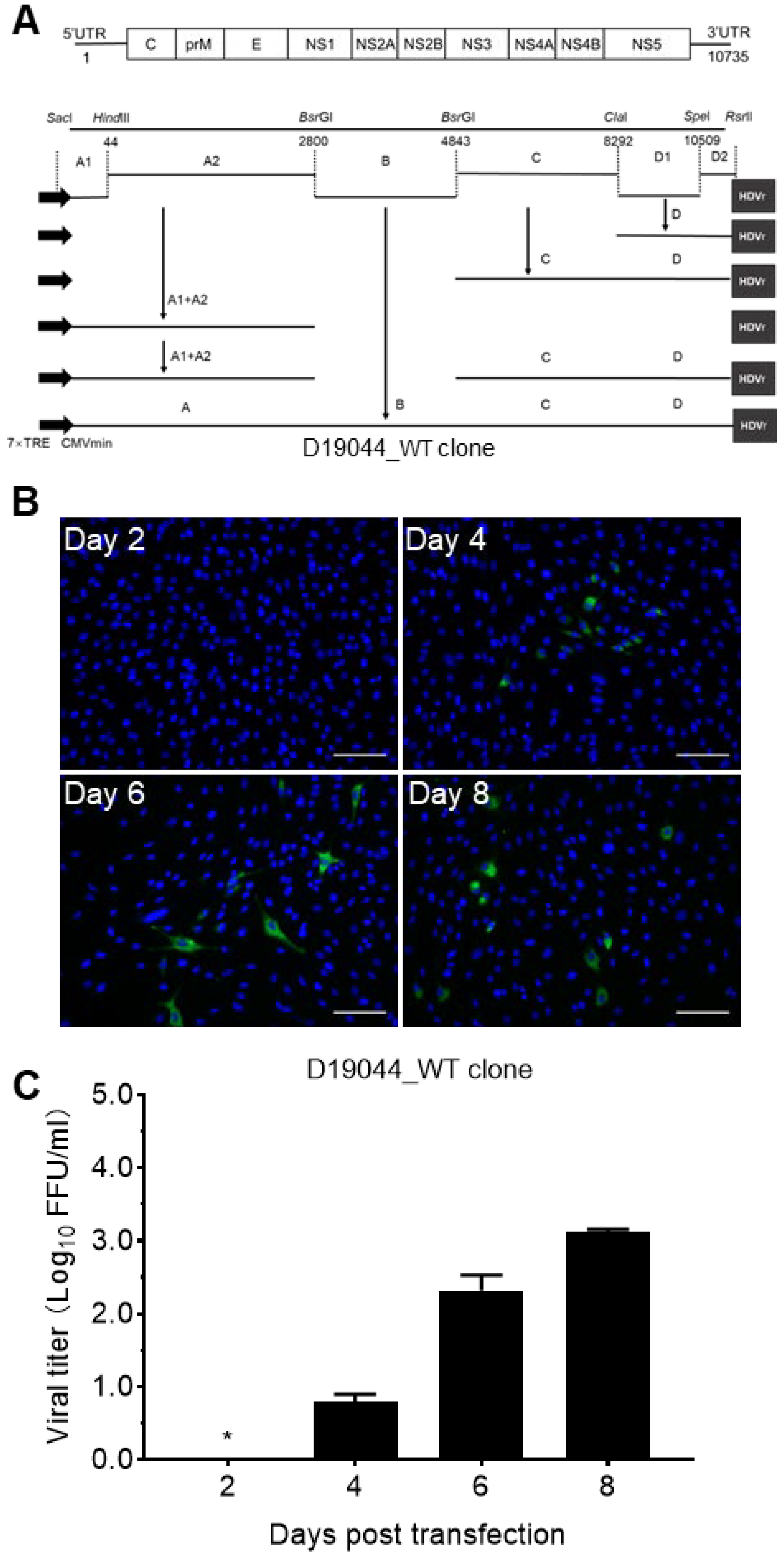
3.2. Construction of Full-Length D19044 cDNA Clone
3.3. Transfection of D19044 Clone Plasmid Produced Infectious DENV Particles
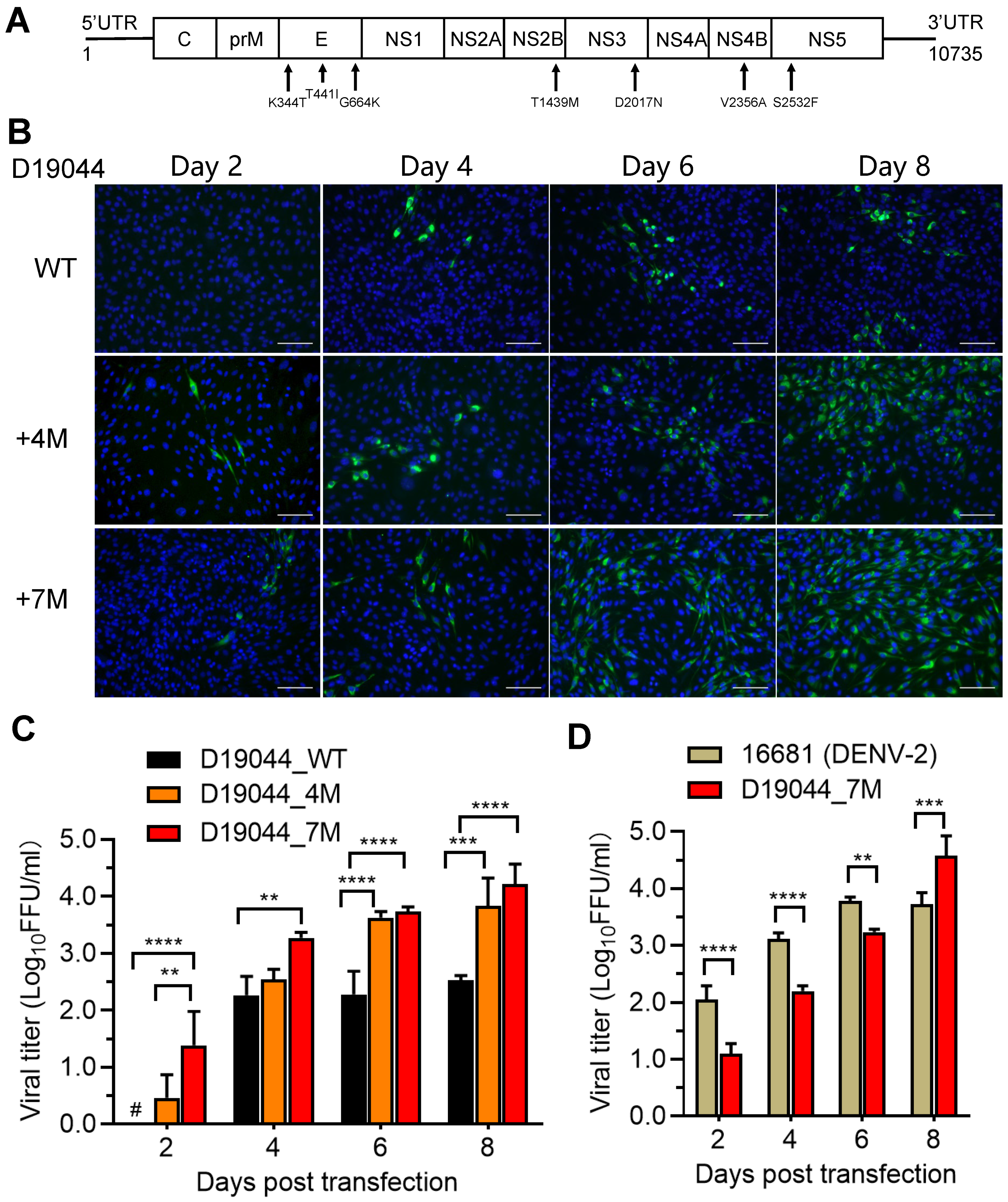
3.4. Efficient Virus Production of D19044 Clone Carrying Amino Acid Substitutions
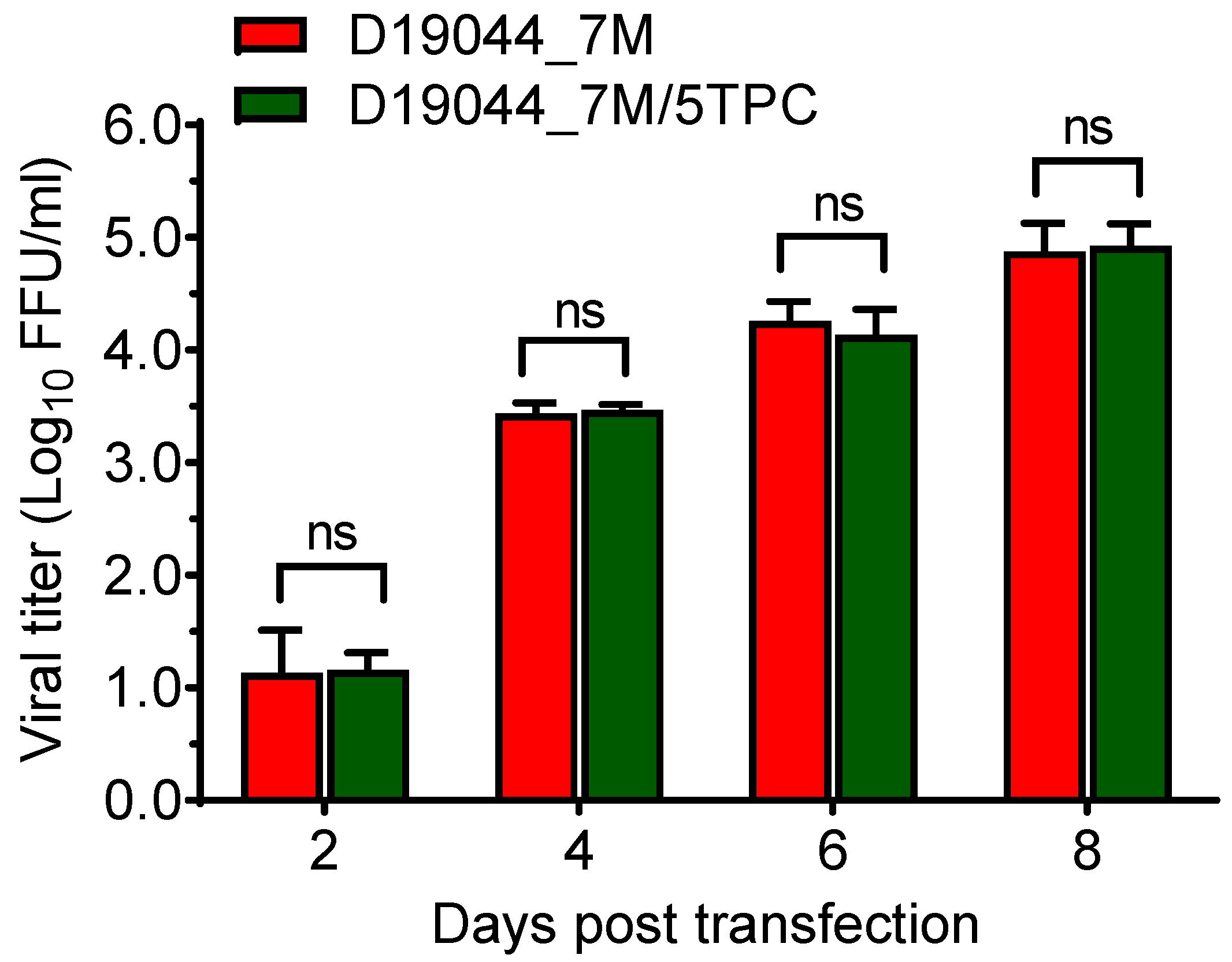
3.5. D19044_7M Clone Was Genetically Stable in Bacteria after Multiple Transformation-Purification Cycles
3.6. Differential Efficiency of D19044_7M and D19044_WT in Various Vell Lines
3.7. D19044_7M Virus Was Inhibited by Antiviral Compound
