
Compounding Achromobacter Phages for Therapeutic Applications
 , , , , , , , , , add
Show full author list
, , , , , , , , , add
Show full author list
Abstract
:1. Introduction
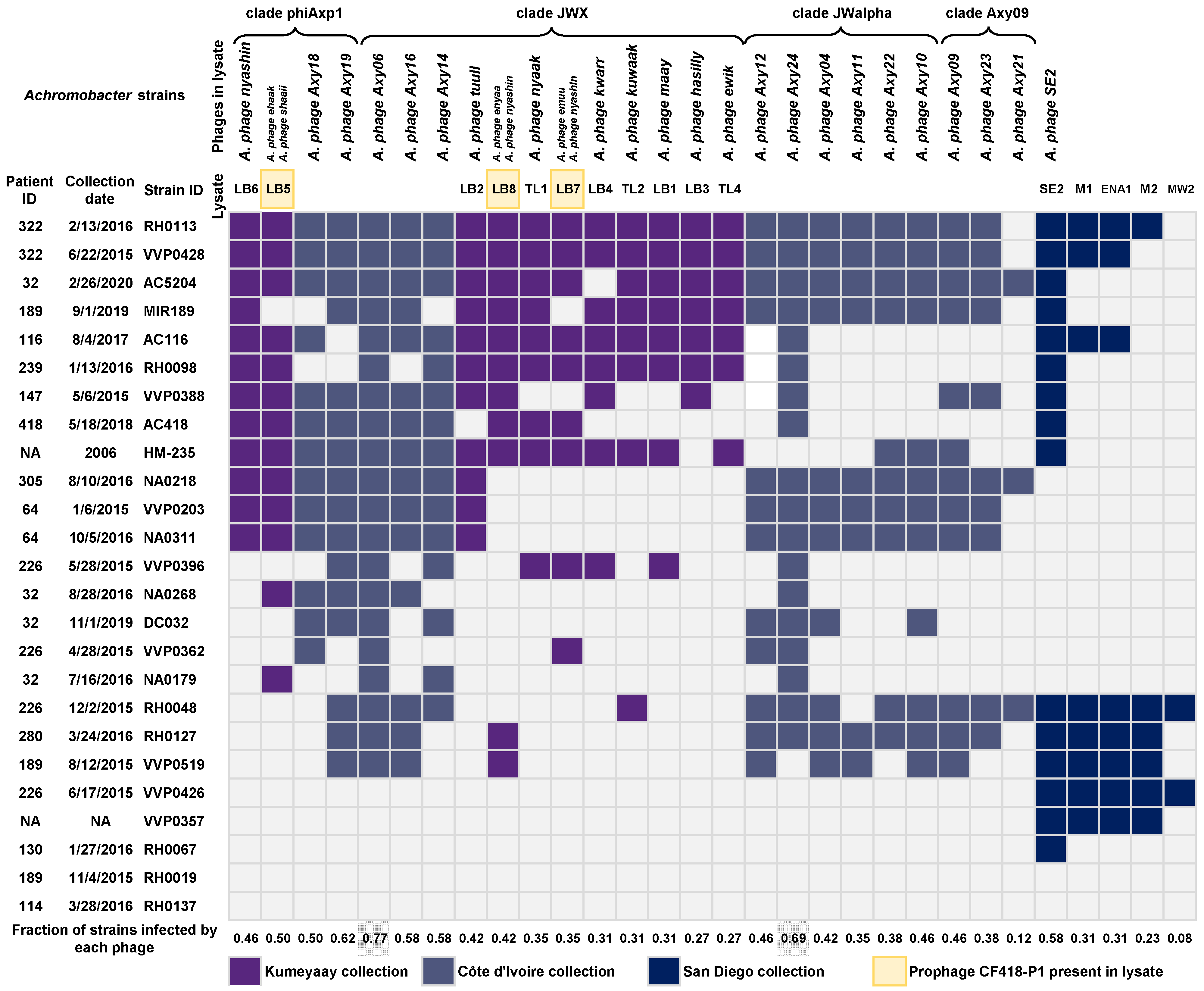
2. Results
2.1. Cystic Fibrosis and Achromobacter
2.2. Achromobacter Clinical Isolates
2.3. Isolation and Characterization of Achromophages
2.4. Comparative Genomics of Achromobacter Phages
2.5. Achromophage Genome Annotation
2.6. Annotation of Toxin-Coding and Antibiotic Resistance Coding ORFs in Achromophages
2.7. Achromophages Lifestyle Determination
2.8. Prophage CF418-P1 Induction and Characterization
2.9. Host Range Determination for Achromophages
2.10. Generalized Transduction Potential of the maay (LB1) and nyashin (LB6) Phages
2.11. Preparation of Achromobacter Phages Lysates for Therapeutic Applications
2.12. Evaluation of Phages Cytotoxicity in Lung Epithelial Cells
3. Discussion
3.1. Hunting for Achromophages
3.2. Induction of Cryptic Prophages by Lytic Phage Replication
3.3. Endotoxin Removal from Phage Lysates for Therapeutic Applications
3.4. Identification of Toxin and Antibiotic Resistance Genes in Phage Genomes
3.5. Evaluation of Phage Cytotoxicity in Lung Epithelial Cells
3.6. Beyond Phage Hunting
4. Materials and Methods
4.1. Cystic Fibrosis Metagenomes
4.2. Achromobacter Strains
4.3. Phage Hunting
4.4. Transmission Electron Microscopy (TEM) of Phages
4.5. Determination of Phage Genome Size by Pulsed Field Gel Electrophoresis (PFGE)
4.6. Determination of Phage Host Range
4.7. Isolation of Phage DNA for Sequencing
4.8. Illumina Sequencing of Phages
4.9. Nanopore Sequencing of Phages
4.10. Assembly of Phage Genomes
4.11. Annotation of Phage Genomes
4.12. Comparative Genomics of Achromophages
4.13. Testing Potential for Generalized Transduction of Phages Maay and Nyashin
4.14. High-Titer Phage Production and Endotoxin Removal and Quantification
4.15. Evaluation of Phage Cytotoxicity in Lung Epithelial Cells
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ridderberg, W.; Nielsen, S.M.; Nørskov-Lauritsen, N. Genetic Adaptation of Achromobacter sp. during Persistence in the Lungs of Cystic Fibrosis Patients. PLoS ONE 2015, 10, e0136790. [Google Scholar] [CrossRef]
