
Maternal Th17 Profile after Zika Virus Infection Is Involved in Congenital Zika Syndrome Development in Children

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. PRNT (Plaque Reduction Neutralization Test)
2.3. Immunophenotyping Analysis of Lymphocyte Populations
2.4. Cytokine Evaluation by Cytometric Bead Array (CBA)
2.5. Data Analysis
2.6. Ethical Aspects
3. Results
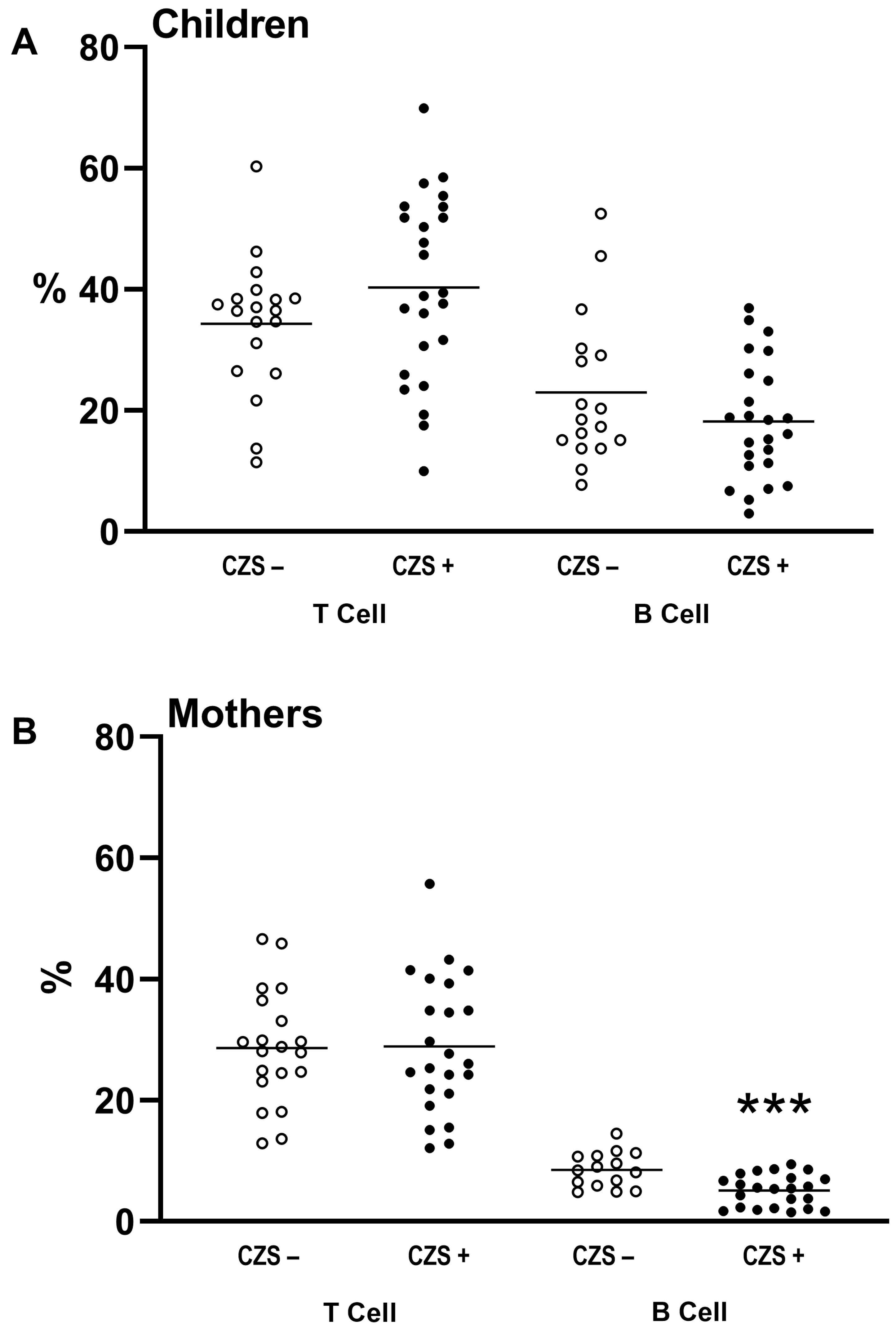
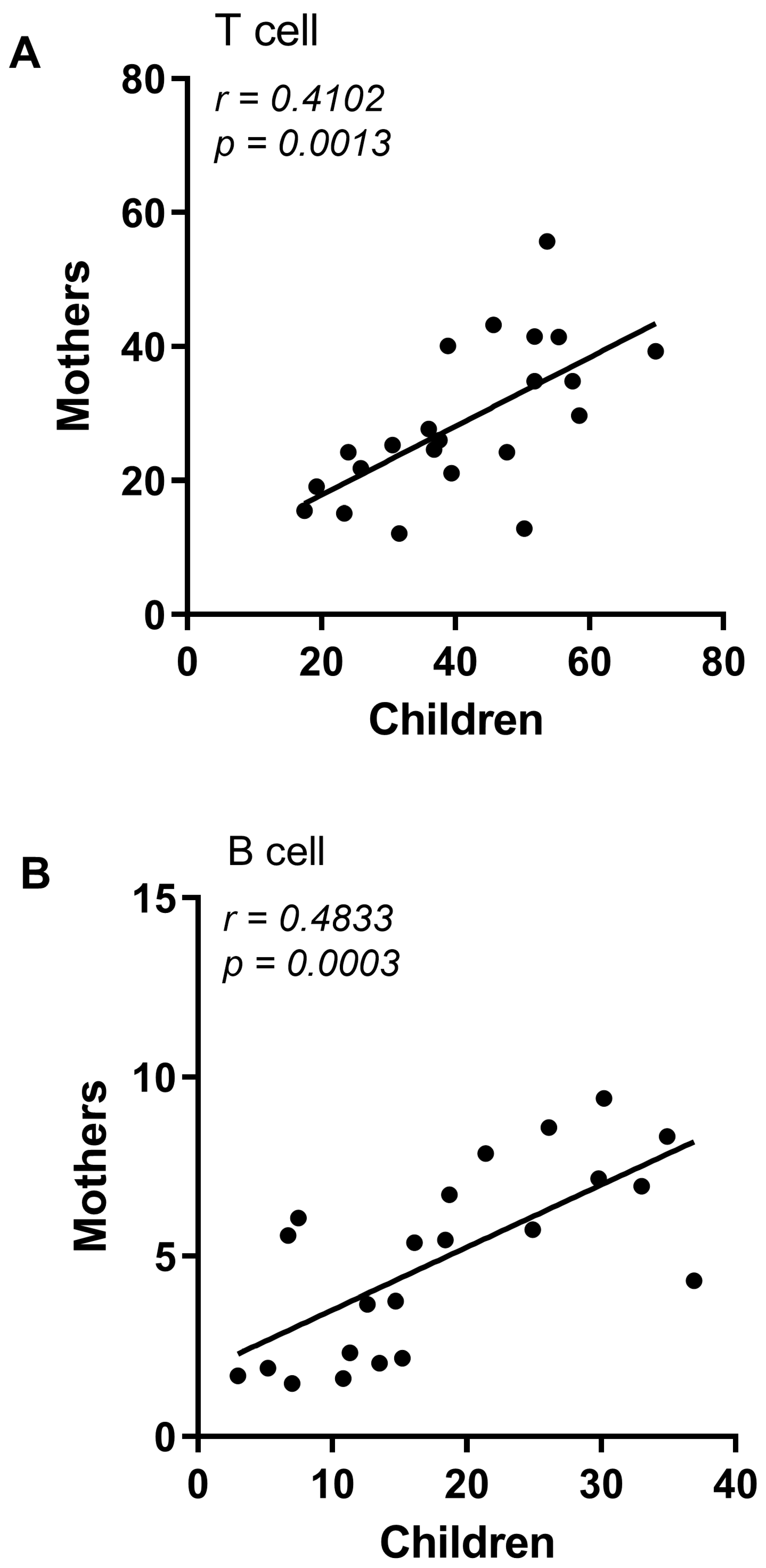
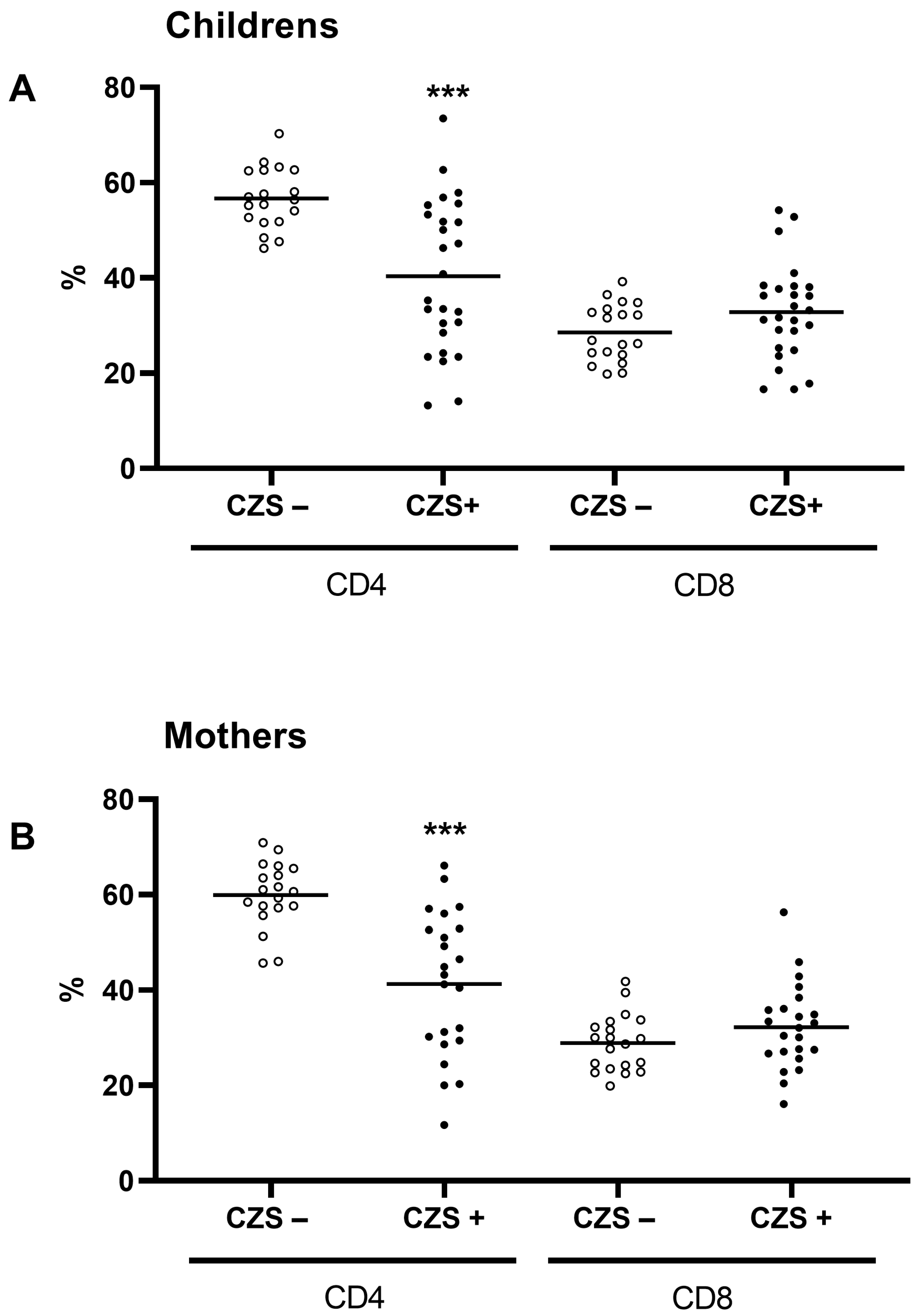
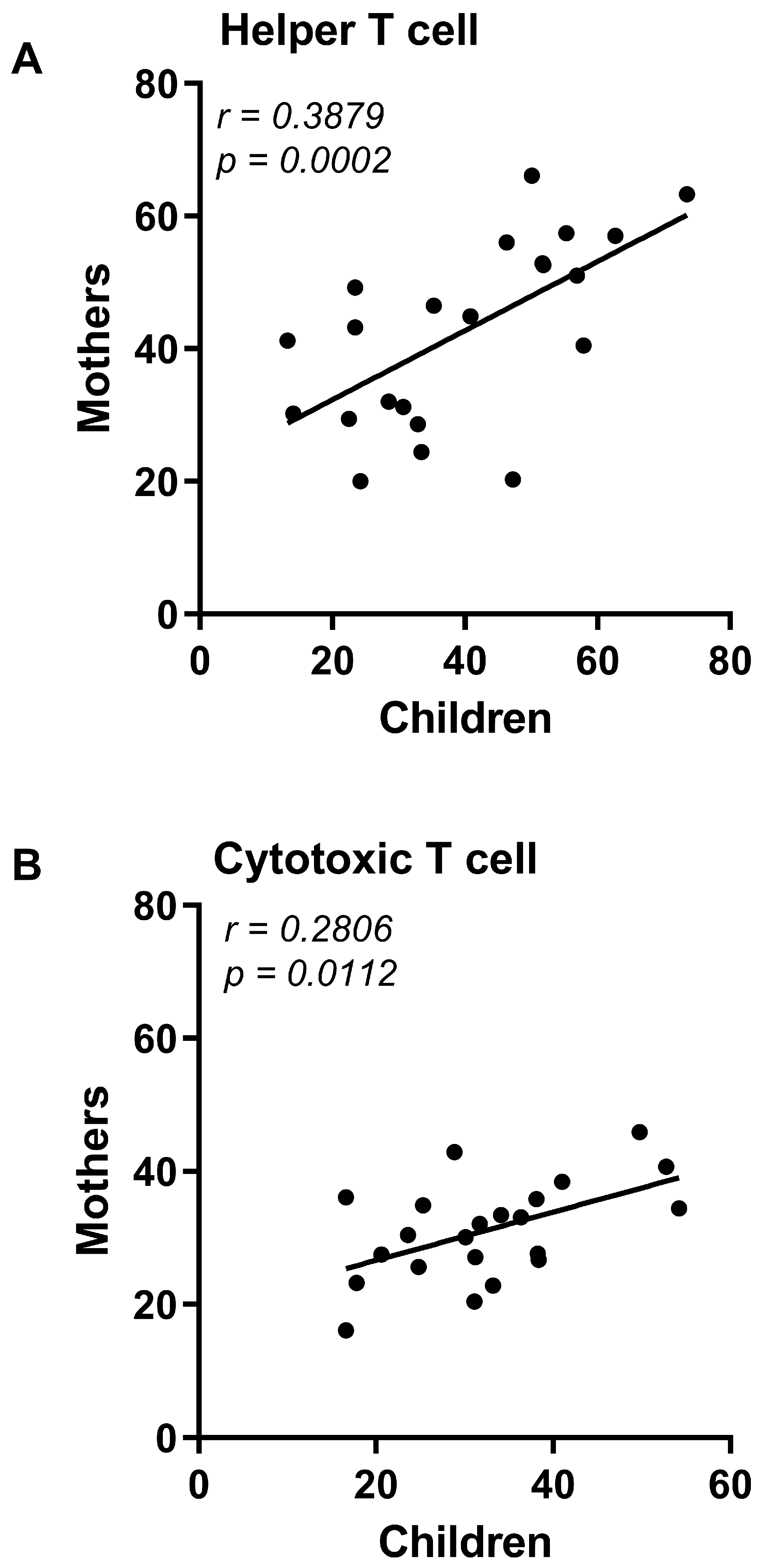
3.1. CZS+ Children and Their Mothers Present Similar T and B Lymphocyte Profile
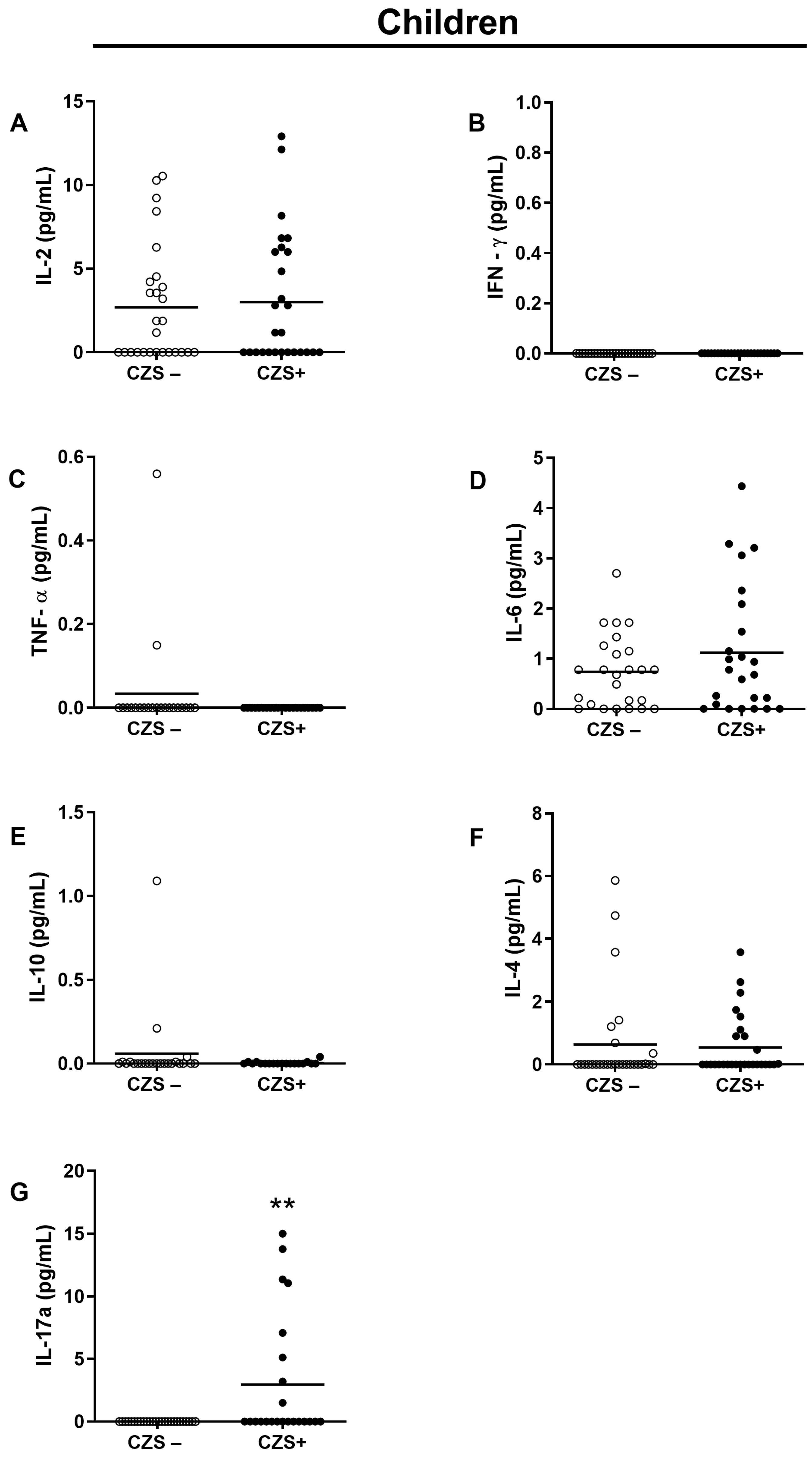
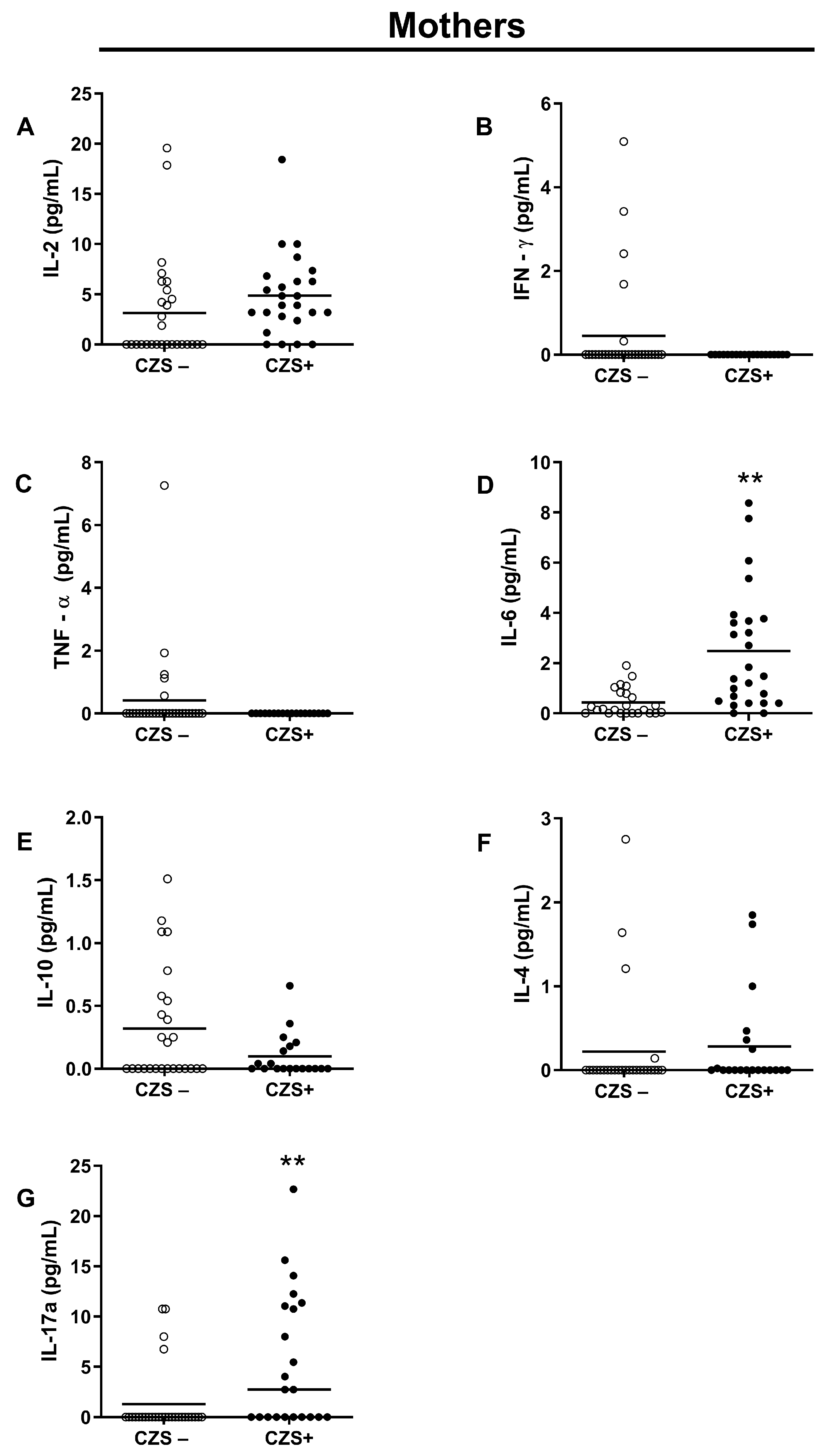
3.2. CZS+ Children Present Augmented IL-17 Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zanluca, C.; De Melo, V.C.A.; Mosimann, A.L.P.; Dos Santos, G.I.V.; dos Santos, C.N.D.; Luz, K. First Report of Autochthonous Transmission of Zika Virus in Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Fantinato, F.F.S.T.; Araújo, E.L.L.; Ribeiro, I.G.; de Andrade, M.R.; de Dantas, A.L.M.; Rios, J.M.T.; da Silva, O.M.V.; da Silva, M.D.S.; Nóbrega, R.V.; de Batista, D.A.; et al. Descrição Dos Primeiros Casos de Febre Pelo Vírus Zika Investigados Em Municípios Da Região Nordeste Do Brasil, 2015. Epidemiol. Serviços Saúde 2016, 25, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Faria, N.R.; da Silva Azevedo, R.D.S.; Kraemer, M.U.G.; Souza, R.; Cunha, M.S.; Hill, S.C.; Thézé, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika Virus in the Americas: Early Epidemiological and Genetic Findings. Science 2016, 352, aaf5036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasil, P.; Pereira, J.P., Jr.; Moreira, M.E.; Ribeiro Nogueira, R.M.; Damasceno, L.; Wakimoto, M.; Rabello, R.S.; Valderramos, S.G.; Halai, U.-A.; Salles, T.S.; et al. Zika Virus Infection in Pregnant Women in Rio de Janeiro. N. Engl. J. Med. 2016, 375, 2321–2334. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.A.; Staples, J.E.; Dobyns, W.B.; Pessoa, A.; Ventura, C.V.; Da Fonseca, E.B.; Ribeiro, E.M.; Ventura, L.O.; Neto, N.N.; Arena, J.F.; et al. Characterizing the Pattern of Anomalies in Congenital Zika Syndrome for Pediatric Clinicians. JAMA Pediatr. 2017, 171, 288–295. [Google Scholar] [CrossRef] [Green Version]
