
Detection of Alpha- and Betacoronaviruses in Small Mammals in Western Yunnan Province, China
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
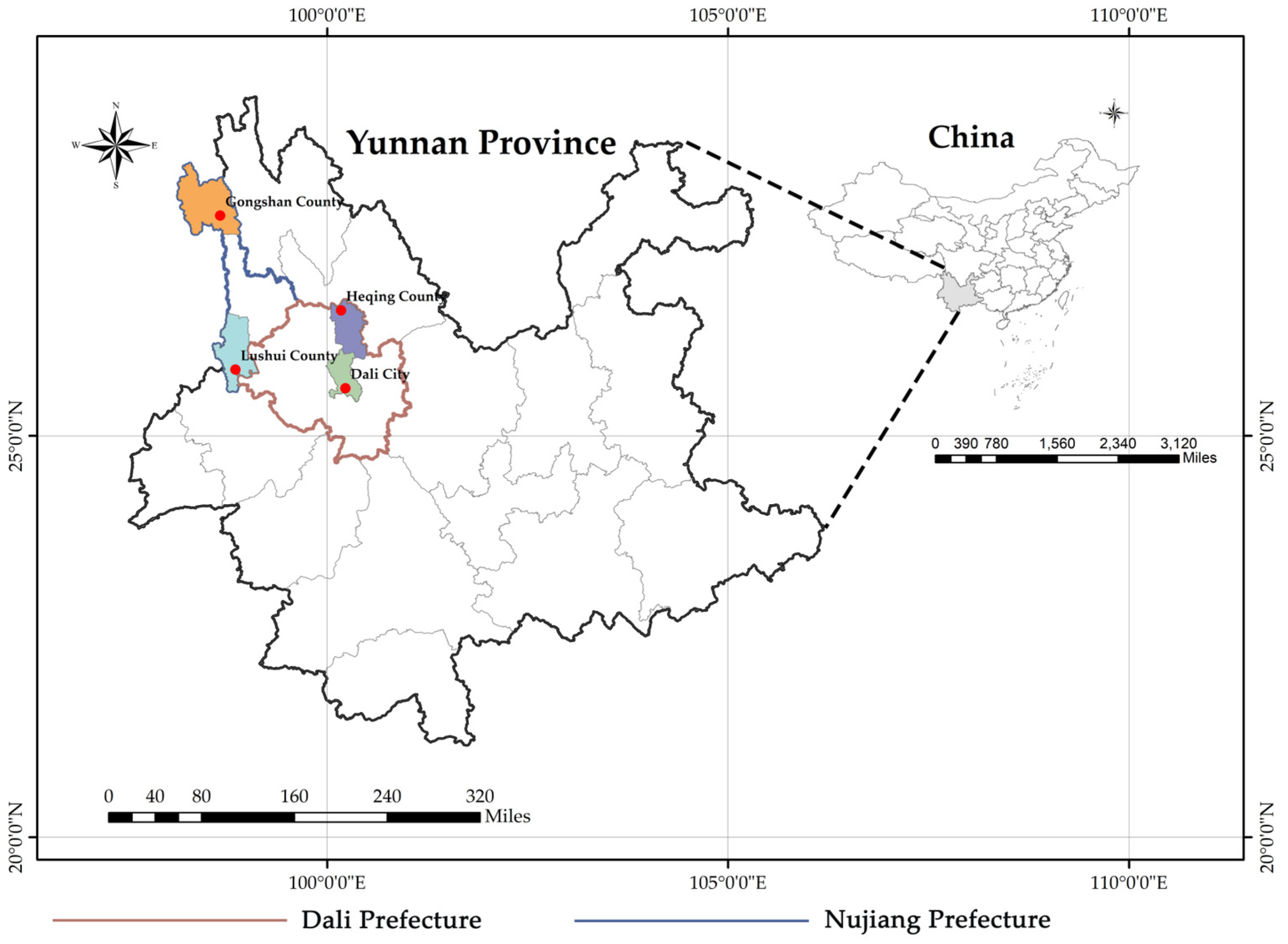
2.2. Sample Collection and Processing
2.3. DNA and RNA Extraction
2.4. Primary Screening of CoV and Amplification of Partial RdRp Fragments
2.5. Virus Sequence Identification and Phylogenetic Analysis
2.6. Construction of Plasmids and Determination of Virus Copies
2.7. Primer and TaqMan Probe Design and Optimization
2.8. Establishment of Standard Curves
2.9. Evaluation of qRT-PCR Methods
2.9.1. Sensitivity
2.9.2. Specificity
2.9.3. Repeatability and Stability
2.9.4. Comparison of qRT-PCR and RT-PCR Assays
2.10. qRT-PCR for Small Mammalian Samples
2.11. Tissue Tropism of Small Mammalian CoV
2.12. Statistical Analysis
3. Results
3.1. Collection of Samples and Detection of CoV
3.2. Comparison of Partial RdRp Gene
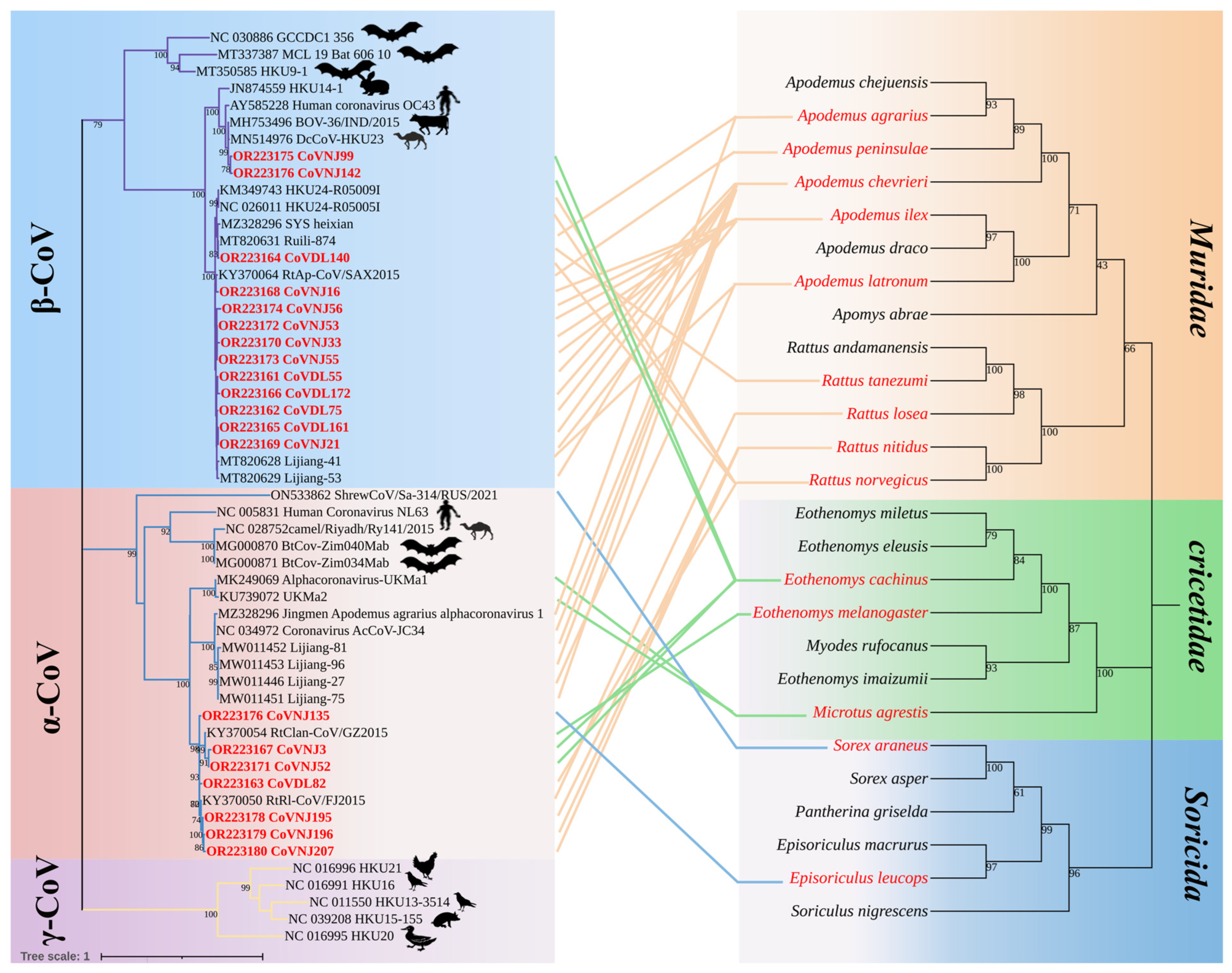
3.3. Phylogenetic Analysis
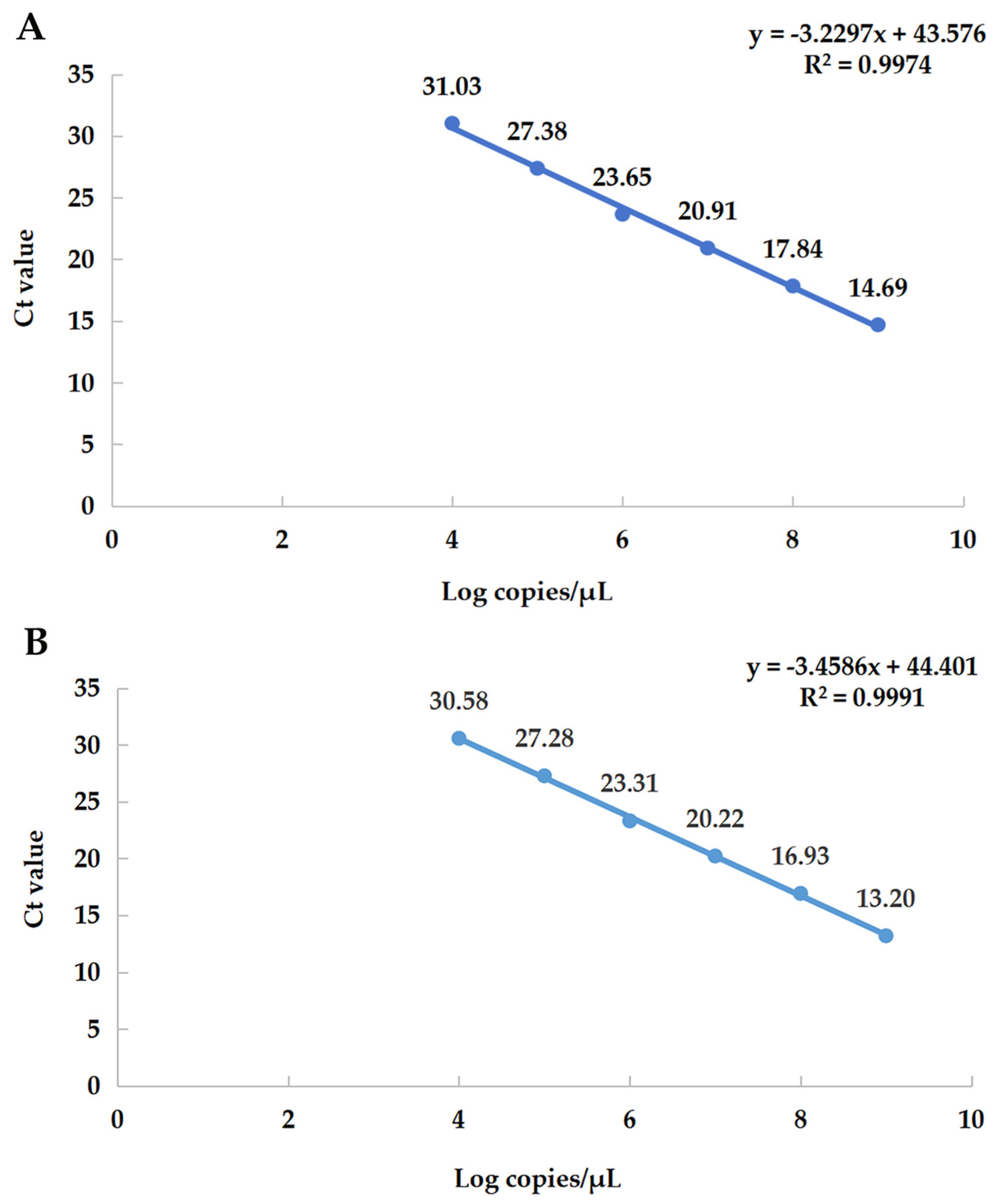
3.4. Establishment of qRT-PCR Standard Curves
3.5. Evaluation of qRT-PCR Methods
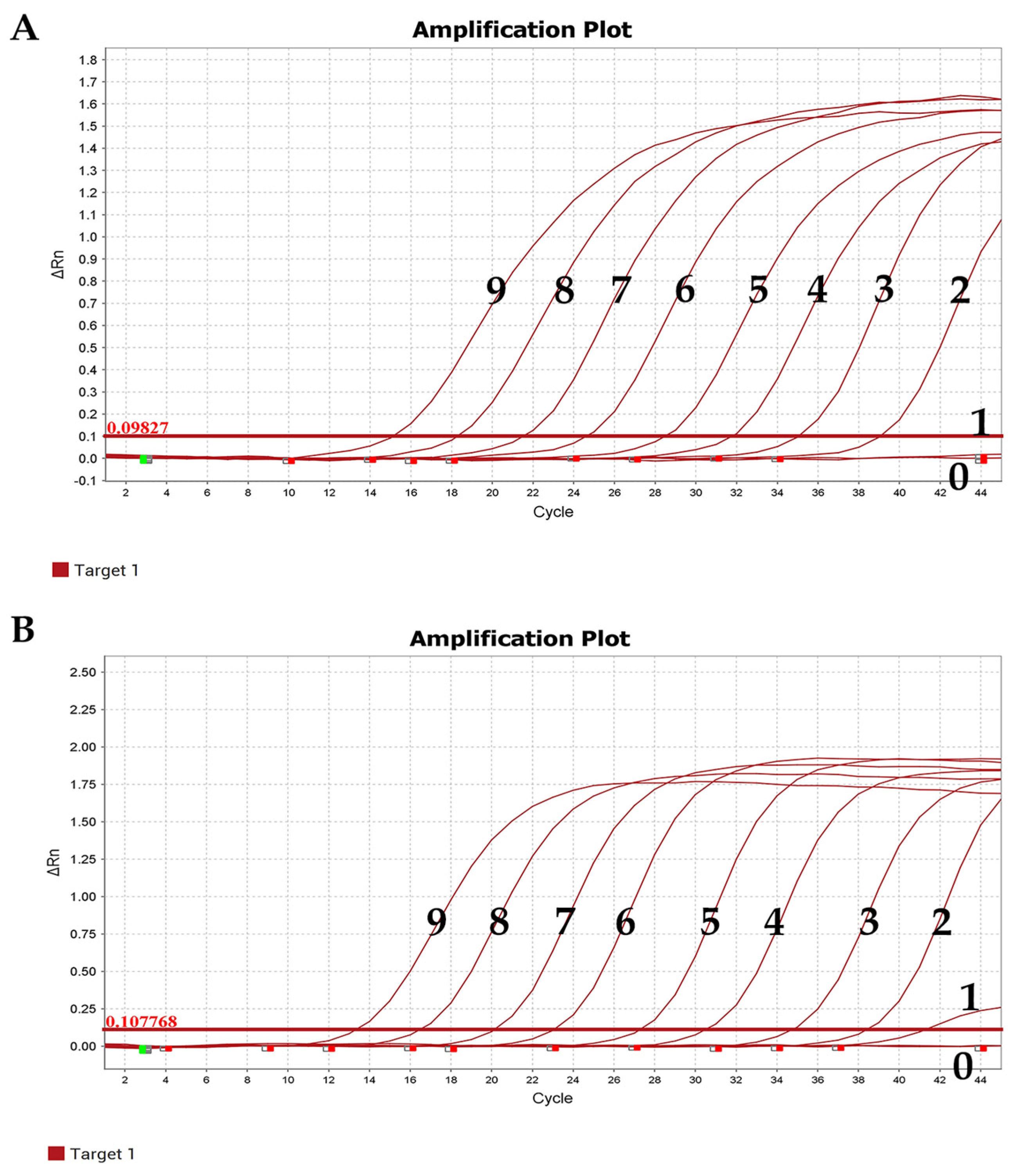
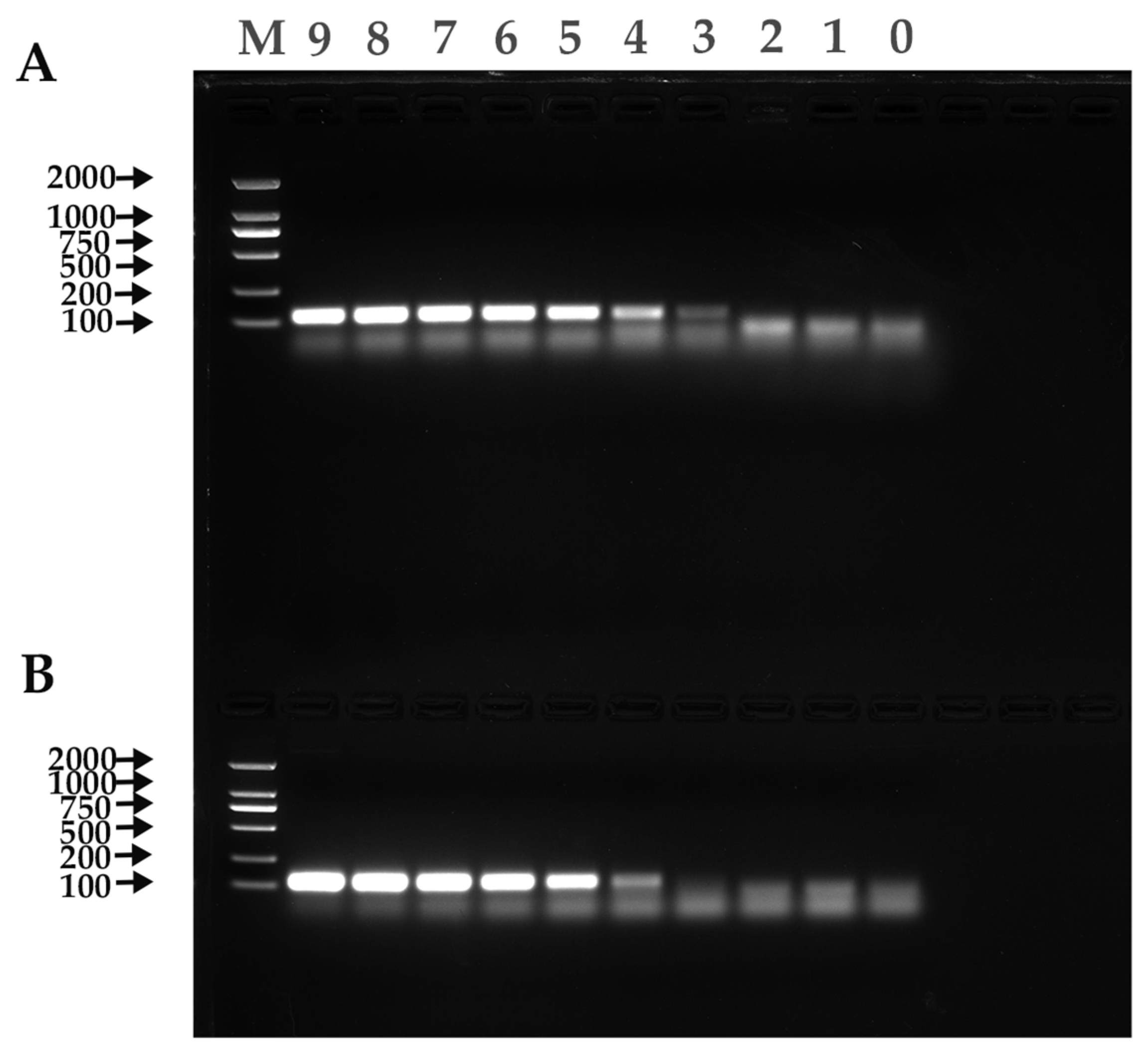
3.5.1. Evaluation of Sensitivity
3.5.2. Evaluation of Specificity
3.5.3. Evaluation of Repeatability and Stability
3.6. Comparison of qRT-PCR and RT-PCR Sensitivity
3.7. Tissue Tropism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, Z.; Qiu, Y.; Ge, X. The taxonomy, host range and pathogenicity of coronaviruses and other viruses in the Nidovirales order. Anim. Dis. 2021, 1, 5. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Yip, C.C.; Huang, Y.; Tsoi, H.W.; Chan, K.H.; Yuen, K.Y. Comparative analysis of 22 coronavirus HKU1 genomes reveals a novel genotype and evidence of natural recombination in coronavirus HKU1. J. Virol. 2006, 80, 7136–7145. [Google Scholar] [CrossRef]
- Hamre, D.; Procknow, J.J. A new virus isolated from the human respiratory tract. Proc. Soc. Exp. Biol. Med. 1966, 121, 190–193. [Google Scholar] [CrossRef]