- Jakobsen, T.H.; Hansen, M.A.; Jensen, P.Ø.; Hansen, L.; Riber, L.; Kolpen, M.; Hansen, C.R.; Rønne Hansen, C.; Ridderberg, W.; Eickhardt, S.; et al. Complete Genome Sequence of the Cystic Fibrosis Pathogen Achromobacter xylosoxidans NH44784-1996 Complies with Important Pathogenic Phenotypes. PLoS ONE 2013, 8, e68484. [Google Scholar] [CrossRef] [PubMed]
- Tena, D.; González-Praetorius, A.; Pérez-Balsalobre, M.; Sancho, O.; Bisquert, J. Urinary Tract Infection Due to Achromobacter xylosoxidans: Report of 9 Cases. Scand. J. Infect. Dis. 2008, 40, 84–87. [Google Scholar] [CrossRef]
- Tokuyasu, H.; Fukushima, T.; Nakazaki, H.; Shimizu, E. Infective Endocarditis Caused by Achromobacter xylosoxidans: A Case Report and Review of the Literature. Intern. Med. 2012, 51, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Manckoundia, P.; Mazen, E.; Coste, A.S.; Somana, S.; Marilier, S.; Duez, J.M.; Camus, A.; Popitean, L.; Bador, J.; Pfitzenmeyer, P. A Case of Meningitis Due to Achromobacter xylosoxidans Denitrificans 60 Years after a Cranial Trauma. Med. Sci. Monit. 2011, 17, 2010–2012. [Google Scholar] [CrossRef]
- Park, J.H.; Song, N.H.; Koh, J.W. Achromobacter xylosoxidans Keratitis after Contact Lens Usage. Korean J. Ophthalmol. 2012, 26, 49–53. [Google Scholar] [CrossRef]
- Gordillo-Altamirano, F.L.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage Therapy: A Renewed Approach to Combat Antibiotic-Resistant Bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef]
- Dedrick, R.M.; Guerrero-bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered Bacteriophages for Treatment of a Patient with a Disseminated Drug-Resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef]
- Law, N.; Logan, C.; Yung, G.; Furr, C.L.L.; Lehman, S.M.; Morales, S.; Rosas, F.; Gaidamaka, A.; Bilinsky, I.; Grint, P.; et al. Successful Adjunctive Use of Bacteriophage Therapy for Treatment of Multidrug-Resistant Pseudomonas aeruginosa Infection in a Cystic Fibrosis Patient. Infection 2019, 47, 665–668. [Google Scholar] [CrossRef]
