
Structure of Bovine CD46 Ectodomain

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning and Expression
2.2. Purification and Crystallisation
2.3. Data Collection, Structure Solution and Refinement
3. Results
3.1. Structure Determination
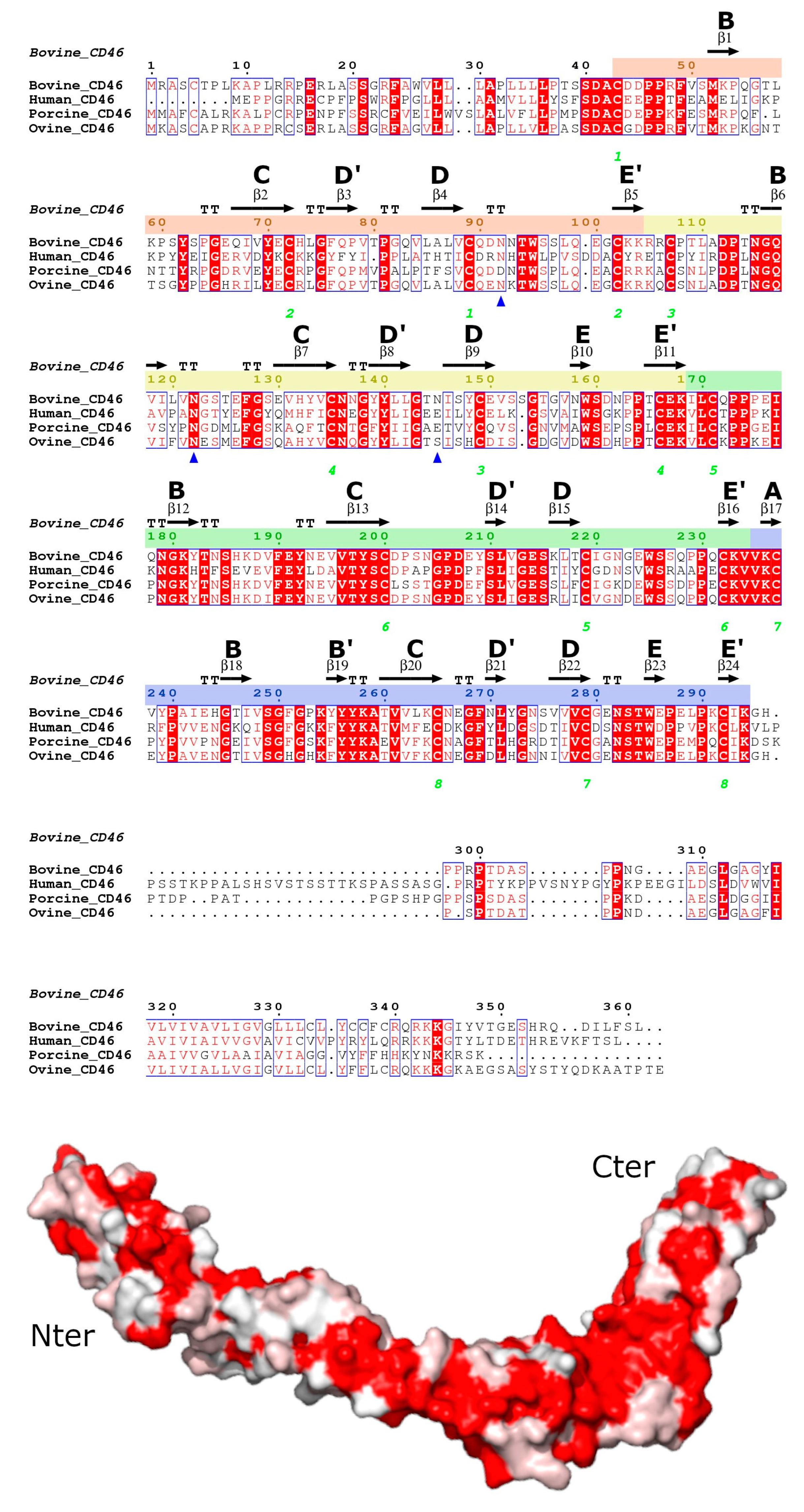
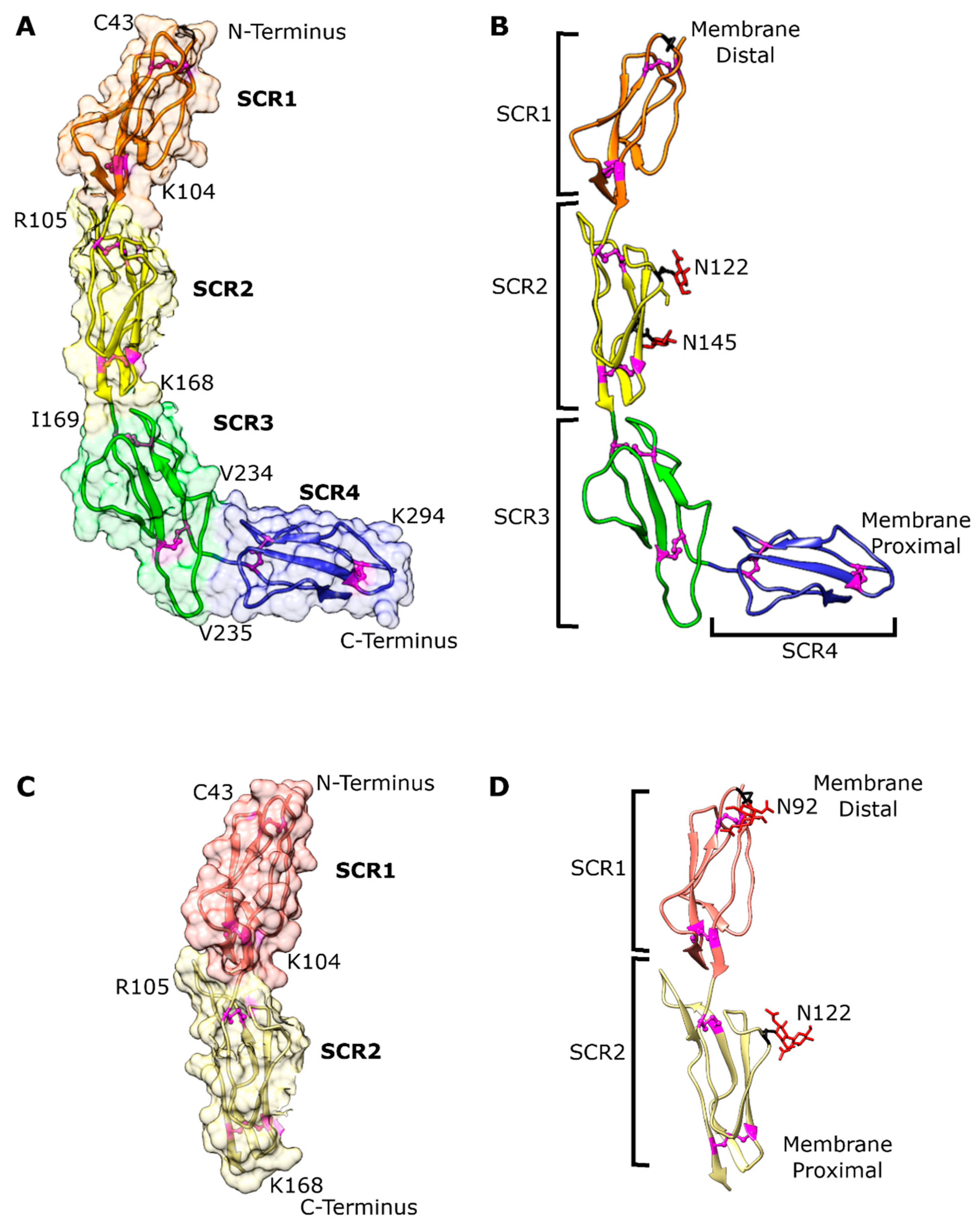
3.2. Overall bovCD46 Fold
3.3. Individual Domain Fold
3.4. Putative BVDV Binding Site
3.5. Crystallographic Dimer
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seya, T.; Turner, J.R.; Atkinson, J.P. Purification and characterization of a membrane protein (gp45-70) that is a cofactor for cleavage of C3b and C4b. J. Exp. Med. 1986, 163, 837–855. [Google Scholar] [CrossRef] [PubMed]
- Liszewski, M.K.; Post, T.W.; Atkinson, J.P. Membrane Cofactor Protein (MCP or CD46): Newest Member of the Regulators of Complement Activation Gene Cluster. Annu. Rev. Immunol. 1991, 9, 431–455. [Google Scholar] [CrossRef] [PubMed]
- Persson, B.D.; Schmitz, N.B.; Santiago, C.; Zocher, G.; Larvie, M.; Scheu, U.; Casasnovas, J.M.; Stehle, T. Structure of the Extracellular Portion of CD46 Provides Insights into Its Interactions with Complement Proteins and Pathogens. PLoS Pathog. 2010, 6, e1001122. [Google Scholar] [CrossRef] [Green Version]
- Arlaud, G.J.; Barlow, P.N.; Gaboriaud, C.; Gros, P.; Narayana, S.V.L. Deciphering complement mechanisms: The contributions of structural biology. Mol. Immunol. 2007, 44, 3809–3822. [Google Scholar] [CrossRef] [Green Version]
- Post, T.W.; Liszewski, M.K.; Adams, E.M.; Tedja, I.; Miller, E.A.; Atkinson, J.P. Membrane cofactor protein of the complement system: Alternative splicing of serine/threonine/proline-rich exons and cytoplasmic tails produces multiple isoforms that correlate with protein phenotype. J. Exp. Med. 1991, 174, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Liszewski, M.K.; Atkinson, J.P. Membrane cofactor protein (MCP; CD46): Deficiency states and pathogen connections. Curr. Opin. Immunol. 