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, T.; Wu, Z.; Li, Y.P. Dengue fever and dengue virus in the People’s Republic of China. Rev. Med. Virol. 2022, 32, e2245. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Zachariah, M.; Harris, E. Mycophenolic Acid Inhibits Dengue Virus Infection by Preventing Replication of Viral RNA. Virology 2002, 304, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Zhou, Z.; Wen, Z.; Liu, Y.; Zeng, C.; Xiao, D.; Ou, M.; Han, Y.; Huang, S.; Liu, D.; et al. Global Epidemiology of Dengue Outbreaks in 1990–2015: A Systematic Review and Meta-Analysis. Front. Cell. Infect. Microbiol. 2017, 7, 317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Rong, L.; Li, Y.-P. Flaviviridae Viruses and Oxidative Stress: Implications for Viral Pathogenesis. Oxidative Med. Cell. Longev. 2019, 2019, 1409582. [Google Scholar] [CrossRef]
- Wilder-Smith, A. Dengue vaccine development: Status and future. Bundesgesundheitsblatt Gesundh. Gesundh. 2020, 63, 40–44. [Google Scholar] [CrossRef]
- WHO. Ten Threats to Global Health in 2019. WHO Spotlight 2019. Available online: https://www.who.int/news-room/spotlight/ten-threats-to-global-health-in-2019 (accessed on 29 May 2022).