- Hoyle, N.; Zhvaniya, P.; Balarjishvili, N.; Bolkvadze, D.; Nadareishvili, L.; Nizharadze, D.; Wittmann, J.; Rohde, C.; Kutateladze, M. Phage Therapy against Achromobacter xylosoxidans Lung Infection in a Patient with Cystic Fibrosis: A Case Report. Res. Microbiol. 2018, 169, 540–542. [Google Scholar] [CrossRef] [PubMed]
- Kutateladze, M.; Adamia, R. Phage Therapy Experience at the Eliava Institute. Med. Mal. Infect. 2008, 38, 426–430. [Google Scholar] [CrossRef]
- Dreiseikelmann, B.; Bunk, B.; Spröer, C.; Rohde, M.; Nimtz, M.; Wittmann, J. Characterization and Genome Comparisons of Three Achromobacter Phages of the Family Siphoviridae. Arch. Virol. 2017, 162, 2191–2201. [Google Scholar] [CrossRef]
- Wittmann, J.; Dreiseikelmann, B.; Rohde, M.; Meier-Kolthoff, J.P.; Bunk, B.; Rohde, C. First Genome Sequences of Achromobacter Phages Reveal New Members of the N4 Family. Virol. J. 2014, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, J.; Dreiseikelmann, B.; Rohde, C.; Rohde, M.; Sikorski, J. Isolation and Characterization of Numerous Novel Phages Targeting Diverse Strains of the Ubiquitous and Opportunistic Pathogen Achromobacter xylosoxidans. PLoS ONE 2014, 9, e86935. [Google Scholar] [CrossRef]
- Essoh, C.; Vernadet, J.P.; Vergnaud, G.; Coulibaly, A.; Kakou-N’Douba, A.; N’Guetta, A.S.P.; Ouassa, T.; Pourcel, C. Characterization of Sixteen Achromobacter xylosoxidans Phages from Abidjan, Côte d’Ivoire, Isolated on a Single Clinical Strain. Arch. Virol. 2020, 165, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Bradley, J.S.; Hajama, H.; Akong, K.; Jordan, M.; Stout, D.; Rowe, R.S.; Conrad, D.J.; Hingtgen, S.; Segall, A.M. Bacteriophage Therapy of Multidrug-Resistant Achromobacter in an 11-year-old Boy with Cystic Fibrosis Assessed by Metagenome Analysis. Pediatr. Infect. Dis. J. 2023, in press. [Google Scholar] [CrossRef]
- BEI. The Following Reagent Was Obtained through BEI Resources, NIAID, NIH as Part of the Human Microbiome Project: Achromobacter xylosoxidans, Strain C54, HM-235. Available online: https://www.beiresources.org/Catalog/bacteria/HM-235.aspx (accessed on 1 December 2019).
- Leibniz Institute. DSMZ. Available online: https://www.dsmz.de/ (accessed on 1 December 2019).
