

High-Level Drug-Resistant Mutations among HIV-1 Subtype A6 and CRF02_AG in Kazakhstan

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval, Study Design, and Population
2.2. Epidemiological Data
2.3. Sequence Generation
2.4. Sequence Subtyping
2.5. Drug Resistance Profiling
2.6. Phylogenetic Analysis
2.7. Statistical Analysis
3. Results
3.1. Demographic Characterization of Study Population
3.2. Drug Resistance Profiles
3.3. Distinct DRMs among Subtypes A6 and CRF02_AG
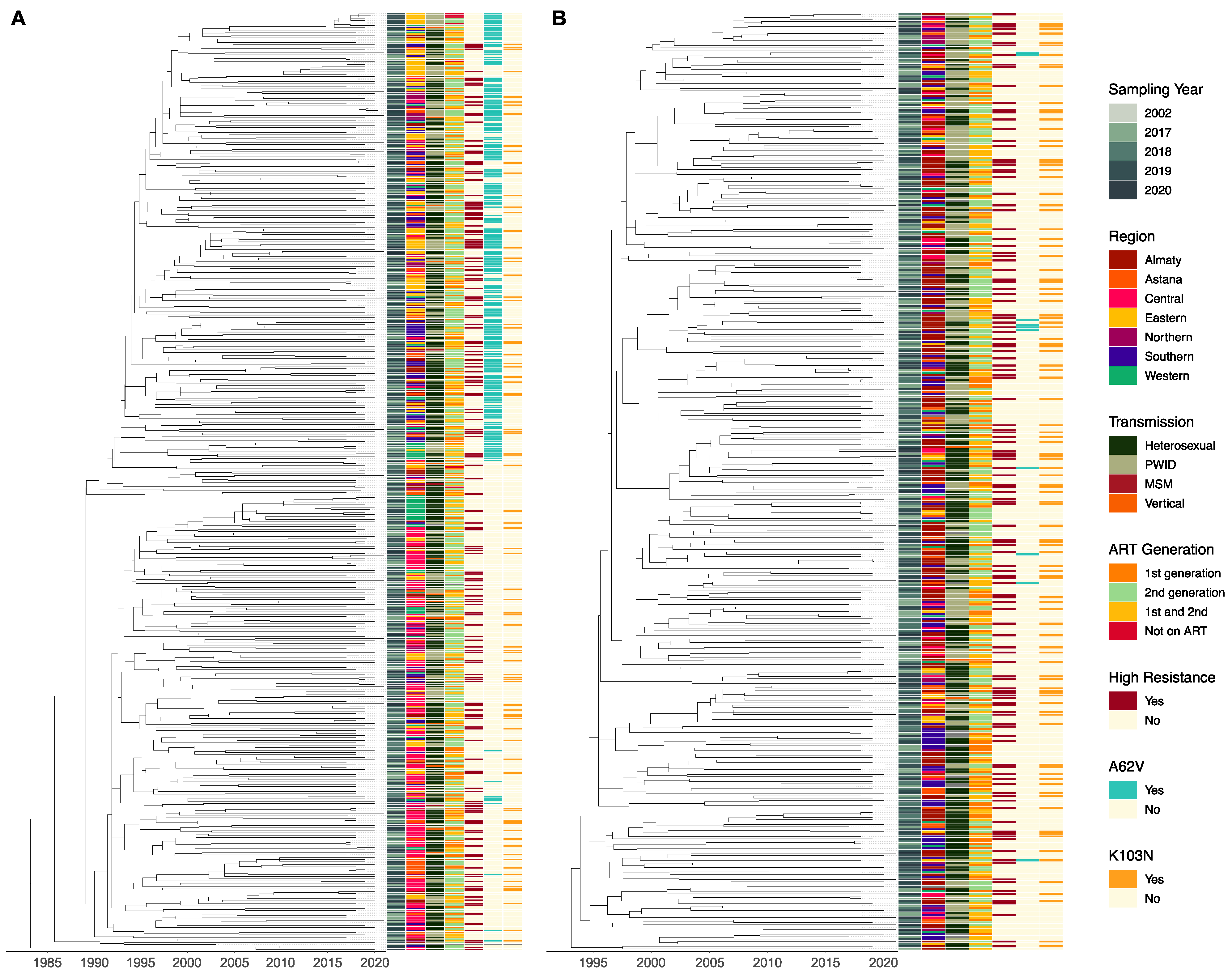
3.4. Phylogenetic Analysis of Subtypes A6 and CRF02_AG
4. Discussion
4.1. Demographics of the HIV-1 Epidemic
4.2. High-Level Drug Resistance and Distinct DRMs between the Subtypes
4.3. Evolutionary History of the HIV-1 Epidemic in Kazakhstan
4.4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- UN Joint Programme on HIV/AIDS IN DANGER; UNAIDS Global AIDS Update 2022; UNAIDS: Geneva, Switzerland, 2022.