- Holmes, E.C.; Twiddy, S.S. The origin, emergence and evolutionary genetics of dengue virus. Infect. Genet. Evol. 2003, 3, 19–28. [Google Scholar] [CrossRef]
- Rico-Hesse, R. Molecular evolution and distribution of dengue viruses type 1 and 2 in nature. Virology 1990, 174, 479–493. [Google Scholar] [CrossRef]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- Khetarpal, N.; Khanna, I. Dengue Fever: Causes, Complications, and Vaccine Strategies. J. Immunol. Res. 2016, 2016, 6803098. [Google Scholar] [CrossRef]
- Langerak, T.; Mumtaz, N.; Tolk, V.I.; Van Gorp, E.C.M.; Martina, B.E.; Rockx, B.; Koopmans, M.P.G. The possible role of cross-reactive dengue virus antibodies in Zika virus pathogenesis. PLOS Pathog. 2019, 15, e1007640. [Google Scholar] [CrossRef]
- Mishra, N.; Boudewijns, R.; Schmid, M.A.; Marques, R.E.; Sharma, S.; Neyts, J.; Dallmeier, K. A Chimeric Japanese Encephalitis Vaccine Protects against Lethal Yellow Fever Virus Infection without Inducing Neutralizing Antibodies. mBio 2020, 11, e02494-19. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Ramasamy, V.; Shanmugam, R.K.; Ahuja, R.; Khanna, N. Antibody-Dependent Enhancement: A Challenge for Developing a Safe Dengue Vaccine. Front. Cell. Infect. Microbiol. 2020, 10, 572681. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B.; Dans, L.F. Dengue infection and advances in dengue vaccines for children. Lancet Child Adolesc. Health 2019, 3, 734–741. [Google Scholar] [CrossRef]
- Thai, K.T.D.; Henn, M.R.; Zody, M.C.; Tricou, V.; Nguyet, N.M.; Charlebois, P.; Lennon, N.J.; Green, L.; de Vries, P.J.; Hien, T.T.; et al. High-Resolution Analysis of Intrahost Genetic Diversity in Dengue Virus Serotype 1 Infection Identifies Mixed Infections. J. Virol. 2012, 86, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Pyke, A.T.; Moore, P.R.; Taylor, C.T.; Hall-Mendelin, S.; Cameron, J.N.; Hewitson, G.R.; Pukallus, D.S.; Huang, B.; Warrilow, D.; van den Hurk, A.F. Highly divergent dengue virus type 1 genotype sets a new distance record. Sci. Rep. 2016, 6, 22356. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Zhang, J.; Madan, V.; Biswas, A.; Schwoerer, M.P.; Cafiero, T.R.; Heller, B.L.; Wang, W.; Ploss, A. Generation and characterization of genetically and antigenically diverse infectious clones of dengue virus serotypes 1–4. Emerg. Microbes Infect. 2022, 11, 227–239. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, T.; Zhang, Z.; Chen, M.; Rong, L.; Ma, L.; Yu, B.; Wu, D.; Zhang, P.; Zhu, X.; et al. Novel genetically stable infectious clone for a Zika virus clinical isolate and identification of RNA elements essential for virus production. Virus Res. 2018, 257, 14–24. [Google Scholar] [CrossRef]
- Pu, S.-Y.; Wu, R.-H.; Tsai, M.-H.; Yang, C.-C.; Chang, C.-M.; Yueh, A. A novel approach to propagate flavivirus infectious cDNA clones in bacteria by introducing tandem repeat sequences upstream of virus genome. J. Gen. Virol. 2014, 95, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Pu, S.-Y.; Wu, R.-H.; Yang, C.-C.; Jao, T.-M.; Tsai, M.-H.; Wang, J.-C.; Lin, H.-M.; Chao, Y.-S.; Yueh, A. Successful Propagation of Flavivirus Infectious cDNAs by a Novel Method to Reduce the Cryptic Bacterial Promoter Activity of Virus Genomes. J. Virol. 2011, 85, 2927–2941. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Zhang, J.; Zhu, J.; Hu, Z.; Liu, J.; Yang, J.; Yuan, W.; Shang, W.; Rao, X. Genomic phylogenetic analysis of one Dengue virus type 1 strain GZ2002 isolated in Guangzhou. China J. Mod. Med. 2012, 22, 25–33. [Google Scholar]