2021, 72, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Liszewski, M.K.; Atkinson, J.P. Complement regulator CD46: Genetic variants and disease associations. Hum. Genom. 2015, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Krey, T.; Himmelreich, A.; Heimann, M.; Menge, C.; Thiel, H.-J.; Maurer, K.; Rümenapf, T. Function of Bovine CD46 as a Cellular Receptor for Bovine Viral Diarrhea Virus Is Determined by Complement Control Protein 1. J. Virol. 2006, 80, 3912–3922. [Google Scholar] [CrossRef] [Green Version]
- Childs, T. X Disease of Cattle-Saskatchewan. Can. J. Comp. Med. Vet. Sci. 1946, 10, 316–319. [Google Scholar]
- Underdahl, N.R.; Grace, O.D.; Hoerlein, A.B. Cultivation in tissue-culture of cytopathogenic agent from bovine mucosal disease. Proc. Soc. Exp. Biol. Med. 1957, 94, 795–797. [Google Scholar] [CrossRef]
- Richter, V.; Lebl, K.; Baumgartner, W.; Obritzhauser, W.; Käsbohrer, A.; Pinior, B. A systematic worldwide review of the direct monetary losses in cattle due to bovine viral diarrhoea virus infection. Vet. J. 2017, 220, 80–87. [Google Scholar] [CrossRef]
- Gethmann, J.; Probst, C.; Bassett, J.; Blunk, P.; Hövel, P.; Conraths, F.J. An Epidemiological and Economic Simulation Model to Evaluate Strategies for the Control of Bovine Virus Diarrhea in Germany. Front. Vet. Sci. 2019, 6, 406. [Google Scholar] [CrossRef]
- Cagatay, G.N.; Antos, A.; Suckstorff, O.; Isken, O.; Tautz, N.; Becher, P.; Postel, A. Porcine Complement Regulatory Protein CD46 Is a Major Receptor for Atypical Porcine Pestivirus but Not for Classical Swine Fever Virus. J. Virol. 2021, 95, e02186-20. [Google Scholar] [CrossRef] [PubMed]
- Riedel, C.; Chen, H.-W.; Reichart, U.; Lamp, B.; Laketa, V.; Rümenapf, T. Real Time Analysis of Bovine Viral Diarrhea Virus (BVDV) Infection and Its Dependence on Bovine CD46. Viruses 2020, 12, 116. [Google Scholar] [CrossRef] [Green Version]