- de Noronha, L.; Zanluca, C.; Azevedo, M.L.V.; Luz, K.G.; dos Santos, C.N.D. Zika Virus Damages the Human Placental Barrier and Presents Marked Fetal Neurotropism. Mem. Inst. Oswaldo Cruz 2016, 111, 287–293. [Google Scholar] [CrossRef]
- Li, C.; Xu, D.; Ye, Q.; Hong, S.; Jiang, Y.; Liu, X.; Zhang, N.; Shi, L.; Qin, C.F.; Xu, Z. Correction: Zika Virus Disrupts Neural Progenitor Development and Leads to Microcephaly in Mice. Cell Stem Cell 2016, 19, 672. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, L.S.; Marques, R.E.; De Jesus, A.M.R.; De Almeida, R.P.; Teixeira, M.M. Zika Crisis in Brazil: Challenges in Research and Development. Curr. Opin. Virol. 2016, 18, 76–81. [Google Scholar] [CrossRef]
- Tricarico, P.M.; Caracciolo, I.; Crovella, S.; D’Agaro, P. Zika Virus Induces Inflammasome Activation in the Glial Cell Line U87-MG. Biochem. Biophys. Res. Commun. 2017, 492, 597–602. [Google Scholar] [CrossRef]
- Levine, D.; Jani, J.C.; Castro-Aragon, I.; Cannie, M. How Does Imaging of Congenital Zika Compare with Imaging of Other TORCH Infections? Radiology 2017, 285, 744–761. [Google Scholar] [CrossRef] [Green Version]
- França, G.; Schuler-Faccini, L.; Oliveira, W.K.; Henriques, C.M.P.; Carmo, E.H.; Pedi, V.D.; Nunes, M.L.; Castro, M.C.; Serruya, S.; Silveira, M.F.; et al. Congenital Zika Virus Syndrome in Brazil: A Case Series of the First 1501 Livebirths with Complete Investigation. Lancet 2016, 388, 891–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, D.; Gubler, D.J.; Schaub, B.; Lanteri, M.C.; Musso, D. An Update on Zika Virus Infection. Lancet 2017, 390, 2099–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, S.A.; Jamieson, D.J.; Honein, M.A.; Petersen, L.R. Zika Virus and Birth Defects—Reviewing the Evidence for Causality. N. Engl. J. Med. 2016, 374, 1981–1987. [Google Scholar] [CrossRef]