- The World Bank Data for Kazakhstan, Uzbekistan, Turkmenistan, Tajikistan, Kyrgyz Republic. Available online: https://data.worldbank.org/?locations=KZ-UZ-TM-TJ-KG (accessed on 13 January 2023).
- UN Joint Programme on HIV/AIDS UNAIDS Data 2021; UNAIDS: Geneva, Switzerland, 2021.
- World Health Organisation Consolidated Guidelines on HIV Prevention, Testing, Treatment, Service Delivery and Monitoring; Recommendations for a Public Health Approach; World Health Organisation: Geneva, Switzerland, 2021.
- Amangaldiyeva, A.; Davlidova, S.; Baiserkin, B.; Dzissyuk, N.; DeHovitz, J.; Ali, S. Implementation of antiretroviral therapy (ART) in former Soviet Union countries. AIDS Res. Ther. 2019, 16, 35. [Google Scholar] [CrossRef] [PubMed]
- International Treatment Preparedness Coalition Eastern Europe and Central Asia Clinical Protocol for the Diagnosis and Treatment of HIV Infection in Adults. Available online: https://itpc-eeca.org/2018/06/19/klinicheskij-protokol-diagnostiki-i-lecheniya-vich-infektsii-u-vzroslyh/ (accessed on 13 January 2023). (In Russian).
- Mukhatayeva, A.; Mustafa, A.; Dzissyuk, N.; Issanov, A.; Mukhatayev, Z.; Bayserkin, B.; Vermund, S.H.; Ali, S. Antiretroviral therapy resistance mutations among HIV infected people in Kazakhstan. Sci. Rep. 2022, 12, 17195. [Google Scholar] [CrossRef] [PubMed]
- International Treatment Preparedness Coalition Eastern Europe and Central Asia Recommendations for the Diagnosis and Treatment Od HIV Infection in Adults and Children Have Been Updated in Kazakhstan. Available online: https://itpc-eeca.org/2020/06/17/v-kazahstane-obnovleny-rekomendaczii-po-diagnostike-i-lecheniyu-vich-infekczii-u-vzroslyh-i-detej/ (accessed on 13 January 2023). (In Russian).
- World Health Organisation HIV Drug Resistance Report; World Health Organisation: Geneva, Switzerland, 2021.
- Cambiano, V.; Bertagnolio, S.; Jordan, M.R.; Pillay, D.; Perriëns, J.H.; Venter, F.; Lundgren, J.; Phillips, A. redicted Levels of HIV Drug Resistance: Potential Impact of Expanding Diagnosis, Retention, and Eligibility Criteria for Antiretroviral Therapy Initiation. AIDS 2014, 28, S15–S23. [Google Scholar] [CrossRef] [PubMed]
- Clutter, D.S.; Jordan, M.R.; Bertagnolio, S.; Shafer, R.W. HIV-1 drug resistance and resistance testing. Infect. Genet. Evol. 2016, 46, 292–307. [Google Scholar] [CrossRef] [Green Version]
- Clavel, F.; Hance, A.J. HIV Drug Resistance. N. Engl. J. Med. 2004, 350, 1023–1035. [Google Scholar] [CrossRef]
- Lapovok, I.; Kazennova, E.; Laga, V.; Vasilyev, A.; Utegenova, A.; Abishev, A.; Dzissyuk, N.; Tukeev, M.; Bobkova, M. Short Communication: Molecular Epidemiology of HIV Type 1 Infection in Kazakhstan: CRF02_AG Prevalence Is Increasing in the Southeastern Provinces. AIDS Res. Hum. Retroviruses 2014, 30, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Eyzaguirre, L.M.; Erasilova, I.B.; Nadai, Y.; Saad, M.D.; Kovtunenko, N.G.; Gomatos, P.J.; Zeman, V.V.; A Botros, B.; Sanchez, J.L.; Birx, D.L.; et al. Genetic Characterization of HIV-1 Strains Circulating in Kazakhstan. J. Acquir. Immune Defic. Syndr. 2007, 46, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Aibekova, L.; Foley, B.; Hortelano, G.; Raees, M.; Abdraimov, S.; Toichuev, R.; Ali, S. Molecular epidemiology of HIV-1 subtype A in former Soviet Union countries. PLoS ONE 2018, 13, e0191891. [Google Scholar] [CrossRef] [Green Version]