- Workman, A.M.; Heaton, M.P.; Vander Ley, B.L.; Webster, D.A.; Sherry, L.; Bostrom, J.R.; Larson, S.; Kalbfleisch, T.S.; Harhay, G.P.; Jobman, E.E.; et al. First gene-edited calf with reduced susceptibility to a major viral pathogen. PNAS Nexus 2023, 2, pgad125. [Google Scholar] [CrossRef] [PubMed]
- Casasnovas, J.M.; Larvie, M.; Stehle, T. Crystal structure of two CD46 domains reveals an extended measles virus-binding surface. EMBO J. 1999, 18, 2911–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Liu, B.; Du, J.; Zhang, J.; Lu, L.; Zhu, G.; Han, Y.; Su, H.; Yang, L.; Zhang, S.; et al. Discovery of Diverse Rodent and Bat Pestiviruses With Distinct Genomic and Phylogenetic Characteristics in Several Chinese Provinces. Front. Microbiol. 2018, 9, 2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Elegheert, J.; Behiels, E.; Bishop, B.; Scott, S.; Woolley, R.E.; Griffiths, S.C.; Byrne, E.F.X.; Chang, V.T.; Stuart, D.I.; Jones, E.Y.; et al. Lentiviral transduction of mammalian cells for fast, scalable and high-level production of soluble and membrane proteins. Nat. Protoc. 2018, 13, 2991–3017. [Google Scholar] [CrossRef]
- Brewster, A.S.; Waterman, D.G.; Parkhurst, J.M.; Gildea, R.J.; Young, I.D.; O’Riordan, L.J.; Yano, J.; Winter, G.; Evans, G.; Sauter, N.K. Improving signal strength in serial crystallography with DIALS geometry refinement. Acta Crystallogr. D 2018, 74, 877–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tickle, I.J.; Flensburg, C.; Keller, P.; Paciorek, W.; Sharff, A.; Vonrhein, C.; Bricogne, G. Staraniso; Global Phasing Ltd.: Cambridge, UK, 2018. [Google Scholar]
- Winter, G. xia2: An expert system for macromolecular crystallography data reduction. J. Appl. Crystallogr. 2010, 43, 186–190. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkóczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.W.; Jain, S.; McCoy, A.J.; et al. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. D 2019, 75, 861–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Santiago, C.; Celma, M.L.; Stehle, T.; Casasnovas, J.M. Structure of the measles virus hemagglutinin bound to the CD46 receptor. Nat. Struct. Mol. Biol. 2010, 17, 124–129. [Google Scholar] [CrossRef]