- Manangeeswaran, M.; Ireland, D.D.C.; Verthelyi, D. Zika (PRVABC59) Infection Is Associated with T Cell Infiltration and Neurodegeneration in CNS of Immunocompetent Neonatal C57Bl/6 Mice. PLoS Pathog. 2016, 12, e1006004. [Google Scholar] [CrossRef] [PubMed]
- Xavier-Neto, J.; Carvalho, M.; dos Pascoalino, B.S.; Cardoso, A.C.; Costa, Â.M.S.; Pereira, A.H.M.; Santos, L.N.; Saito, Â.; Marques, R.E.; Smetana, J.H.C.; et al. Hydrocephalus and Arthrogryposis in an Immunocompetent Mouse Model of ZIKA Teratogeny: A Developmental Study. PLoS Negl. Trop. Dis. 2017, 11, e0005363. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, R.S.S.; de Sousa, J.R.; Araujo, M.T.F.; Martins Filho, A.J.; de Alcantara, B.N.; Araujo, F.M.C.; Queiroz, M.G.L.; Cruz, A.C.R.; Vasconcelos, B.H.B.; Chiang, J.O.; et al. In Situ Immune Response and Mechanisms of Cell Damage in Central Nervous System of Fatal Cases Microcephaly by Zika Virus. Sci. Rep. 2018, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ornelas, A.M.M.; Pezzuto, P.; Silveira, P.P.; Melo, F.O.; Ferreira, T.A.; Oliveira-Szejnfeld, P.S.; Leal, J.I.; Amorim, M.M.R.; Hamilton, S.; Rawlinson, W.D.; et al. Immune Activation in Amniotic Fluid from Zika Virus-Associated Microcephaly. Ann. Neurol. 2017, 81, 152–156. [Google Scholar] [CrossRef]
- Naveca, F.G.; Pontes, G.S.; Chang, A.Y.; da Silva, G.A.V.; do Nascimento, V.A.; da Monteiro, D.C.S.; da Silva, M.S.; Abdalla, L.F.; Santos, J.H.A.; de Almeida, T.A.P.; et al. Analysis of the Immunological Biomarker Profile during Acute Zika Virus Infection Reveals the Overexpression of CXCL10, a Chemokine Linked to Neuronal Damage. Mem. Inst. Oswaldo Cruz 2018, 113, e170542. [Google Scholar] [CrossRef]
- Lum, F.M.; Low, D.K.S.; Fan, Y.; Tan, J.J.L.; Lee, B.; Chan, J.K.Y.; Rénia, L.; Ginhoux, F.; Ng, L.F.P. Zika Virus Infects Human Fetal Brain Microglia and Induces Inflammation. Clin. Infect. Dis. 2017, 64, 914–920. [Google Scholar] [CrossRef] [Green Version]
- Foo, S.S.; Chen, W.; Chan, Y.; Bowman, J.W.; Chang, L.C.; Choi, Y.; Yoo, J.S.; Ge, J.; Cheng, G.; Bonnin, A.; et al. Asian Zika Virus Strains Target CD14+blood Monocytes and Induce M2-Skewed Immunosuppression during Pregnancy. Nat. Microbiol. 2017, 2, 1558–1570. [Google Scholar] [CrossRef] [Green Version]