- Abidi, S.H.; Aibekova, L.; Davlidova, S.; Amangeldiyeva, A.; Foley, B.; Ali, S. Origin and evolution of HIV-1 subtype A6. PLoS ONE 2021, 16, e0260604. [Google Scholar] [CrossRef]
- Sivay, M.V.; Totmenin, A.V.; Zyryanova, D.P.; Osipova, I.P.; Nalimova, T.M.; Gashnikova, M.P.; Ivlev, V.V.; Meshkov, I.O.; Chokmorova, U.Z.; Narmatova, E.; et al. Characterization of HIV-1 Epidemic in Kyrgyzstan. Front. Microbiol. 2021, 12, 753675. [Google Scholar] [CrossRef]
- Pineda-Peña, A.-C.; Faria, N.R.; Imbrechts, S.; Libin, P.; Abecasis, A.B.; Deforche, K.; Gómez-López, A.; Camacho, R.J.; de Oliveira, T.; Vandamme, A.-M. Automated subtyping of HIV-1 genetic sequences for clinical and surveillance purposes: Performance evaluation of the new REGA version 3 and seven other tools. Infect. Genet. Evol. 2013, 19, 337–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pond, S.L.K.; Posada, D.; Stawiski, E.; Chappey, C.; Poon, A.; Hughes, G.; Fearnhill, E.; Gravenor, M.B.; Brown, A.L.; Frost, S. An Evolutionary Model-Based Algorithm for Accurate Phylogenetic Breakpoint Mapping and Subtype Prediction in HIV-1. PLOS Comput. Biol. 2009, 5, e1000581. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.Y.; Gonzales, M.J.; Kantor, R.; Betts, B.J.; Ravela, J.; Shafer, R.W. Human immunodeficiency virus reverse transcriptase and protease sequence database. Nucleic Acids Res. 2003, 31, 298–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.F.; Shafer, R.W. Web Resources for HIV Type 1 Genotypic-Resistance Test Interpretation. Clin. Infect. Dis. 2006, 42, 1608–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadfield, J.; Megill, C.; Bell, S.M.; Huddleston, J.; Potter, B.; Callender, C.; Sagulenko, P.; Bedford, T.; Neher, R.A. Nextstrain: Real-time tracking of pathogen evolution. Bioinformatics 2018, 34, 4121–4123. [Google Scholar] [CrossRef] [Green Version]
- Los Alamos National Laboratory HIV Sequence Database. Available online: http://www.hiv.lanl.gov/ (accessed on 13 January 2023).
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Sagulenko, P.; Puller, V.; Neher, R.A. TreeTime: Maximum-likelihood phylodynamic analysis. Virus Evol. 2018, 4, vex042. [Google Scholar] [CrossRef] [Green Version]
- R Core Team R: A Language and Environment for Statistical Computing 2021. Available online: https://www.R-project.org/ (accessed on 2 June 2023).
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Xu, S.; Dai, Z.; Guo, P.; Fu, X.; Liu, S.; Zhou, L.; Tang, W.; Feng, T.; Chen, M.; Zhan, L.; et al. ggtreeExtra: Compact Visualization of Richly Annotated Phylogenetic Data. Mol. Biol. Evol. 2021, 38, 4039–4042. [Google Scholar] [CrossRef] [PubMed]
- Agency for Strategic Planning and Reforms of the Republic of Kazakhstan-Bureau of National Statistics Demographic Statistics. Available online: https://stat.gov.kz/official/industry/61/statistic/8 (accessed on 13 January 2023).
- Davlidova, S.; Haley-Johnson, Z.; Nyhan, K.; Farooq, A.; Vermund, S.H.; Ali, S. Prevalence of HIV, HCV and HBV in Central Asia and the Caucasus: A systematic review. Int. J. Infect. Dis. 2020, 104, 510–525. [Google Scholar] [CrossRef] [PubMed]
- Center for Prevention and Control of AIDS Dynamics of HIV Transmission Routes in Kazakhstan. Available online: https://gcaids.kz/ru/ctatistika (accessed on 13 January 2023). (In Russian).