- Lau, S.K.; Woo, P.C.; Li, K.S.; Tsang, A.K.; Fan, R.Y.; Luk, H.K.; Cai, J.P.; Chan, K.H.; Zheng, B.J.; Wang, M.; et al. Discovery of a novel coronavirus, China Rattus coronavirus HKU24, from Norway rats supports the murine origin of Betacoronavirus 1 and has implications for the ancestor of Betacoronavirus lineage A. J. Virol. 2015, 89, 3076–3092. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Song, H.D.; Tu, C.C.; Zhang, G.W.; Wang, S.Y.; Zheng, K.; Lei, L.C.; Chen, Q.X.; Gao, Y.W.; Zhou, H.Q.; Xiang, H.; et al. Cross-host evolution of severe acute respiratory syndrome coronavirus in palm civet and human. Proc. Natl. Acad. Sci. USA 2005, 102, 2430–2435. [Google Scholar] [CrossRef]
- Ge, X.Y.; Li, J.L.; Yang, X.L.; Chmura, A.A.; Zhu, G.; Epstein, J.H.; Mazet, J.K.; Hu, B.; Zhang, W.; Peng, C.; et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 2013, 503, 535–538. [Google Scholar] [CrossRef]
- Hu, B.; Zeng, L.P.; Yang, X.L.; Ge, X.Y.; Zhang, W.; Li, B.; Xie, J.Z.; Shen, X.R.; Zhang, Y.Z.; Wang, N.; et al. Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus. PLoS Pathog. 2017, 13, e1006698. [Google Scholar] [CrossRef]
- Azhar, E.I.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for camel-to-human transmission of MERS coronavirus. N. Engl. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef]
- Kin, N.; Miszczak, F.; Diancourt, L.; Caro, V.; Moutou, F.; Vabret, A.; Ar Gouilh, M. Comparative molecular epidemiology of two closely related coronaviruses, bovine coronavirus (BCoV) and human coronavirus OC43 (HCoV-OC43), reveals a different evolutionary pattern. Infect. Genet. Evol. 2016, 40, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Malik, Y.A. Properties of Coronavirus and SARS-CoV-2. Malays J. Pathol. 2020, 42, 3–11. [Google Scholar] [PubMed]
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef] [PubMed]
- Cheever, F.S.; Daniels, J.B. A murine virus (JHM) causing disseminated encephalomyelitis with extensive destruction of myelin. J. Exp. Med. 1949, 90, 181–210. [Google Scholar] [CrossRef]