- McNair, K.; Zhou, C.; Dinsdale, E.A.; Souza, B.; Edwards, R.A. PHANOTATE: A Novel Approach to Gene Identification in Phage Genomes. Bioinformatics 2019, 35, 4537–4542. [Google Scholar] [CrossRef]
- Rohwer, F.; Edwards, R. The Phage Proteomic Tree: A Genome-Based Taxonomy for Phage. J. Bacteriol. 2002, 184, 4529–4535. [Google Scholar] [CrossRef]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. Genome Analysis ViPTree: The Viral Proteomic Tree Server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A Modular and Extensible Implementation of the RAST Algorithm for Building Custom Annotation Pipelines and Annotating Batches of Genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The Conserved Domain Database in 2020. Nucleic Acids Res. 2020, 48, D265–D2668. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Bryant, S.H. CD-Search: Protein Domain Annotations on the Fly. Nucleic Acids Res. 2004, 32, 327–331. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the Functional Annotation of Proteins. Nucleic Acids Res. 2011, 39, 225–229. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER Web Server: 2018 Update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Cantú, V.A.; Salamon, P.; Seguritan, V.; Redfield, J.; Edwards, R.A.; Segall, A.M. PhANNs, a fast and accurate tool and web server to predict and classify phage structural proteins. PLoS Comput. Biol. 2020, 16, e1007845. [Google Scholar] [CrossRef]
- Rao, V.B.; Feiss, M. Mechanisms of DNA Packaging by Large Double-Stranded DNA Viruses. Annu. Rev. Virol. 2015, 2, 351–378. [Google Scholar] [CrossRef]
- Leplae, R.; Hebrant, A.; Wodak, S.J.; Toussaint, A. ACLAME: A Classification of Mobile Genetic Elements. Nucleic Acids Res. 2004, 32, 45–49. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and Refined Dataset for Big Data Analysis—10 Years On. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- Sayers, S.; Li, L.; Ong, E.; Deng, S.; Fu, G.; Lin, Y.; Yang, B.; Zhang, S.; Fa, Z.; Zhao, B.; et al. Victors: A Web-Based Knowledge Base of Virulence Factors in Human and Animal Pathogens. Nucleic Acids Res. 2019, 47, D693–D700. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the All-Bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- McNair, K.; Bailey, B.A.; Edwards, R.A. PHACTS, a Computational Approach to Classifying the Lifestyle of Phages. Bioinformatics 2012, 28, 614–618. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Saied, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucl Acids Res 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Huang, H.; Masters, M. Bacteriophage P1 pac sites inserted into the chromosome greatly increase packaging and transduction of Escherichia coli genomic DNA. Virology 2014, 468–470, 274–282. [Google Scholar] [CrossRef]
- Penadés, J.R.; Chen, J.; Quiles-Puchalt, N.; Carpena, N.; Novick, R.P. Bacteriophage-mediated spread of bacterial virulence genes. Curr. Opin. Microbiol. 2015, 23, 171–178. [Google Scholar] [CrossRef]
- Young, K.K.; Edlin, G. Physical and Genetical Analysis of Bacteriophage T4 Generalized Transduction. Mol. Gen. Genet. 1983, 192, 241–246. [Google Scholar] [CrossRef]
- Young, K.K.; Edlin, G.J.; Wilson, G.G. Genetic Analysis of Bacteriophage T4 transducing Bacteriophages. J. Virol. 1982, 41, 345–347. [Google Scholar] [CrossRef]
- Lee, A.K.; Falkow, S. Constitutive and Inducible Green Fluorescent Protein Expression in Bartonella henselae. Infect. Immun. 1998, 66, 3964–3967. [Google Scholar] [CrossRef]