- McCrimmon, T.; Sundelson, A.; Darisheva, M.; Gilbert, L.; Hunt, T.; Terlikbayeva, A.; Primbetova, S.; El-Bassel, N. HIV Care Continuum Services for People Who Inject Drugs in Kazakhstan During COVID-19: A Qualitative Study of Service Provider Perspectives. Glob. Health Sci. Pract. 2022, 10, e2100619. [Google Scholar] [CrossRef] [PubMed]
- El-Bassel, N.; McCrimmon, T.; Wu, E.; Chang, M.; Terlikbayeva, A.; Hunt, T.; Darisheva, M.; Primbetova, S.; Davis, A.; Metsch, L.R.; et al. Effectiveness of an Intervention to Improve HIV Service Delivery for People Who Inject Drugs in Kazakhstan: A Cluster Trial. JAMA Netw. Open 2022, 5, e2244734. [Google Scholar] [CrossRef]
- Carr, J.K.; Nadai, Y.; Eyzaguirre, L.; Saad, M.D.; Khakimov, M.M.; Yakubov, S.K.; Birx, D.L.; Graham, R.R.; Wolfe, N.D.; Earhart, K.C.; et al. Outbreak of a West African recombinant of HIV-1 in Tashkent, Uzbekistan. J. Acquir. Immune Defic. Syndr. 2005, 39, 570–575. [Google Scholar]
- Zarandia, M.; Tsertsvadze, T.; Carr, J.K.; Nadai, Y.; Sanchez, J.L.; Nelson, K. HIV-1 Genetic Diversity and Genotypic Drug Susceptibility in the Republic of Georgia. AIDS Res. Hum. Retroviruses 2006, 22, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Bacheler, L.; Jeffrey, S.; Hanna, G.; D’Aquila, R.; Wallace, L.; Logue, K.; Cordova, B.; Hertogs, K.; Larder, B.; Buckery, R.; et al. Genotypic Correlates of Phenotypic Resistance to Efavirenz in Virus Isolates from Patients Failing Nonnucleoside Reverse Transcriptase Inhibitor Therapy. J. Virol. 2001, 75, 4999–5008. [Google Scholar] [CrossRef] [Green Version]
- Sukhanova, A.L.; Bogoslovskaia, E.V.; Kruglova, A.I.; Bashkirova, L.I.; Tsyganova, G.M.; Shipulin, G.A.; Kazennova, E.V.; Alikina, I.I.; Zverev, S.I.; Grishechkin, A.E.; et al. Protease and Reverse Transcriptase Genetic Polymorphism in HIV Type 1 Subtype A Variants Predominating in CIS Countries. Mol. Biol. 2005, 39, 1063–1071. (In Russian) [Google Scholar] [CrossRef]