- Camacho-Zavala, E.; Santacruz-Tinoco, C.; Muñoz, E.; Chacón-Salinas, R.; Salazar-Sanchez, M.I.; Grajales, C.; González-Ibarra, J.; Borja-Aburto, V.H.; Jaenisch, T.; Gonzalez-Bonilla, C.R. Pregnant Women Infected with Zika Virus Show Higher Viral Load and Immunoregulatory Cytokines Profile with CXCL10 Increase. Viruses 2021, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Badolato-Corrêa, J.; Carvalho, F.R.; Paiva, I.A.; Familiar-Macedo, D.; Dias, H.G.; Pauvolid-Corrêa, A.; Fernandes-Santos, C.; da Lima, M.R.Q.; Gandini, M.; Silva, A.A.; et al. Differential Longevity of Memory CD4 and CD8 T Cells in a Cohort of the Mothers With a History of ZIKV Infection and Their Children. Front. Immunol. 2021, 12, 610456. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.A.M.; Ganz, J.S.S.; da Sousa, P.S.; Doriqui, M.J.R.; Ribeiro, M.R.C.; dos Branco, M.R.F.C.; de Queiroz, R.C.S.; de Pacheco, M.J.T.; Vieira da Costa, F.R.; de Silva, F.S.; et al. Early Growth and Neurologic Outcomes of Infants with Probable Congenital Zika Virus Syndrome. Emerg. Infect. Dis. 2016, 22, 1953–1956. [Google Scholar] [CrossRef]
- Baer, A.; Kehn-Hall, K. Viral Concentration Determination through Plaque Assays: Using Traditional and Novel Overlay Systems. J. Vis. Exp. 2014, 93, e52065. [Google Scholar] [CrossRef]
- Fialho, E.M.S.; Veras, E.M.; de Jesus, C.M.; Khouri, R.; Sousa, P.S.; Ribeiro, M.R.C.; Costa, L.C.; Gomes, L.N.; Nascimento, F.R.F.; Silva, A.A.M.; et al. Maternal Immune Response to ZIKV Triggers High-Inflammatory Profile in Congenital Zika Syndrome. Viruses 2023, 15, 220. [Google Scholar] [CrossRef] [PubMed]
- de Melo, A.S.O.; Aguiar, R.S.; Amorim, M.M.R.; Arruda, M.B.; de Melo, F.O.; Ribeiro, S.T.C.; Batista, A.G.M.; Ferreira, T.; dos Santos, M.P.; Sampaio, V.V.; et al. Congenital Zika Virus Infection: Beyond Neonatal Microcephaly. JAMA Neurol. 2016, 73, 1407–1416. [Google Scholar] [CrossRef] [Green Version]