- Rowe, R.S. Identifying Host Factors for Bacteriophage Replication in Achromobacter species. Masters’ Thesis, San Diego State University, San Diego, CA, USA, 2023. [Google Scholar]
- Rowe, R.S.; Hajama, H.; Jacobson, N.E.; Segall, A.M. Identifying Host Factors Involved in Clade JWX Bacteriophage Replication in Achromobacter species. Department of Biology and Viral Information Institute, San Diego State University, San Diego, CA, USA. 2023; manuscript in preparation. [Google Scholar]
- Orchard, S.S.; Rostron, J.E.; Segall, A.M. Escherichia coli enterobactin synthesis and uptake mutants are hypersensitive to an antimicrobial peptide that limits the availability of iron in addition to blocking Holliday junction resolution. Microbiology 2012, 158, 547–559. [Google Scholar] [CrossRef]
- Wagner, P.L.; Neely, M.N.; Acheson, D.W.K.; Friedman, D.I.; Waldor, M.K. Bacteriophage Control of Shiga Toxin 1 Production and Release by Escherichia coli. Mol. Microbiol. 2002, 44, 957–970. [Google Scholar] [CrossRef]
- Li, E.; Yin, Z.; Ma, Y.; Li, H.; Lin, W.; Wei, X.; Zhao, R. Identification and Molecular Characterization of Bacteriophage PhiAxp-2 of Achromobacter xylosoxidans. Sci. Rep. 2016, 6, 34300. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Zhao, J.; Ma, Y.; Wei, X.; Li, H.; Lin, W.; Wang, X.; Li, C.; Shen, Z.; Zhao, R.; et al. Characterization of a Novel Achromobacter xylosoxidans Specific Siphoviruse: PhiAxp-1. Sci. Rep. 2016, 6, 21943. [Google Scholar] [CrossRef]
- Ma, Y.; Li, E.; Qi, Z.; Li, H.; Wei, X.; Lin, W.; Zhao, R.; Jiang, A.; Yang, H.; Yin, Z.; et al. Isolation and Molecular Characterisation of Achromobacter Phage PhiAxp-3, an N4-like Bacteriophage. Sci. Rep. 2016, 6, 24776. [Google Scholar] [CrossRef]
- Tal, N.; Sorek, R. SnapShot: Bacterial Immunity. Cell 2022, 185, 578–578.e1. [Google Scholar] [CrossRef]
- Woods, D.R.; Thomson, J.A. Unstable Generalized Transduction in Achromobacter. J. Gen. Microbiol. 1975, 88, 86–92. [Google Scholar] [CrossRef]
- Thomson, J.A.; Woods, D.R. Bacteriophages and Cryptic Lysogeny in Achromobacter. J. Gen. Virol. 1974, 22, 153–157. [Google Scholar]
- Penadés, J.R.; Christie, G.E. The Phage-Inducible Chromosomal Islands: A Family of Highly Evolved Molecular Parasites. Annu. Rev. Virol. 2015, 2, 181–201. [Google Scholar] [CrossRef]
- Fillol-Salom, A.; Martínez-Rubio, R.; Abdulrahman, R.F.; Chen, J.; Davies, R.; Penadés, J.R. Phage-Inducible Chromosomal Islands Are Ubiquitous within the Bacterial Universe. ISME J. 2018, 12, 2114–2128. [Google Scholar] [CrossRef]
- Pourcel, C.; Midoux, C.; Hauck, Y.; Vergnaud, G.; Latino, L. Large Preferred Region for Packaging of Bacterial DNA by phiC725A, a Novel Pseudomonas aeruginosa F116-Like Bacteriophage. PLoS ONE 2017, 12, e0169684. [Google Scholar] [CrossRef]
- Malik, D.J.; Sokolov, I.J.; Vinner, G.K.; Mancuso, F.; Cinquerrui, S.; Vladisavljevic, G.T.; Clokie, M.R.J.; Garton, N.J.; Stapley, A.G.F.; Kirpichnikova, A. Formulation, Stabilisation and Encapsulation of Bacteriophage for Phage Therapy. Adv. Colloid Interface Sci. 2017, 249, 100–133. [Google Scholar] [CrossRef]
- Danielsson, O.; Atrian, S.; Luque, T.; Hjelmqvist, L.; Gonzalez-Duarte, R.; Jornvall, H. Fundamental Molecular Differences Between Alcohol Dehydrogenase Classes. Proc. Natl. Acad. Sci. USA 1994, 91, 4980–4984. [Google Scholar] [PubMed]
- Jelski, W.; Chrostek, Ã.L.; Markiewicz, W.; Szmitkowski, M. Activity of Alcohol Dehydrogenase (ADH) Isoenzymes and Aldehyde Dehydrogenase (ALDH) in the Sera of Patients With Breast Cancer. Cancer 2006, 108, 105–108. [Google Scholar] [CrossRef]