- Schlösser, M.; Kartashev, V.V.; Mikkola, V.H.; Shemshura, A.; Saukhat, S.; Kolpakov, D.; Suladze, A.; Tverdokhlebova, T.; Hutt, K.; Heger, E.; et al. HIV-1 Sub-Subtype A6: Settings for Normalised Identification and Molecular Epidemiology in the Southern Federal District, Russia. Viruses 2020, 12, 475. [Google Scholar] [CrossRef] [Green Version]
- Kolomeets, A.N.; Varghese, V.; Lemey, P.; Bobkova, M.; Shafer, R.W. A uniquely prevalent nonnucleoside reverse transcriptase inhibitor resistance mutation in Russian subtype A HIV-1 viruses. AIDS 2014, 28, F1–F8. [Google Scholar] [CrossRef] [Green Version]
- Bobkova, M. Current status of HIV-1 diversity and drug resistance monitoring in the former USSR. AIDS Rev. 2013, 15, 204–212. [Google Scholar] [PubMed]
- Aldridge, R.W.; Zenner, D.; White, P.J.; Muzyamba, M.C.; Loutet, M.; Dhavan, P.; Mosca, D.; Hayward, A.C.; Abubakar, I. Prevalence of and risk factors for active tuberculosis in migrants screened before entry to the UK: A population-based cross-sectional study. Lancet Infect. Dis. 2016, 16, 962–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díez-Fuertes, F.; Cabello, M.; Thomson, M.M. Bayesian phylogeographic analyses clarify the origin of the HIV-1 subtype A variant circulating in former Soviet Union’s countries. Infect. Genet. Evol. 2015, 33, 197–205. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Preferred | Alternative | Sources | |
|---|---|---|---|
| 2017–2020 | 3TC/FTC + TDF + EFV | 3TC + AZT + EFV/NVP | [5,6] |
| 3TC/FTC + TDF + DTG/NVP | |||
| 2020–present | 3TC/FTC + TDF/TAF + DTG/BIC | 3TC/FTC + TDF + EFV | [7,8] |
| 3TC + DTG | 3TC + ABC + EFV/DTG | ||
| 3TC/FTC + ABC/TDF + RAL | |||
| 3TC/FTC + TDF/TAF + EVG/c | |||
| 3TC/FTC + TDF/TAF/ABC + DRV/c/DRV/r |
| A6 | CRF02_AG | Other | Total | |
|---|---|---|---|---|
| n (%) | n (%) | n (%) | n (%) | |