- Parker, J.C.; Cross, S.S.; Rowe, W.P. Rat coronavirus (RCV): A prevalent, naturally occurring pneumotropic virus of rats. Arch. Gesamte Virusforsch. 1970, 31, 293–302. [Google Scholar] [CrossRef]
- Wang, W.; Lin, X.D.; Guo, W.P.; Zhou, R.H.; Wang, M.R.; Wang, C.Q.; Ge, S.; Mei, S.H.; Li, M.H.; Shi, M.; et al. Discovery, diversity and evolution of novel coronaviruses sampled from rodents in China. Virology 2015, 474, 19–27. [Google Scholar] [CrossRef]
- Wu, Z.; Lu, L.; Du, J.; Yang, L.; Ren, X.; Liu, B.; Jiang, J.; Yang, J.; Dong, J.; Sun, L.; et al. Comparative analysis of rodent and small mammal viromes to better understand the wildlife origin of emerging infectious diseases. Microbiome 2018, 6, 178. [Google Scholar] [CrossRef]
- Wasberg, A.; Raghwani, J.; Li, J.; Pettersson, J.H.; Lindahl, J.F.; Lundkvist, A.; Ling, J. Discovery of a Novel Coronavirus in Swedish Bank Voles (Myodes glareolus). Viruses 2022, 14, 1205. [Google Scholar] [CrossRef]
- Tsoleridis, T.; Onianwa, O.; Horncastle, E.; Dayman, E.; Zhu, M.; Danjittrong, T.; Wachtl, M.; Behnke, J.M.; Chapman, S.; Strong, V.; et al. Discovery of Novel Alphacoronaviruses in European Rodents and Shrews. Viruses 2016, 8, 84. [Google Scholar] [CrossRef]
- Lwande, O.W.; Mohamed, N.; Bucht, G.; Ahlm, C.; Olsson, G.; Evander, M. Seewis hantavirus in common shrew (Sorex araneus) in Sweden. Virol. J. 2020, 17, 198. [Google Scholar] [CrossRef]
- Ge, X.Y.; Yang, W.H.; Zhou, J.H.; Li, B.; Zhang, W.; Shi, Z.L.; Zhang, Y.Z. Detection of alpha- and betacoronaviruses in rodents from Yunnan, China. Virol. J. 2017, 14, 98. [Google Scholar] [CrossRef]
- FRANCIS2, A.G.-S.a.C.M. A new species of bat of the Hipposideros bicolor group (Chiroptera:Hipposideridae) from Central Laos, with evidence of convergent evolution with Sundaic taxa. Acta Chiropterologica 2006, 8, 39–61. [Google Scholar]