- de França, T.L.B.; Medeiros, W.R.; de Souza, N.L.; Longo, E.; Pereira, S.A.; de Oliveira França, T.B.; Sousa, K.G. Growth and Development of Children with Microcephaly Associated with Congenital Zika Virus Syndrome in Brazil. Int. J. Environ. Res. Public Health 2018, 15, 1990. [Google Scholar] [CrossRef] [Green Version]
- Cauchemez, S.; Besnard, M.; Bompard, P.; Dub, T.; Guillemette-Artur, P.; Eyrolle-Guignot, D.; Salje, H.; Van Kerkhove, M.D.; Abadie, V.; Garel, C.; et al. Association between Zika Virus and Microcephaly in French Polynesia, 2013–2015: A Retrospective Study. Lancet 2016, 387, 2125–2132. [Google Scholar] [CrossRef] [Green Version]
- van der Linden, V.; Pessoa, A.; Dobyns, W.; Barkovich, A.J.; van der Júnior, H.L.; Filho, E.L.R.; Ribeiro, E.M.; de Leal, M.C.; de Coimbra, P.P.A.; de Aragão, M.F.V.V.; et al. Description of 13 Infants Born during October 2015–January 2016 with Congenital Zika Virus Infection without Microcephaly at Birth—Brazil. MMWR. Morb. Mortal. Wkly. Rep. 2016, 65, 1343–1348. [Google Scholar] [CrossRef]
- Rice, M.E.; Galang, R.R.; Roth, N.M.; Ellington, S.R.; Moore, C.A.; Valencia-Prado, M.; Ellis, E.M.; Tufa, A.J.; Taulung, L.A.; Alfred, J.M.; et al. Vital Signs: Zika-Associated Birth Defects and Neurodevelopmental Abnormalities Possibly Associated with Congenital Zika Virus Infection—U.S. Territories and Freely Associated States, 2018. MMWR. Morb. Mortal. Wkly. Rep. 2018, 67, 858–868. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, M.R.C.; Khouri, R.; Sousa, P.S.; Branco, M.R.F.C.; Batista, R.F.L.; Costa, E.P.F.; Alves, M.T.S.S.B.; Amaral, G.A.; Borges, M.C.R.; Takahasi, E.H.M.; et al. Plaque Reduction Neutralization Test (PRNT) in the Congenital Zika Syndrome: Positivity and Associations with Laboratory, Clinical, and Imaging Characteristics. Viruses 2020, 12, 1244. [Google Scholar] [CrossRef] [PubMed]