| All sequences | 553 (57.1) | 401 (41.4) | 14 (1.4) | 968 (100) |
| Gender | ||||
| Female | 272 (49.2) | 190 (47.4) | 6 (42.9) | 468 (48.3) |
| Male | 281 (50.8) | 211 (52.6) | 8 (57.1) | 500 (51.7) |
| Sampling year | ||||
| 2017 | 124 (22.4) | 104 (25.9) | 1 (7.1) | 229 (23.7) |
| 2018 | 138 (25) | 110 (27.4) | 1 (7.1) | 249 (25.7) |
| 2019 | 240 (43.4) | 140 (34.9) | 9 (64.3) | 389 (40.2) |
| 2020 | 51 (9.2) | 47 (11.7) | 3 (21.4) | 101 (10.4) |
| Region | ||||
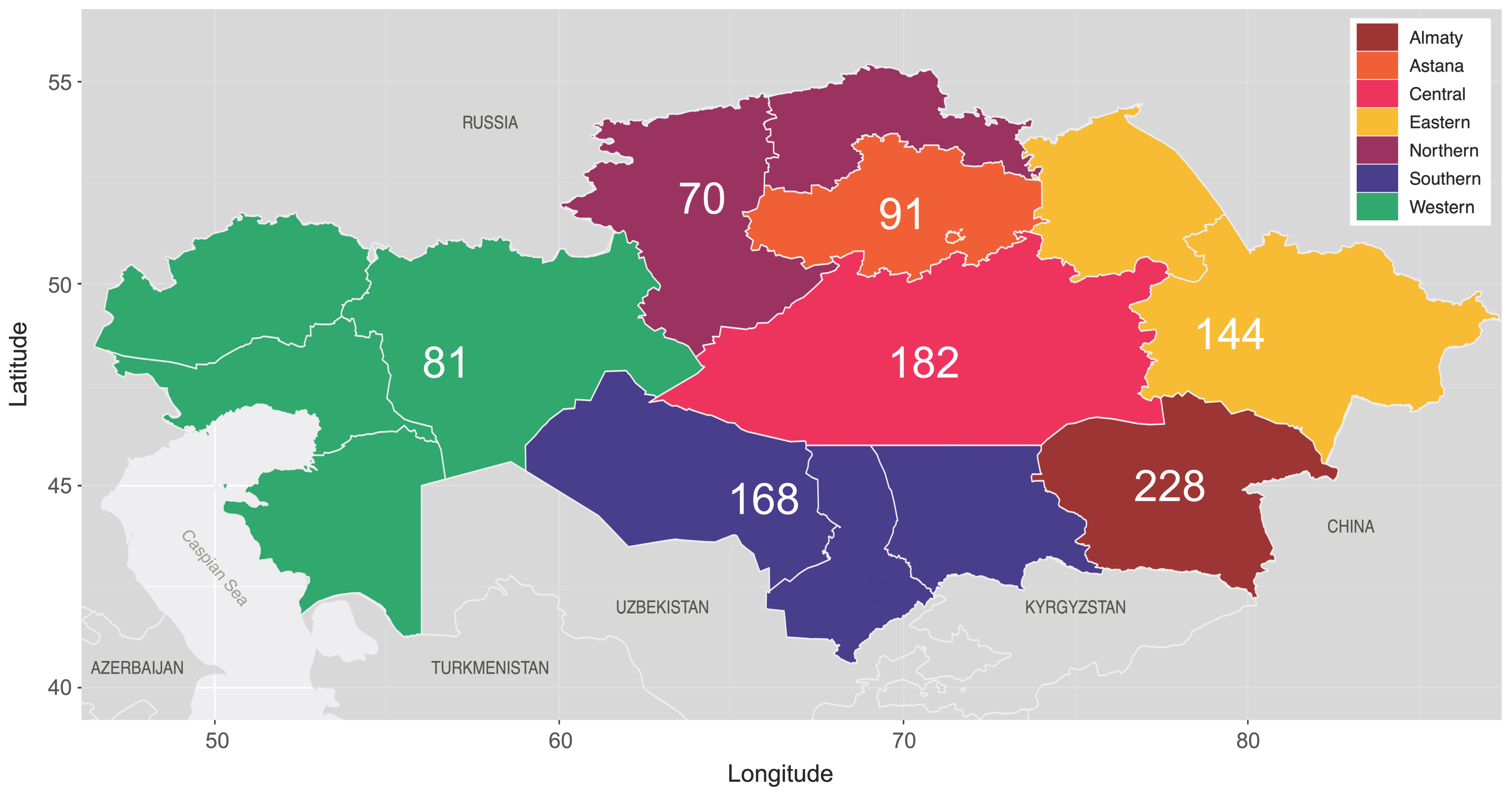
| Almaty | 49 (8.9) | 175 (43.6) | 4 (28.6) | 228 (23.6) |
| Astana | 58 (10.5) | 33 (8.2) | 0 | 91 (9.4) |
| Central Kazakhstan | 148 (26.8) | 33 (8.2) | 1 (7.1) | 182 (18.8) |
| Eastern Kazakhstan | 116 (21) | 26 (6.5) | 2 (14.3) | 144 (14.9) |
| Northern Kazakhstan | 45 (8.1) | 21 (5.2) | 4 (28.6) | 70 (7.2) |
| Southern Kazakhstan | 80 (14.5) | 86 (21.4) | 2 (14.3) | 168 (17.4) |
| Western Kazakhstan | 56 (10.1) | 24 (6) | 1 (7.1) | 81 (8.4) |
| NA | 1 (0.2) | 3 (0.7) | 0 | 4 (0.4) |
| Transmission 1 | ||||
| Heterosexual | 349 (63.1) | 209 (52.1) | 8 (57.1) | 566 (58.5) |
| PWID | 184 (33.3) | 177 (44.1) | 3 (21.4) | 364 (37.6) |
| MSM | 3 (0.5) | 0 | 2 (14.3) | 5 (0.5) |
| Vertical | 13 (2.4) | 3 (0.7) | 1 (7.1) | 17 (1.8) |
| NA | 4 (0.7) | 12 (3) | 0 | 16 (1.7) |
| ART regimen 2 | ||||
| FTC + TDF + EFV | 231 (41.8) | 148 (36.9) | 7 (50) | 386 (39.9) |
| 3TC + AZT + EFV | 58 (10.5) | 48 (12) | 0 | 106 (11) |
| 3TC + AZT + NVP | 60 (10.8) | 65 (16.2) | 0 | 125 (12.9) |
| Other | 196 (35.4) | 139 (34.7) | 7 (50) | 342 (35.3) |
| Not on ART | 7 (1.3) | 0 | 0 | 7 (0.7) |
| NA | 1 (0.2) | 1 (0.2) | 0 | 2 (0.2) |
| High-level resistance 3 | ||||
| Any | 172 (31.1) | 135 (33.7) | 4 (28.6) | 311 (32.1) |
| NRTI only 4 | 11 (2) | 9 (2.2) | 0 | 20 (2.1) |
| NNRTI only | 72 (13) | 73 (18.2) | 3 (21.4) | 148 (15.3) |
| NRTI + NNRTI 5 | 89 (16.1) | 53 (13.2) | 1 (7.1) | 143 (14.8) |
