
Domestic Cat Hepadnavirus and Pathogenic Retroviruses; A Sero-Molecular Survey of Cats in Santiago, Chile

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Population Sampled and Data Collection
2.3. Serology
2.4. Molecular Testing
2.4.1. Domestic Cat Hepadnavirus Quantitative and Conventional PCR Testing
2.4.2. DCH Genome Sequencing and Analysis

{kind=link}
{kind=link}
| Assay | Target Gene (Product Size) | Name | Purpose | Sequence | Cycling Conditions |
|---|---|---|---|---|---|
| DCH qPCR (Lanave et al. 2019) [5] | Polymerase (132 bp) | FHBV-for | Forward | CGTCATCATGGGTTTAGGAA | Initial Activation: 95 °C, 3 min Denaturation: 95 °C, 10 s Annealing-extension: 60 °C for 30 s40 cycles |
| FHBV-rev | Reverse | TCCATATAAGCAAACACCATACAAT | |||
| FHBV-prob | Probe | [FAM]TCCTCCTAACCATTGAAGCCAGACTACT [QSY] | |||
| DCH cPCR (Aghazadeh et al. 2018) [4] | Core (258 bp) | Hgap-F | Forward | CTAGAATGGCTACATGGGTTAG | Initial denaturation: 95 °C, 1 min Denaturation: 95 °C, 30 s Annealing: 55 °C, 30 s Extension: 72 °C, 1 min, 35 cycles Final extension: 72 °C, 5 min |
| Hgap-R | Reverse | GTGCTCTGATAACCGTATGCTC | |||
| FeLV nested PCR (Miyazawa et al. 1997) [28] | U3-gag region (1st round: 770 bp 2nd round: 601 bp) | U3-F(1) | 1st round Forward | ACAGCAGAAGTTTCAAGGCC | Initial denaturation: 95 °C, 2 min Denaturation: 95 °C, 45 s Annealing: 55 °C, 30 s Extension: 72 °C, 1 min, 35 cycles Final extension: 72 °C, 10 min |
| G-R(1) | 1st round Reverse | GACCAGTGATCAAGGGTGAG | |||
| U3-F(2) | 2nd round Forward | GCTCCCCAGTTGACCAGAGT | |||
| G-R(2) | 2nd round Reverse | GCTTCGGTACCAAACCGAAA |
| Primer Set | Nested PCR Round | Name | Primer Binding Site | Sequence | Product Size (bp) | Cycling Conditions | ||
|---|---|---|---|---|---|---|---|---|
| (Numbering Based on NC040719) | Denaturation | Annealing | Extension | |||||
| 1 | 1st round | 2239OF_1 | 2234-2253 | CAGAAGACGTACTCCCTYKC | 599 | 95 °C, 30 s | 57 °C, 30 s | 72 °C, 1 min |
| 2842OR_1 | 2832-2810 | TCYTTGAGGGGATTGTGRTCGAA | ||||||
| 2nd round | 2264IF_1 | 2259-2278 | AGACGCAGATCTCAATCTYC | 456 | 95 °C, 30 s | 55 °C, 30 s | 72 °C, 1 min | |
| 2724IR_1 | 2714-2694 | AARTGCAGTTTTTGTTCCCAG | ||||||
| 2 | 1st round | 2677OF_2 | 2676-2698 | TAATGGACAATTATACCCCTGGG | 575 | 95 °C, 30 s | 55 °C, 30 s | 72 °C, 1 min |
| 63OR_2 | 63-43 | CMAGAAAGATCCCACCTGTAA | ||||||
| 2nd round | 2695IF_2 | 2694-2714 | CTGGGAACAAAAACTGCAYTT | 444 | 95 °C, 30 s | 58 °C, 30 s | 72 °C, 1 min | |
| 3138IR_2 | 3137-3118 | CAGTAGGGGCCACAGTAGTC | ||||||