- Mendes, A.K.T.; Ribeiro, M.R.C.; Lamy-Filho, F.; Amaral, G.A.; Borges, M.C.R.; Costa, L.C.; Cavalcante, T.B.; Batista, R.F.L.; da Sousa, P.S.; da Silva, A.A.M. Congenital Zika Syndrome: Association between the Gestational Trimester of Maternal Infection, Severity of Brain Computed Tomography Findings and Microcephaly at Birth. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Paixao, E.S.; Leong, W.-Y.; Rodrigues, L.C.; Wilder-Smith, A. Asymptomatic Prenatal Zika Virus Infection and Congenital Zika Syndrome. Open Forum Infect. Dis. 2018, 5, ofy073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Linden, V.; de Lima Petribu, N.C.; Pessoa, A.; Faquini, I.; Paciorkowski, A.R.; van der Linden, H., Jr.; Silveira-Moriyama, L.; Cordeiro, M.T.; Hazin, A.N.; Barkovich, A.J.; et al. Association of Severe Hydrocephalus With Congenital Zika Syndrome. JAMA Neurol. 2019, 76, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Pardy, R.D.; Richer, M.J. Protective to a T: The Role of T Cells during Zika Virus Infection. Cells 2019, 8, 820. [Google Scholar] [CrossRef] [Green Version]
- Vyas, A.K.; Negi, P.; Patra, S.; Maras, J.S.; Ramakrishna, G.; Sarin, S.K.; Trehanpati, N. Maternal Immunity Influences Vertical Transmission of Hepatitis B to Newborns. Hepatol. Commun. 2019, 3, 795–811. [Google Scholar] [CrossRef] [Green Version]
- Cugola, F.R.; Fernandes, I.R.; Russo, F.B.; Freitas, B.C.; Dias, J.L.M.; Guimarães, K.P.; Benazzato, C.; Almeida, N.; Pignatari, G.C.; Romero, S.; et al. The Brazilian Zika Virus Strain Causes Birth Defects in Experimental Models. Nature 2016, 534, 15. [Google Scholar] [CrossRef] [Green Version]
- Azamor, T.; Cunha, D.P.; da Silva, A.M.V.; de Bezerra, O.C.L.; Ribeiro-Alves, M.; Calvo, T.L.; de Kehdy, F.S.G.; de Manta, F.S.N.; de Pinto, T.G.T.; Ferreira, L.P.; et al. Congenital Zika Syndrome Is Associated with Interferon Alfa Receptor 1. Front. Immunol. 2021, 12, 5029. [Google Scholar] [CrossRef]
- Cornish, E.F.; Filipovic, I.; Åsenius, F.; Williams, D.J.; McDonnell, T. Innate Immune Responses to Acute Viral Infection during Pregnancy. Front. Immunol. 2020, 11, 572567. [Google Scholar] [CrossRef]