| A6 | CRF02_AG | Other | Total | |
|---|---|---|---|---|
| n (%) | n (%) | n (%) | n (%) | |
| All sequences | 553 (57.1) | 401 (41.4) | 14 (1.4) | 968 (100) |
| NRTI major | ||||
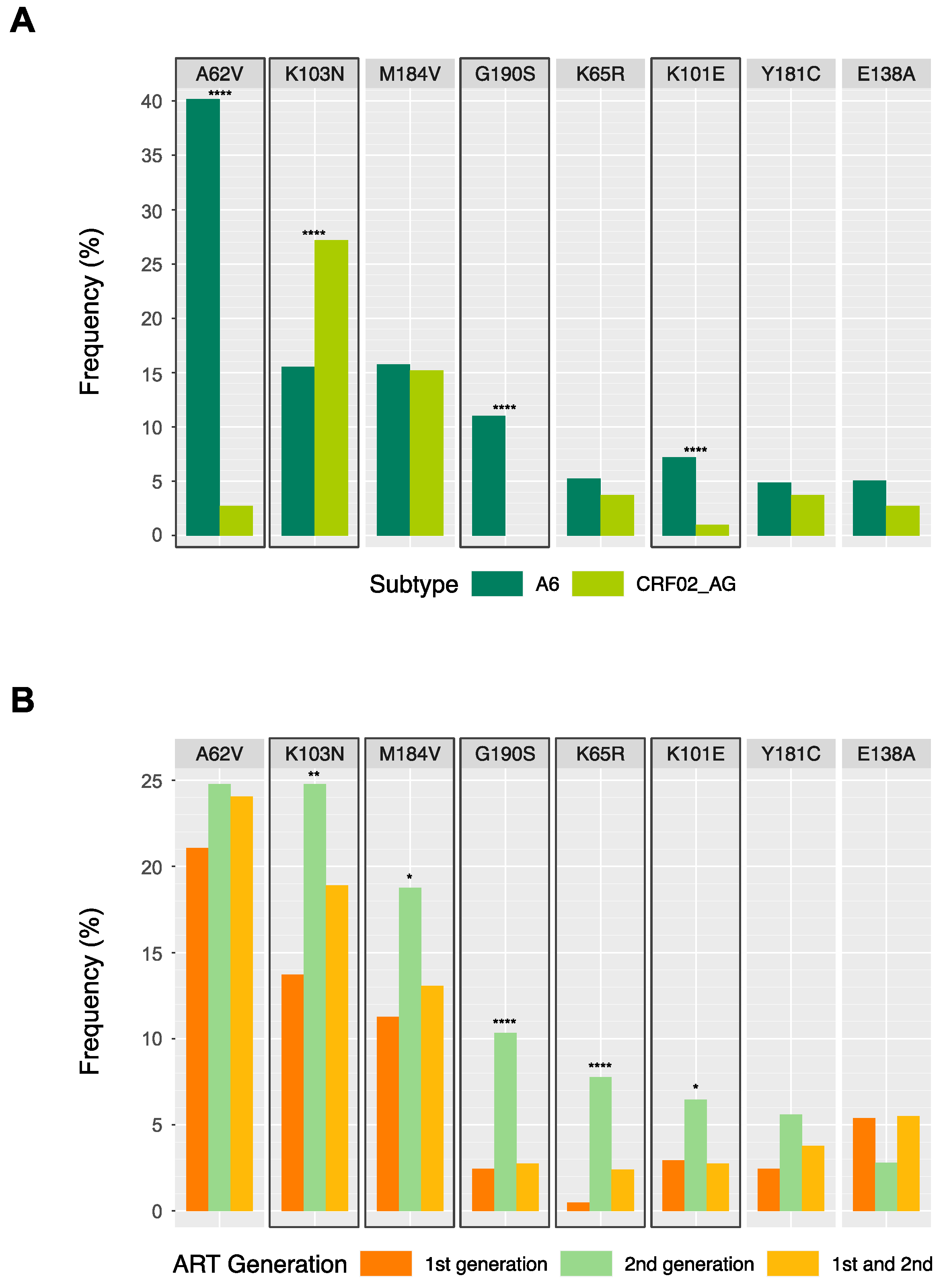
| M184V | 87 (15.7) | 61 (15.2) | 1 (7.1) | 149 (15.4) |
| K65R | 29 (5.2) | 15 (3.7) | 1 (7.1) | 45 (4.6) |
| D67N | 16 (2.9) | 1 (0.2) | 0 | 17 (1.8) |
| K70R | 10 (1.8) | 5 (1.2) | 1 (7.1) | 16 (1.7) |
| Y115F | 9 (1.6) | 6 (1.5) | 0 | 15 (1.5) |
| M41L | 8 (1.4) | 6 (1.5) | 0 | 14 (1.4) |
| K70E | 9 (1.6) | 4 (1) | 0 | 13 (1.3) |
| L74I | 10 (1.8) | 3 (0.7) | 0 | 13 (1.3) |
| L74V | 7 (1.3) | 4 (1) | 0 | 11 (1.1) |
| NRTI accessory | ||||
| A62V | 222 (40.1) | 11 (2.7) | 0 | 233 (24.1) |
| K219E | 13 (2.4) | 10 (2.5) | 0 | 23 (2.4) |
| NNRTI major | ||||
| K103N | 86 (15.6) | 109 (27.2) | 4 (28.6) | 199 (20.6) |
| G190S | 61 (11) | 0 | 0 | 61 (6.3) |
| Y181C | 27 (4.9) | 15 (3.7) | 0 | 42 (4.3) |
| P225H | 15 (2.7) | 15 (3.7) | 1 (7.1) | 31 (3.2) |
| G190A | 5 (0.9) | 8 (2) | 0 | 13 (1.3) |
| K238T | 3 (0.5) | 7 (1.7) | 0 | 10 (1) |
| NNRTI accessory | ||||
| K101E | 40 (7.2) | 4 (1) | 0 | 44 (4.5) |
| E138A | 28 (5.1) | 11 (2.7) | 1 (7.1) | 40 (4.1) |
| A98G | 11 (2) | 5 (1.2) | 0 | 16 (1.7) |
| V179E | 3 (0.5) | 11 (2.7) | 1 (7.1) | 15 (1.5) |
| V108I | 10 (1.8) | 4 (1) | 0 | 14 (1.4) |
| A6 | CRF02_AG | |||
|---|---|---|---|---|
| n (%) | n (%) | |||
| A62V | DRM present | GTT | 191 (86) | 0 |
| GTC | 9 (4.1) | 8 (72.7) | ||
| Other | 22 (9.9) | 3 (27.3) | ||
| DRM absent (χ2 test p-value < 2 × 10−16) | GCT | 318 (96.1) | 22 (5.6) | |
| GCC | 4 (1.2) | 347 (89) | ||
| Other | 9 (2.7) | 21 (5.4) | ||
| K103N | DRM present | AAC | 54 (62.8) | 65 (59.6) |
| Other | 32 (37.2) | 44 (40.4) | ||
| DRM absent | AAA | 438 (93.8) | 275 (94.2) | |
| Other | 29 (6.2) | 17 (5.8) | ||
| G190S | DRM present | AGC | 53 (86.9) | 0 |
| Other | 8 (13.1) | 0 | ||