- Satre, M.A.; Žgombić-Knight, M.; Duester, G. The Complete Structure of Human Class IV Alcohol Dehydrogenase (Retinol Dehydrogenase) Determined from the ADH7 Gene. J. Biol. Chem. 1994, 269, 15606–15612. [Google Scholar] [PubMed]
- McCord, J.P.; Muddiman, D.C.; Khaledi, M.G. Perfluorinated Alcohol Induced Coacervates as Extraction Media for Proteomic Analysis. J. Chromatogr. A 2017, 1523, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Philipson, C.W.; Voegtly, L.J.; Lueder, M.R.; Long, K.A.; Rice, G.K.; Frey, K.G.; Biswas, B.; Cer, R.Z.; Hamilton, T.; Bishop-Lilly, K.A. Characterizing Phage Genomes for Therapeutic Applications. Viruses 2018, 10, 188. [Google Scholar] [CrossRef]
- Porayath, C.; Salim, A.; Veedu, A.P.; Babu, P.; Nair, B.; Madhavan, A.; Pal, S. Characterization of the Bacteriophages Binding to Human Matrix Molecules. Int. J. Biol. Macromol. 2018, 110, 608–615. [Google Scholar] [CrossRef]
- Shan, J.; Ramachandran, A.; Thanki, A.M.; Vukusic, F.B.; Barylski, J.; Clokie, M.R. Bacteriophages Are More Virulent to Bacteria with Human Cells than They Are in Bacterial Culture; Insights from HT-29 Cells. Sci. Rep. 2018, 8, 5091. [Google Scholar] [CrossRef]
- Lemire, S.; Yehl, K.M.; Lu, T.K. Phage-Based Applications in Synthetic Biology. Annu. Rev. Virol. 2018, 5, 453–476. [Google Scholar] [CrossRef]
- Scholl, D. Phage Tail—Like Bacteriocins. Annu. Rev. Virol. 2017, 4, 453–467. [Google Scholar]
- Young, R.; Gill, J.J. Phage Therapy Redux—What Is to Be Done? Science 2015, 350, 1163–1164. [Google Scholar] [CrossRef]
- Chan, B.K.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage Selection Restores Antibiotic Sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef] [PubMed]
- Segall, A.M.; Roach, D.R.; Strathdee, S.A. Stronger Together? Perspectives on Phage-Antibiotic Synergy in Clinical Applications of Phage Therapy. Curr. Opin. Microbiol. 2019, 51, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.A.; Hatfull, G.F. PhagesDB: The Actinobacteriophage Database. Bioinformatics 2017, 33, 784–786. [Google Scholar] [CrossRef] [PubMed]
- Seguritan, V.S.; Alves, N., Jr.; Arnoult, M.; Raymond, A.; Lorimer, D.; Burgin, A.B., Jr.; Salamon, P.; Segall, A.M. Artificial neural networks trained to detect viral and phage structural proteins. PLoS Comput. Biol. 2012, e10002657. [Google Scholar] [CrossRef]
- Gill, J. Phage Genomic DNA Extraction. Available online: https://openwetware.org/wiki/Gill:Phage_genomic_DNA_extraction (accessed on 26 July 2023).
- Cantú, A.; Sadural, J.; Edwards, R. PRINSEQ++, a Multi-Threaded Tool for Fast and Efficient Quality Control and Preprocessing of Sequencing Datasets. PeerJ Prepr. 2019, 7, e27553v1. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Genome Analysis Bandage: Interactive Visualization of de Novo Genome Assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef]
- McNair, K. Circulaline. GitHub. 2019. Available online: https://github.com/deprekate/circulaline (accessed on 1 March 2021).
- EBI. HMMER Search. Available online: https://www.ebi.ac.uk/Tools/hmmer/search/hmmscan (accessed on 26 July 2023).
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A Genome Comparison Visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Alcock, B.P.; Huynh, W.; Chalil, R.; Smith, K.W.; Raphenya, A.R.; Wlodarski, M.A.; Edalatmand, A.; Petkau, A.; Syed, S.A.; Tsang, K.K.; et al. CARD 2023: Expanded curation, support for machine learning, and resistome prediction at the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2023, 51, D690–D699. [Google Scholar] [CrossRef]