- Yockey, L.J.; Iwasaki, A. Role of Interferons and Cytokines in Pregnancy and Fetal Development. Immunity 2018, 49, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Choi, G.B.; Yim, Y.S.; Wong, H.; Kim, S.; Kim, H.; Kim, S.V.; Hoeffer, C.A.; Littman, D.R.; Huh, J.R. The Maternal Interleukin-17a Pathway in Mice Promotes Autism-like Phenotypes in Offspring. Science 2016, 351, 933–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihara, M.; Hashizume, M.; Yoshida, H.; Suzuki, M.; Shiina, M. IL-6/IL-6 Receptor System and Its Role in Physiological and Pathological Conditions. Clin. Sci. 2011, 122, 143–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.E.P.; Li, J.; Garbett, K.; Mirnics, K.; Patterson, P.H. Maternal Immune Activation Alters Fetal Brain Development through Interleukin-6. J. Neurosci. 2007, 27, 10695–10702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrode-Brusés, G.; Brusés, J.L. Maternal Immune Activation by Poly(I:C) Induces Expression of Cytokines IL-1β and IL-13, Chemokine MCP-1 and Colony Stimulating Factor VEGF in Fetal Mouse Brain. J. Neuroinflamm. 2012, 9, 83. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, Y.; Tsukada, T.; Sakata-Haga, H.; Sakai, D.; Shoji, H.; Saikawa, Y.; Hatta, T. Exposure to Maternal Immune Activation Causes Congenital Unfolded Protein Response Defects and Increases the Susceptibility to Postnatal Inflammatory Stimulation in Offspring. J. Inflamm. Res. 2021, 14, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Krebs, C.F.; Reimers, D.; Zhao, Y.; Paust, H.-J.; Bartsch, P.; Nuñez, S.; Rosemblatt, M.V.; Hellmig, M.; Kilian, C.; Borchers, A.; et al. Pathogen-Induced Tissue-Resident Memory T H 17 (T RM 17) Cells Amplify Autoimmune Kidney Disease. Sci. Immunol. 2020, 5, eaba4163. [Google Scholar] [CrossRef]
- Zhao, Y.; Kilian, C.; Turner, J.-E.; Bosurgi, L.; Roedl, K.; Bartsch, P.; Gnirck, A.-C.; Cortesi, F.; Schultheiß, C.; Hellmig, M.; et al. Clonal Expansion and Activation of Tissue-Resident Memory-like Th17 Cells Expressing GM-CSF in the Lungs of Severe COVID-19 Patients. Sci. Immunol. 2021, 6, eabf6692. [Google Scholar] [CrossRef]
- Vesely, M.C.A.; Pallis, P.; Bielecki, P.; Low, J.S.; Zhao, J.; Harman, C.C.D.; Kroehling, L.; Jackson, R.; Bailis, W.; Licona-Limón, P.; et al. Effector TH17 Cells Give Rise to Long-Lived TRM Cells That Are Essential for an Immediate Response against Bacterial Infection. Cell 2019, 178, 1176–1188.e15. [Google Scholar] [CrossRef]