- Watanabe, S.; Masangkay, J.S.; Nagata, N.; Morikawa, S.; Mizutani, T.; Fukushi, S.; Alviola, P.; Omatsu, T.; Ueda, N.; Iha, K.; et al. Bat coronaviruses and experimental infection of bats, the Philippines. Emerg. Infect. Dis. 2010, 16, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-H.; Rawal, G.; Aljets, E.; Yim-Im, W.; Yang, Y.-L.; Huang, Y.-W.; Krueger, K.; Gauger, P.; Main, R.; Zhang, J. Development and Clinical Applications of a 5-Plex Real-Time RT-PCR for Swine Enteric Coronaviruses. Viruses 2022, 14, 1536. [Google Scholar] [CrossRef] [PubMed]
- Hemida, M.G.; Chu, D.K.; Poon, L.L.; Perera, R.A.; Alhammadi, M.A.; Ng, H.Y.; Siu, L.Y.; Guan, Y.; Alnaeem, A.; Peiris, M. MERS coronavirus in dromedary camel herd, Saudi Arabia. Emerg. Infect. Dis. 2014, 20, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- Hemida, M.G.; Elmoslemany, A.; Al-Hizab, F.; Alnaeem, A.; Almathen, F.; Faye, B.; Chu, D.K.; Perera, R.A.; Peiris, M. Dromedary Camels and the Transmission of Middle East Respiratory Syndrome Coronavirus (MERS-CoV). Transbound Emerg. Dis. 2017, 64, 344–353. [Google Scholar] [CrossRef]
- Weidinger, P.; Kolodziejek, J.; Khafaga, T.; Loney, T.; Howarth, B.; Sher Shah, M.; Abou Tayoun, A.; Alsheikh-Ali, A.; Camp, J.V.; Nowotny, N. Potentially Zoonotic Viruses in Wild Rodents, United Arab Emirates, 2019-A Pilot Study. Viruses 2023, 15, 695. [Google Scholar] [CrossRef]
- Chidoti, V.; De Nys, H.; Pinarello, V.; Mashura, G.; Missé, D.; Guerrini, L.; Pfukenyi, D.; Cappelle, J.; Chiweshe, N.; Ayouba, A.; et al. Longitudinal Survey of Coronavirus Circulation and Diversity in Insectivorous Bat Colonies in Zimbabwe. Viruses 2022, 14, 781. [Google Scholar] [CrossRef]
- Xiao, F.; Tang, M.; Zheng, X.; Liu, Y.; Li, X.; Shan, H. Evidence for Gastrointestinal Infection of SARS-CoV-2. Gastroenterology 2020, 158, 1831–1833.e1833. [Google Scholar] [CrossRef]
- Chan, K.H.; Poon, L.L.; Cheng, V.; Guan, Y.; Hung, I.; Kong, J.; Yam, L.Y.; Seto, W.H.; Yuen, K.Y.; Peiris, J.S.M. Detection of SARS coronavirus in patients with suspected SARS. Emerg. Infect. Dis. 2004, 10, 294–299. [Google Scholar] [CrossRef]