- de Borba, L.; Villordo, S.M.; Iglesias, N.G.; Filomatori, C.V.; Gebhard, L.G.; Gamarnik, A.V. Overlapping Local and Long-Range RNA-RNA Interactions Modulate Dengue Virus Genome Cyclization and Replication. J. Virol. 2015, 89, 3430–3437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, W.C.; Soto-Acosta, R.; Bradrick, S.S.; Garcia-Blanco, M.A.; Ooi, E.E. The 5’ and 3’ Untranslated Regions of the Flaviviral Genome. Viruses 2017, 9, 137. [Google Scholar] [CrossRef]
- Payne, A.F.; Binduga-Gajewska, I.; Kauffman, E.B.; Kramer, L.D. Quantitation of flaviviruses by fluorescent focus assay. J. Virol. Methods 2006, 134, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Schoepp, R.J.; Beaty, B.J. Titration of dengue viruses by immunofluorescence in microtiter plates. J. Clin. Microbiol. 1984, 20, 1017–1019. [Google Scholar] [CrossRef]
- Sriburi, R.; Keelapang, P.; Duangchinda, T.; Pruksakorn, S.; Maneekarn, N.; Malasit, P.; Sittisombut, N. Construction of infectious dengue 2 virus cDNA clones using high copy number plasmid. J. Virol. Methods 2001, 92, 71–82. [Google Scholar] [CrossRef]
- Kinney, R.M.; Butrapet, S.; Chang, G.-J.J.; Tsuchiya, K.R.; Roehrig, J.; Bhamarapravati, N.; Gubler, D.J. Construction of Infectious cDNA Clones for Dengue 2 Virus: Strain 16681 and Its Attenuated Vaccine Derivative, Strain PDK-53. Virology 1997, 230, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Jin, H.; Nie, X.; Zhao, Y.; Feng, N.; Cao, Z.; Tan, S.; Zhang, B.; Gai, W.; Yan, F.; et al. Development of a reverse genetics system for Japanese encephalitis virus strain SA14-14-2. Virus Genes 2019, 55, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Mishin, V.P.; Cominelli, F.; Yamshchikov, V.F. A ‘minima’ approach in design of flavivirus infectious DNA. Virus Res. 2001, 81, 113–123. [Google Scholar] [CrossRef]
- Schwarz, M.C.; Sourisseau, M.; Espino, M.M.; Gray, E.S.; Chambers, M.T.; Tortorella, D.; Evans, M.J. Rescue of the 1947 Zika Virus Prototype Strain with a Cytomegalovirus Promoter-Driven cDNA Clone. mSphere 2016, 1, e00246-16. [Google Scholar] [CrossRef] [PubMed]
- Barrows, N.J.; Campos, R.K.; Powell, S.T.; Prasanth, K.R.; Schott-Lerner, G.; Soto-Acosta, R.; Galarza-Muñoz, G.; McGrath, E.L.; Urrabaz-Garza, R.; Gao, J.; et al. A Screen of FDA-Approved Drugs for Inhibitors of Zika Virus Infection. Cell Host Microbe 2016, 20, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Kang, S.; Dimopoulos, G. Identification of anti-flaviviral drugs with mosquitocidal and anti-Zika virus activity in Aedes aegypti. PLoS Negl. Trop. Dis. 2019, 13, e0007681. [Google Scholar] [CrossRef] [Green Version]
- Shandil, R.K.; Jayaram, R.; Kaur, P.; Gaonkar, S.; Suresh, B.L.; Mahesh, B.N.; Jayashree, R.; Nandi, V.; Bharath, S.; Balasubramanian, V. Moxifloxacin, ofloxacin, sparfloxacin, and ciprofloxacin against Mycobacterium tuberculosis: Evaluation of in vitro and pharmacodynamic indices that best predict in vivo efficacy. Antimicrob. Agents Chemother. 2007, 51, 576–582. [Google Scholar] [CrossRef]