- Maurer, K.; Krey, T.; Moennig, V.; Thiel, H.-J.; Rümenapf, T. CD46 Is a Cellular Receptor for Bovine Viral Diarrhea Virus. J. Virol. 2004, 78, 1792–1799. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| bovCD46_SCR1-2 | bovCD46_SCR1-4 | |
|---|---|---|
| Wavelength (Å) | 0.98 | 0.98 |
| Resolution range (Å) | 44.12–2.33 (2.41–2.33) | 91.73–2.84 (3.24–2.84) |
| Space group | P 41 21 2 | P 41 21 2 |
| a, b, c (Å) | 91.09 91.09 121.12 | 129.73 129.73 120.19 |
| α, β, γ (°) | 90 90 90 | 90 90 90 |
| Total no. of reflections | 570,316 (55,868) | 278,989 (11,165) |
| No. of unique reflections | 22,417 (2168) | 10,884 (544) |
| Multiplicity | 25.40 (25.80) | 25.60 (20.50) |
| Completeness (%) | 99.40 (99.45) | Spherical: 44.1 (7.00) Ellipsoidal: 93.30 (73.30) |
| 〈I/σ(I)〉 | 10.19 (1.12) | 7.10 (1.80) |
| Overall B factor, Wilson plot (Å2) | 52.03 | 50.37 |
| Rmerge | 0.23 (3.60) | 0.49 (2.23) |
| Rmeas | 0.24 (3.67) | 0.50 (2.28) |
| Rpim | 0.05 (0.72) | 0.10 (0.47) |
| CC1/2 | 1.00 (0.74) | 1.00 (0.63) |
| Final Rwork | 0.23 (0.35) | 0.25 (0.39) |
| Final Rfree | 0.26 (0.38) | 0.29 (0.28) |
| RMSD bonds (Å)/angles (°) | 0.01/0.90 | 0.02/1.84 |
| Average B factors (Å2) | 79.1 | 87.5 |
| Ramachandran: favoured/allowed/outliers (%) | 97.64/2.36/0.00 | 95.50/4.50/0.00 |
| Rotamer outliers (%) | 1.77 | 0.25 |
| Clashscore | 2.49 | 14.55 |
| Number of TLS groups | 4 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aitkenhead, H.; Stuart, D.I.; El Omari, K. Structure of Bovine CD46 Ectodomain. Viruses 2023, 15, 1424. https://doi.org/10.3390/v15071424
Aitkenhead H, Stuart DI, El Omari K. Structure of Bovine CD46 Ectodomain. Viruses. 2023; 15(7):1424. https://doi.org/10.3390/v15071424
Chicago/Turabian StyleAitkenhead, Hazel, David I. Stuart, and Kamel El Omari. 2023. "Structure of Bovine CD46 Ectodomain" Viruses 15, no. 7: 1424. https://doi.org/10.3390/v15071424