- Latif, A.A.; Mukaratirwa, S. Zoonotic origins and animal hosts of coronaviruses causing human disease pandemics: A review. Onderstepoort J. Vet. Res. 2020, 87, e1–e9. [Google Scholar] [CrossRef] [PubMed]
- Lane, T.E.; Hosking, M.P. The pathogenesis of murine coronavirus infection of the central nervous system. Crit. Rev. Immunol. Vol. 2010, 30, 119–130. [Google Scholar]
- Weiner, L.P. Pathogenesis of demyelination induced by a mouse hepatitis. Arch. Neurol. 1973, 28, 298–303. [Google Scholar] [CrossRef]
- De Albuquerque, N.; Baig, E.; Ma, X.; Zhang, J.; He, W.; Rowe, A.; Habal, M.; Liu, M.; Shalev, I.; Downey, G.P.; et al. Murine hepatitis virus strain 1 produces a clinically relevant model of severe acute respiratory syndrome in A/J mice. J. Virol. 2006, 80, 10382–10394. [Google Scholar] [CrossRef]
- Funk, C.J.; Manzer, R.; Miura, T.A.; Groshong, S.D.; Ito, Y.; Travanty, E.A.; Leete, J.; Holmes, K.V.; Mason, R.J. Rat respiratory coronavirus infection: Replication in airway and alveolar epithelial cells and the innate immune response. J. Gen. Virol. 2009, 90, 2956–2964. [Google Scholar] [CrossRef]
- Liang, S.C.; Schoeb, T.R.; Davis, J.K.; Simecka, J.W.; Cassell, G.H.; Lindsey, J.R. Comparative Severity of Respiratory Lesions of Sialodacryoadenitis Virus and Sendai Virus Infections in LEW and F344 Rats. Vet. Pathol. 1995, 32, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Alekseev, K.P.; Vlasova, A.N.; Jung, K.; Hasoksuz, M.; Zhang, X.; Halpin, R.; Wang, S.; Ghedin, E.; Spiro, D.; Saif, L.J. Bovine-like coronaviruses isolated from four species of captive wild ruminants are homologous to bovine coronaviruses, based on complete genomic sequences. J. Virol. 2008, 82, 12422–12431. [Google Scholar] [CrossRef]
- Harrison, A.G.; Lin, T.; Wang, P. Mechanisms of SARS-CoV-2 Transmission and Pathogenesis. Trends Immunol. 2020, 41, 1100–1115. [Google Scholar] [CrossRef]
- Jin, L.; Cebra, C.K.; Baker, R.J.; Mattson, D.E.; Cohen, S.A.; Alvarado, D.E.; Rohrmann, G.F. Analysis of the genome sequence of an alpaca coronavirus. Virology 2007, 365, 198–203. [Google Scholar] [CrossRef]