- Gallichotte, E.N.; Menachery, V.D.; Yount, B.L., Jr.; Widman, D.G.; Dinnon, K.H., III; Hartman, S.; de Silva, A.M.; Baric, R.S. Epitope Addition and Ablation via Manipulation of a Dengue Virus Serotype 1 Infectious Clone. mSphere 2017, 2, e00380-16. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.D. Development and Application of Dengue Virus Reverse Genetic Systems. Methods Mol. Biol. 2014, 1138, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.J.; Zhao, B.T.; Hori, H.; Bray, M. Infectious RNA transcribed from stably cloned full-length cDNA of dengue type 4 virus. Proc. Natl. Acad. Sci. USA 1991, 88, 5139–5143. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; de Borba, L.; Duarte dos Santos, C.N.; Mason, P.W. Construction of an infectious cDNA clone for a Brazilian prototype strain of dengue virus type 1: Characterization of a temperature-sensitive mutation in NS1. Virology 2007, 362, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Usme-Ciro, J.A.; Lopera, J.A.; Enjuanes, L.; Almazán, F.; Gallego-Gomez, J.C. Development of a novel DNA-launched dengue virus type 2 infectious clone assembled in a bacterial artificial chromosome. Virus Res. 2014, 180, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Polo, S.; Ketner, G.; Levis, R.; Falgout, B. Infectious RNA transcripts from full-length dengue virus type 2 cDNA clones made in yeast. J. Virol. 1997, 71, 5366–5374. [Google Scholar] [CrossRef]
- Kelly, E.P.; Puri, B.; Sun, W.; Falgout, B. Identification of mutations in a candidate dengue 4 vaccine strain 341750 PDK20 and construction of a full-length cDNA clone of the PDK20 vaccine candidate. Vaccine 2010, 28, 3030–3037. [Google Scholar] [CrossRef]
- Puri, B.; Polo, S.; Hayes, C.G.; Falgout, B. Construction of a full length infectious clone for dengue-1 virus Western Pacific, 74 strain. Virus Genes 2000, 20, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.; Cordeiro, M.T.; Bertani, G.R.; Marques, E.; Gil, L.H.V.G. Construction and characterisation of a complete reverse genetics system of dengue virus type 3. Mem. Inst. Oswaldo Cruz 2013, 108, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Tian, J. Circular Polymerase Extension Cloning of Complex Gene Libraries and Pathways. PLoS ONE 2009, 4, e6441. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; He, Y.; Wang, X.; Jiang, B.; Lin, X.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Zhao, X.; et al. Stabilization of a full-length infectious cDNA clone for duck Tembusu virus by insertion of an intron. J. Virol. Methods 2020, 283, 113922. [Google Scholar] [CrossRef]
- Govindarajan, D.; Guan, L.; Meschino, S.; Fridman, A.; Bagchi, A.; Pak, I.; ter Meulen, J.; Casimiro, D.R.; Bett, A.J. A Rapid and Improved Method to Generate Recombinant Dengue Virus Vaccine Candidates. PLoS ONE 2016, 11, e0152209. [Google Scholar] [CrossRef] [PubMed]
- Aubry, F.; Nougairède, A.; De Fabritus, L.; Querat, G.; Gould, E.A.; De Lamballerie, X. Single-stranded positive-sense RNA viruses generated in days using infectious subgenomic amplicons. J. Gen. Virol. 2014, 95, 2462–2467. [Google Scholar] [CrossRef] [PubMed]