- Wu, Q.; Pei, J.; Turse, C.; Ficht, T.A. Mariner mutagenesis of Brucella melitensis reveals genes with previously uncharacterized roles in virulence and survival. BMC Microbiol. 2006, 6, 102. [Google Scholar] [CrossRef]
- Perry, B.J.; Yost, C.K. Construction of a Mariner-Based Transposon Vector for Use inInsertion Sequence Mutagenesis in Selected Members of the Rhizobiaceae. BMC Microbiol. 2014, 14, 298. [Google Scholar] [CrossRef]
- Morrison, D.; Leive, L. Fractions of Lipopolysaccharide from Escherichia coli O111:B4 Prepared by Two Extraction Procedures. J. Biol. Chem. 1976, 2750, 2911–2919. [Google Scholar]
- Szermer-Olearnik, B.; Boratyński, J. Removal of Endotoxins from Bacteriophage Preparations by Extraction with Organic Solvents. PLoS ONE 2015, 10, e0122672. [Google Scholar] [CrossRef]
- Bonilla, N.; Rojas, M.I.; Cruz, G.N.F.; Hung, S.-H.; Rohwer, F.; Barr, J.J. Phage on Tap–A Quick and Efficient Protocol for the Preparation of Bacteriophage Laboratory Stocks. PeerJ 2016, 4, e2261. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage Name (GenBank Accession Number) | Genome Length (bp) | GC Content | CDS | tRNA | Repeat Regions | Clade | Host | Phage Name Etymology |
|---|---|---|---|---|---|---|---|---|
| nyashin (OQ817840) | 45,982 | 56.31 | 68 | 0 | 2 | phiAxp1 | A.ruhlandii CF418 | nyashin–today |
| shaaii (OQ817841) | 45,029 | 56.11 | 63 | 0 | 0 | phiAxp1 | A.ruhlandii CF418 | shaaii–buzzard |
| nyaak ( OQ817839) | 46,478 | 55.77 | 66 | tRNA-Pro-TGG | 2 | JWX | A. ruhlandii CF116 | nyaak–east |
| kuwaak (OQ817836) | 46,215 | 56.19 | 66 | tRNA-Pro-TGG | 0 | JWX | A. ruhlandii CF116 | kuwaak–south |
| ewik (OQ817842) | 50,543 | 55.75 | 83 | tRNA-Pro-TGG | 37 | JWX | A. ruhlandii CF116 | ewik–west |
| tuull (OR396896) | 47,460 | 55.79 | 92 | tRNA-Pro-TGG | 15 | JWX | A. ruhlandii CF116 | tuull–north |
| maay (OQ817838) | 46,086 | 56.31 | 62 | tRNA-Pro-TGG | 2 | JWX | A. ruhlandii CF116 | maay–sky |
| hasilly (OQ817843) | 46,478 | 55.77 | 65 | tRNA-Pro-TGG | 2 | JWX | A. ruhlandii CF116 | hasilly–sea |
| ehaak (OQ817833) | 46,435 | 56.17 | 64 | tRNA-Pro-TGG | 0 | JWX | A.ruhlandii CF418 | ehaak–raven |
| emuu (OQ817834) | 46,012 | 55.86 | 62 | tRNA-Pro-TGG | 2 | JWX | A.ruhlandii CF418 | emuu–mountain sheep |
| enyaa (OQ817835) | 43,305 | 55.51 | 64 | tRNA-Pro-TGG | 0 | JWX | A.ruhlandii CF418 | enyaa–sun |
| kwarr (OQ817837) | 33,215 | 55.59 | 73 | 0 | 6 | JWX | A. ruhlandii CF116 | kwarr–red earth to paint the body |
| SE2 (OQ817844) | 45,648 | 55.88 | 55 | tRNA-Pro-TGG | 0 | JWX | Achromobacter sp. VPP0426 | |
| prophage CF418-P1 (OQ817832) | 58,030 | 65.63 | 73 | tRNA-Val-AAC | 2 | A.ruhlandii CF418 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cobián Güemes, A.G.; Le, T.; Rojas, M.I.; Jacobson, N.E.; Villela, H.; McNair, K.; Hung, S.-H.; Han, L.; Boling, L.; Octavio, J.C.; et al. Compounding Achromobacter Phages for Therapeutic Applications. Viruses 2023, 15, 1665. https://doi.org/10.3390/v15081665
Cobián Güemes AG, Le T, Rojas MI, Jacobson NE, Villela H, McNair K, Hung S-H, Han L, Boling L, Octavio JC, et al. Compounding Achromobacter Phages for Therapeutic Applications. Viruses. 2023; 15(8):1665. https://doi.org/10.3390/v15081665
Chicago/Turabian StyleCobián Güemes, Ana Georgina, Tram Le, Maria Isabel Rojas, Nicole E. Jacobson, Helena Villela, Katelyn McNair, Shr-Hau Hung, Lili Han, Lance Boling, Jessica Claire Octavio, and et al. 2023. "Compounding Achromobacter Phages for Therapeutic Applications" Viruses 15, no. 8: 1665. https://doi.org/10.3390/v15081665