- Lorusso, A.; Desario, C.; Mari, V.; Campolo, M.; Lorusso, E.; Elia, G.; Martella, V.; Buonavoglia, C.; Decaro, N. Molecular characterization of a canine respiratory coronavirus strain detected in Italy. Virus Res. 2009, 141, 96–100. [Google Scholar] [CrossRef]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Pfefferle, S.; Oppong, S.; Drexler, J.F.; Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Müller, M.A.; Annan, A.; Vallo, P.; Adu-Sarkodie, Y.; et al. Distant Relatives of Severe Acute Respiratory Syndrome Coronavirus and Close Relatives of Human Coronavirus 229E in Bats, Ghana. Emerg. Infect. Dis. 2009, 15, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Vijgen, L.; Keyaerts, E.; Lemey, P.; Maes, P.; Van Reeth, K.; Nauwynck, H.; Pensaert, M.; Van Ranst, M. Evolutionary history of the closely related group 2 coronaviruses: Porcine hemagglutinating encephalomyelitis virus, bovine coronavirus, and human coronavirus OC43. J. Virol. 2006, 80, 7270–7274. [Google Scholar] [CrossRef] [PubMed]
- Vijgen, L.; Keyaerts, E.; Moes, E.; Thoelen, I.; Wollants, E.; Lemey, P.; Vandamme, A.M.; Van Ranst, M. Complete genomic sequence of human coronavirus OC43: Molecular clock analysis suggests a relatively recent zoonotic coronavirus transmission event. J. Virol. 2005, 79, 1595–1604. [Google Scholar] [CrossRef]
- Goraichuk, I.V.; Arefiev, V.; Stegniy, B.T.; Gerilovych, A.P. Zoonotic and Reverse Zoonotic Transmissibility of SARS-CoV-2. Virus Res. 2021, 302, 198473. [Google Scholar] [CrossRef]
- Berto, A.; Anh, P.H.; Carrique-Mas, J.J.; Simmonds, P.; Van Cuong, N.; Tue, N.T.; Van Dung, N.; Woolhouse, M.E.; Smith, I.; Marsh, G.A.; et al. Detection of potentially novel paramyxovirus and coronavirus viral RNA in bats and rats in the Mekong Delta region of southern Viet Nam. Zoonoses Public Health 2018, 65, 30–42. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′→3′) | bp | Amplify the Region | References | |
|---|---|---|---|---|---|
| CoV RT-PCR primer | CoV-FWD3 | GGTTGGGAYTAYCCHAARTGTGA | 434 bp | RdRp | [23] |