- Brown, A.S. Epidemiologic Studies of Exposure to Prenatal Infection and Risk of Schizophrenia and Autism. Dev. Neurobiol. 2012, 72, 1272–1276. [Google Scholar] [CrossRef] [Green Version]
- Ireland, D.D.C.; Manangeeswaran, M.; Lewkowicz, A.P.; Engel, K.; Clark, S.M.; Laniyan, A.; Sykes, J.; Lee, H.N.; McWilliams, I.L.; Kelley-Baker, L.; et al. Long-Term Persistence of Infectious Zika Virus: Inflammation and Behavioral Sequela in Mice. PLoS Pathog. 2020, 16, e1008689. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fialho, E.M.S.; Veras, E.M.; Jesus, C.M.d.; Gomes, L.N.; Khouri, R.; Sousa, P.S.; Ribeiro, M.R.C.; Batista, R.F.L.; Costa, L.C.; Nascimento, F.R.F.; et al. Maternal Th17 Profile after Zika Virus Infection Is Involved in Congenital Zika Syndrome Development in Children. Viruses 2023, 15, 1320. https://doi.org/10.3390/v15061320
Fialho EMS, Veras EM, Jesus CMd, Gomes LN, Khouri R, Sousa PS, Ribeiro MRC, Batista RFL, Costa LC, Nascimento FRF, et al. Maternal Th17 Profile after Zika Virus Infection Is Involved in Congenital Zika Syndrome Development in Children. Viruses. 2023; 15(6):1320. https://doi.org/10.3390/v15061320
Chicago/Turabian StyleFialho, Eder M. S., Emanoel M. Veras, Caroline M. de Jesus, Líllian N. Gomes, Ricardo Khouri, Patrícia S. Sousa, Marizélia R. C. Ribeiro, Rosângela F. L. Batista, Luciana C. Costa, Flávia R. F. Nascimento, and et al. 2023. "Maternal Th17 Profile after Zika Virus Infection Is Involved in Congenital Zika Syndrome Development in Children" Viruses 15, no. 6: 1320. https://doi.org/10.3390/v15061320
APA StyleFialho, E. M. S., Veras, E. M., Jesus, C. M. d., Gomes, L. N., Khouri, R., Sousa, P. S., Ribeiro, M. R. C., Batista, R. F. L., Costa, L. C., Nascimento, F. R. F., Silva, A. A. M., & Soeiro-Pereira, P. V. (2023). Maternal Th17 Profile after Zika Virus Infection Is Involved in Congenital Zika Syndrome Development in Children. Viruses, 15(6), 1320. https://doi.org/10.3390/v15061320