- Driouich, J.-S.; Ali, S.M.; Amroun, A.; Aubry, F.; De Lamballerie, X.; Nougairède, A. SuPReMe: A rapid reverse genetics method to generate clonal populations of recombinant RNA viruses. Emerg. Microbes Infect. 2018, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zheng, F.; Yuan, G.; Duan, X.; Rong, L.; Liu, J.; Feng, S.; Wang, Z.; Wang, M.; Feng, Y.; et al. Development of an Infectious Cell Culture System for Hepatitis C Virus Genotype 6a Clinical Isolate Using a Novel Strategy and Its Sensitivity to Direct-Acting Antivirals. Front. Microbiol. 2018, 9, 2950. [Google Scholar] [CrossRef]
- Li, J.; Zhou, Q.; Rong, L.; Rong, D.; Yang, Y.; Hao, J.; Zhang, Z.; Ma, L.; Rao, G.; Zhou, Y.; et al. Development of cell culture infectious clones for hepatitis C virus genotype 1b and transcription analysis of 1b-infected hepatoma cells. Antivir. Res. 2021, 193, 105136. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Position | Sequence (5′-3′) | Product Length (nt) |
|---|---|---|---|
| F17 | 17–37 | GGACCGACAAGAACAGTTTCG | 3331 |
| R3347 | 3324–3347 | CATGAATTATCTTCCCTGTGACTG | |
| F2724 | 2724–2743 | AAATGATTAGGCCACAACCC | 2174 |
| R4897 | 4877–4897 | AATGGCTCCAACTTCACCTTC | |
| F4755 | 4755–4778 | ATGGAGGAGGTTGGAGGCTTCAAG | 3593 |
| R8347 | 8324–8347 | GCCTAGGTCCACGTCTCTTTCATA | |
| F7485 | 7485–7510 | TTTCCATGGCAAACATTTTCAGAGGA | 3060 |
| R10544 | 10,524–10,544 | GTTGTGTCTTGGGAGGGGTCT | |
| F-5′UTR | Vector-15 | gagctcgtttagtgaaccgtcAGTTGTTAGTCTACG | 70 |
| R-5′UTR | 15–49 | AAGCTTCCGATTTGAAACTGTTCTTGTCGGTCCAC | |
| F-3′UTR | 10,509–10,529 | ACTAGTGGTTAGAGGAGACCC | 277 |
| R-3′UTR | Vector | ttcggatgcccaggtcggaccgcgaggaggtggagat | |
| R6260 * | 6239–6260 | CCACGTCCATGTTCTCCTCCAA | |
| R10654 * | 10,633–10,654 | GGTCTCTCCCAGCGTCAATATG |
| Number of Passage | Nucleotide Position | Nucleotide Change * | Gene Region | Amino Acid Position | Amino Acid Change |
|---|---|---|---|---|---|
| 21 | 2084 | G–A | E | 664 | Glu–Lys |
| 21 | 4410 | C–T | NS2B | 1439 | Thr–Met |
| 21 | 6143 | G–A | NS3 | 2017 | Asp–Asn |
| 36 | 7161 | T–C/t | NS4B | 2356 | Val–Ala |
| 36 | 7689 | C–C/t | NS5 | 2532 | Ser–Phe |
| 37 | 1125 | A–C | E | 344 | Lys–Thr |
| 37 | 1416 | C–T/c | E | 441 | Thr–Ile |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, M.; Wu, T.; Yang, Y.; Chen, T.; Hao, J.; Wei, Y.; Luo, T.; Wu, D.; Li, Y.-P. Development and Characterization of a Genetically Stable Infectious Clone for a Genotype I Isolate of Dengue Virus Serotype 1. Viruses 2022, 14, 2073. https://doi.org/10.3390/v14092073
Hu M, Wu T, Yang Y, Chen T, Hao J, Wei Y, Luo T, Wu D, Li Y-P. Development and Characterization of a Genetically Stable Infectious Clone for a Genotype I Isolate of Dengue Virus Serotype 1. Viruses. 2022; 14(9):2073. https://doi.org/10.3390/v14092073
Chicago/Turabian StyleHu, Mingyue, Tiantian Wu, Yang Yang, Tongling Chen, Jiawei Hao, Youchuan Wei, Tingrong Luo, De Wu, and Yi-Ping Li. 2022. "Development and Characterization of a Genetically Stable Infectious Clone for a Genotype I Isolate of Dengue Virus Serotype 1" Viruses 14, no. 9: 2073. https://doi.org/10.3390/v14092073
APA StyleHu, M., Wu, T., Yang, Y., Chen, T., Hao, J., Wei, Y., Luo, T., Wu, D., & Li, Y.-P. (2022). Development and Characterization of a Genetically Stable Infectious Clone for a Genotype I Isolate of Dengue Virus Serotype 1. Viruses, 14(9), 2073. https://doi.org/10.3390/v14092073