| 3 | 1st round | 2914OF_3 | 2913-2932 | CTCTCAGTTTCGCAACCCAG | 609 | 95 °C, 30 s | 55 °C, 30 s | 72 °C, 1 min |
| 334OR_3 | 2996-3015 | CAGGAGGAAAAGGACATACC | ||||||
| 2nd round | 2997IF_3 | 262-240 | CCTTTAACTCCTCCTGTTCG | 454 | 95 °C, 30 s | 58 °C, 30 s | 72 °C, 1 min | |
| 262IR_3 | 334-315 | CCAGGAGCAAGAGGTAAATGATA | ||||||
| 4 | 1st round | FHBV-F | 458-478 | CGTCATCATGGGCTTTAGGAA | 575 | 95 °C, 30 s | 57 °C, 30 s | 72 °C, 1 min |
| 1032OR_4 | 1032-1012 | CAAACACCTGGCARGACATTG | ||||||
| 2nd round | 572IF_4 | 572-594 | GGTGTTTGCTTATATGGATGAYG | 428 | 95 °C, 30 s | 57 °C, 30 s | 72 °C, 1 min | |
| 999IR_4 | 999-979 | GRACAAAGTACAAGTGTGTGT | ||||||
| 5 | 1st round | 687F_2+ | 687-706 | ACAAAAACCAAACGCTGGGG | 811 | 95 °C, 30 s | 55 °C, 30 s | 72 °C, 1 min |
| 1497OR_5 | 1445-1428 | GGCACRAGTCCAATCATG | ||||||
| 2nd round | 895OF_5 | 895-915 | CTTTRATGCCTTTGTATACCG | 551 | 95 °C, 30 s | 57 °C, 30 s | 72 °C, 1 min | |
| 1445IR_5 | 1497-1480 | TGAARCGAAGGACACACG | ||||||
| 6 | 1st round | 1752OF_7 | 1751-1770 | GGACATTGACCCTTATAAAG | 600 | 95 °C, 30 s | 53 °C, 30 s | 72 °C, 1 min |
| 2351OR_7 | 2350-2332 | TCAAAGACCCCCATTTATG | ||||||
| 2nd round | 1798IF_7 | 1797-1816 | TCTTTTTTGCCGTCTGACTT | 503 | 95 °C, 30 s | 56 °C, 30 s | 72 °C, 1 min | |
| 2300IR_7 | 2299-2282 | GATTGAGARCGTCTGCGA | ||||||
2.4.3. FeLV Quantitative and Nested PCR Testing
2.4.4. Interpretation of Infection Status
2.5. Statistical Analyses
3. Results
3.1. Population Characteristics
3.2. Domestic Cat Hepadnavirus Detection and Sequencing in Chilean Cats
3.3. Phylogenetic Analysis of DCH Chile Sequence
3.4. Retrovirus Infections in Chilean Cats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Budget Directorate (Chile), S.M.R. Responsible Pet Ownership Program. Under Secretary of Regional and Administrative Development. Ministry of the Interior and Public Security. Available online: http://bibliotecadigital.dipres.gob.cl/handle/11626/19316?show=full (accessed on 1 November 2023).
- Undersecretary of Development Regional and Administrative (SUBDERE). Estimation of the Canine Feline Population of the Country and the Diagnosis of Responsible Ownerships. Available online: https://www.studocu.com/cl/document/pontificia-universidad-catolica-de-chile/bases-y-fundamentos-de-la-medicina-i/analisis-poblacion-canina-y-felina-en-chile-subdere-2022/66652204 (accessed on 11 December 2023).
- United Nations—Department of Economic and Social Affairs Population Division. World Population Prospects 2022. Available online: https://population.un.org/wpp/ (accessed on 11 December 2023).