| CoV-FWD4/other | GAYTAYCCHAARTGTGAUMGWGC | ||||
| CoV-RVS3 | CCATCATCASWYRAATCATCATA | ||||
| α-RCoV qRT-PCR primer and probe | α-RCoV-F14493 | ACATCTGGTGATGCTAGTAC | 110 bp | RdRp | This study |
| α-RCoV-R14602 | TTCCTRCAAACATTACTATCAACAG | ||||
| α-RCoV-Probe | FAM-TTTTCAGGCTGTTAGTGCTAATGTAAATAAATTGC-BHQ1 | ||||
| β-RCoV qRT-PCR primer and probe | β-RCoV-F15324 | AGTATGATGATTTTGAGTGATGATGGYGTTG | 117 bp | RdRp | This study |
| β-RCoV-R15440 | CACGTTATTTTGATAATACAGCACCTGTTG | ||||
| β-RCoV-Probe | FAM-TATGCGTCCAAAGGTTATATTGCTAATATTAGTGCCT-BHQ1 | ||||
| Rodent identification primer | L14724 | ATGATATGAAAAACCATCGTTG | 1200 bp | mt-Cytb | [22] |
| H15915 | TTTCCNTTTCTGGTTTACAAGAC |
| Order | Species | Locations | Composition, % | Prevalence, % | ||
|---|---|---|---|---|---|---|
| qRT-PCR | RT-PCR | |||||
| α-CoV | β-CoV | |||||
| Rodentia | Chevrieri’s field mouse (Apodemus chevrieri) | Dali, Nujiang | 22.51 (113/502) | 9.73 (11/113) | 0 (0/113) | 3.54 (4/113) |
| Lancangjiang field mouse (Apodemus ilex) | Dali, Nujiang | 17.93 (90/502) | 21.11 (19/90) | 0 (0/90) | 6.67 (6/90) | |
| Kachin red-backed vole (Eothenomys cachinus) | Dali, Nujiang | 16.14 (81/502) | 11.11 (9/81) | 2.22 (2/90) | 2.22 (2/90) | |
| Large oriental vole (Eothenomys miletus) | Dali, Nujiang | 7.77 (39/502) | 12.82 (5/39) | 0 (0/39) | 0 (0/39) | |
| Asian house rat (Rattus tanezumi) | Dali, Nujiang | 14.94 (75/502) | 25.33 (19/75) | 0 (0/75) | 0 (0/75) | |
| Norway rat (Rattus norvegicus) | Dali | 5.18 (26/502) | 46.15 (12/26) | 3.85 (1/26) | 3.85 (1/26) | |
| Grey bellied mouse (Niviventer eha) | Nujiang | 0.6 (3/502) | 0 (0/3) | 0 (0/3) | 0 (0/3) | |
| Ryukyu mouse (Mus caroli) | Dali | 0.6 (3/502) | 0 (0/3) | 0 (0/3) | 0 (0/3) | |
| House mouse (Mus musculus) | Dali | 0.40 (2/502) | 0 (0/2) | 0 (0/2) | 0 (0/2) | |
| Chestnut white-bellied rat (Niviventer fulvescens) | Dali, Nujiang | 0.40 (2/502) | 0 (0/2) | 0 (0/2) | 0 (0/2) | |
| White-footed Indochinese rat (Rattus nitidus) | Dali | 0.80 (4/502) | 75 (3/4) | 75 (3/4) | 0 (0/4) | |