| DRM absent (χ2 test p-value < 2 × 10−16) | GGC | 449 (91.3) | 10 (2.5) | |
| GGG | 3 (0.6) | 358 (89.3) | ||
| Other | 40 (8.1) | 33 (8.2) | ||
| K101E | DRM present | GAA | 36 (90) | 2 (50) |
| Other | 4 (10) | 2 (50) | ||
| DRM absent | AAA | 472 (92) | 369 (92.9) | |
| Other | 41 (8) | 28 (7.1) | ||
| D67N | DRM present | AAT | 7 (43.8) | 1 (100) |
| AAC | 6 (37.5) | 0 | ||
| Other | 3 (18.8) | 0 | ||
| DRM absent (χ2 test p-value < 2 × 10−16) | GAT | 265 (49.3) | 367 (91.8) | |
| GAC | 257 (47.9) | 17 (4.3) | ||
| Other | 15 (2.8) | 16 (4) | ||
| V179E | DRM present | GAA | 1 (33.3) | 9 (81.8) |
| GAG | 1 (33.3) | 0 | ||
| Other | 1 (33.3) | 2 (18.2) | ||
| DRM absent (χ2 test p-value < 2 × 10−16) | GTT | 413 (75.1) | 4 (1) | |
| GTA | 2 (0.4) | 295 (75.6) | ||
| Other | 135 (24.5) | 91 (23.3) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanaubarova, A.; Pujol-Hodge, E.; Dzissyuk, N.; Lemey, P.; Vermund, S.H.; Leigh Brown, A.J.; Ali, S. High-Level Drug-Resistant Mutations among HIV-1 Subtype A6 and CRF02_AG in Kazakhstan. Viruses 2023, 15, 1407. https://doi.org/10.3390/v15071407
Sanaubarova A, Pujol-Hodge E, Dzissyuk N, Lemey P, Vermund SH, Leigh Brown AJ, Ali S. High-Level Drug-Resistant Mutations among HIV-1 Subtype A6 and CRF02_AG in Kazakhstan. Viruses. 2023; 15(7):1407. https://doi.org/10.3390/v15071407
Chicago/Turabian StyleSanaubarova, Ainur, Emma Pujol-Hodge, Natalya Dzissyuk, Philippe Lemey, Sten H. Vermund, Andrew J. Leigh Brown, and Syed Ali. 2023. "High-Level Drug-Resistant Mutations among HIV-1 Subtype A6 and CRF02_AG in Kazakhstan" Viruses 15, no. 7: 1407. https://doi.org/10.3390/v15071407
APA StyleSanaubarova, A., Pujol-Hodge, E., Dzissyuk, N., Lemey, P., Vermund, S. H., Leigh Brown, A. J., & Ali, S. (2023). High-Level Drug-Resistant Mutations among HIV-1 Subtype A6 and CRF02_AG in Kazakhstan. Viruses, 15(7), 1407. https://doi.org/10.3390/v15071407