| Swinhoe’s striped squirrel (Tamiops swinhoei) | Dali | 0.20 (1/502) | 0 (0/1) | 0 (0/1) | 0 (0/1) | |
| Insectivora | Short-tailed shrew (Blarina brevicauda) | Nujiang | 4.18 (21/502) | 4.67 (1/21) | 0 (0/21) | 0 (0/21) |
| Long-tailed red-toothed shrew (Episoriculus leucops) | Nujiang | 3.39 (17/502) | 5.88 (1/17) | 5.88 (1/17) | 0 (0/17) | |
| Asian gray shrew (Crocidura attenuata) | Nujiang | 2.19 (11/502) | 27.27 (3/11) | 0 (0/11) | 0 (0/11) | |
| House musk shrew (Suncus murinus) | Dali | 0.60 (3/502) | 66.67 (2/3) | 0 (0/3) | 0 (0/3) | |
| Lagomorpha | Tibet pika (Ochotona thibetana) | Nujiang | 1.79 (9/502) | 11.11 (1/9) | 0 (0/9) | 0 (0/9) |
| Scandentia | Tree shrew (Tupaia belangeri) | Dali | 0.40 (2/502) | 50 (1/2) | 0 (0/2) | 0 (0/2) |
| Total | 100 (502/502) | 17.33 (87/502) | 1.39 (7/502) | 2.59 (13/502) | ||
| Rectum | Liver | Heart | Spleen | Lung | Kidney | |
|---|---|---|---|---|---|---|
| Mean (copies/g) | 1.35 × 106 | 3.95 × 103 | 2.96 × 103 | 2.93 × 103 | 1.68 × 103 | 0.97 × 103 |
| SEM | 1.09 × 106 | 2.71 × 103 | 2.28 × 103 | 2.77 × 103 | 1.47 × 103 | 0.87 × 103 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, F.-H.; Han, P.-Y.; Tian, J.-W.; Zong, L.-D.; Yin, H.-M.; Zhao, J.-Y.; Yang, Z.; Kong, W.; Ge, X.-Y.; Zhang, Y.-Z. Detection of Alpha- and Betacoronaviruses in Small Mammals in Western Yunnan Province, China. Viruses 2023, 15, 1965. https://doi.org/10.3390/v15091965
Xu F-H, Han P-Y, Tian J-W, Zong L-D, Yin H-M, Zhao J-Y, Yang Z, Kong W, Ge X-Y, Zhang Y-Z. Detection of Alpha- and Betacoronaviruses in Small Mammals in Western Yunnan Province, China. Viruses. 2023; 15(9):1965. https://doi.org/10.3390/v15091965
Chicago/Turabian StyleXu, Fen-Hui, Pei-Yu Han, Jia-Wei Tian, Li-Dong Zong, Hong-Min Yin, Jun-Ying Zhao, Ze Yang, Wei Kong, Xing-Yi Ge, and Yun-Zhi Zhang. 2023. "Detection of Alpha- and Betacoronaviruses in Small Mammals in Western Yunnan Province, China" Viruses 15, no. 9: 1965. https://doi.org/10.3390/v15091965