- Aghazadeh, M.; Shi, M.; Barrs, V.; McLuckie, A.; Lindsay, S.; Jameson, B.; Hampson, B.; Holmes, E.; Beatty, J. A Novel Hepadnavirus Identified in an Immunocompromised Domestic Cat in Australia. Viruses 2018, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- Lanave, G.; Capozza, P.; Diakoudi, G.; Catella, C.; Catucci, L.; Ghergo, P.; Stasi, F.; Barrs, V.; Beatty, J.; Decaro, N.; et al. Identification of hepadnavirus in the sera of cats. Sci. Rep. 2019, 9, 10668. [Google Scholar] [CrossRef] [PubMed]
- Piewbang, C.; Wardhani, S.W.; Chaiyasak, S.; Yostawonkul, J.; Chai-in, P.; Boonrungsiman, S.; Kasantikul, T.; Techangamsuwan, S. Insights into the genetic diversity, recombination, and systemic infections with evidence of intracellular maturation of hepadnavirus in cats. PLoS ONE 2020, 15, e0241212. [Google Scholar] [CrossRef] [PubMed]
- Anpuanandam, K.; Selvarajah, G.T.; Choy, M.M.K.; Ng, S.W.; Kumar, K.; Ali, R.M.; Rajendran, S.K.; Ho, K.L.; Tan, W.S. Molecular detection and characterisation of Domestic Cat Hepadnavirus (DCH) from blood and liver tissues of cats in Malaysia. BMC Vet. Res. 2021, 17, 9. [Google Scholar] [CrossRef] [PubMed]
- Capozza, P.; Carrai, M.; Choi, Y.R.; Tu, T.; Nekouei, O.; Lanave, G.; Martella, V.; Beatty, J.A.; Barrs, V.R. Domestic Cat Hepadnavirus: Molecular Epidemiology and Phylogeny in Cats in Hong Kong. Viruses 2023, 15, 150. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Kaneko, Y.; Shibanai, A.; Yamamoto, S.; Katagiri, A.; Osuga, T.; Inoue, Y.; Kuroda, K.; Tanabe, M.; Okabayashi, T.; et al. Identification of domestic cat hepadnavirus from a cat blood sample in Japan. J. Vet. Med. Sci. 2022, 84, 648–652. [Google Scholar] [CrossRef]
- Stone, C.; Petch, R.; Gagne, R.B.; Nehring, M.; Tu, T.; Beatty, J.A.; VandeWoude, S. Prevalence and Genomic Sequence Analysis of Domestic Cat Hepadnavirus in the United States. Viruses 2022, 14, 2091. [Google Scholar] [CrossRef] [PubMed]
- Jeanes, E.C.; Wegg, M.L.; Mitchell, J.A.; Priestnall, S.L.; Fleming, L.; Dawson, C. Comparison of the prevalence of Domestic Cat Hepadnavirus in a population of cats with uveitis and in a healthy blood donor cat population in the United Kingdom. Vet. Ophthalmol. 2022, 25, 165–172. [Google Scholar] [CrossRef]
- Hartmann, K. Clinical Aspects of Feline Retroviruses: A Review. Viruses 2012, 4, 2684. [Google Scholar] [CrossRef]
- Helfer-Hungerbuehler, A.K.; Widmer, S.; Kessler, Y.; Riond, B.; Boretti, F.S.; Grest, P.; Lutz, H.; Hofmann Lehmann, R. Long-term follow up of feline leukemia virus infection and characterization of viral RNA loads using molecular methods in tissues of cats with different infection outcomes. Virus Res. 2015, 197, 137–150. [Google Scholar] [CrossRef]
- Giselbrecht, J.; Jähne, S.; Bergmann, M.; Meli, M.L.; Pineroli, B.; Boenzli, E.; Teichmann-Knorrn, S.; Zablotski, Y.; Pennisi, M.-G.; Layachi, N.; et al. Prevalence of Different Courses of Feline Leukaemia Virus Infection in Four European Countries. Viruses 2023, 15, 1718. [Google Scholar] [CrossRef] [PubMed]
- Fusco, G.; Marati, L.; Pugliese, A.; Levante, M.; Ferrara, G.; de Carlo, E.; Amoroso, M.G.; Montagnaro, S. Prevalence of feline leukemia virus and feline immunodeficiency virus in cats from southern Italy: A 10-year cross-sectional study. Front. Vet. Sci. 2023, 10, 1260081. [Google Scholar] [CrossRef] [PubMed]
- Biezus, G.; Grima de Cristo, T.; da Silva Casa, M.; Lovatel, M.; Vavassori, M.; Brüggemann de Souza Teixeira, M.; Miletti, L.C.; Maciel da Costa, U.; Assis Casagrande, R. Progressive and regressive infection with feline leukemia virus (FeLV) in cats in southern Brazil: Prevalence, risk factors associated, clinical and hematologic alterations. Prev. Vet. Med. 2023, 216, 105945. [Google Scholar] [CrossRef] [PubMed]
- Leite-Filho, R.V.; Panziera, W.; Bandinelli, M.B.; Henker, L.C.; da Conceição Monteiro, K.; Corbellini, L.G.; Driemeier, D.; Sonne, L.; Pavarini, S.P. Epidemiological, pathological and immunohistochemical aspects of 125 cases of feline lymphoma in Southern Brazil. Vet. Comp. Oncol. 2020, 18, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Buch, J.; Beall, M.; O’Connor, T.; Chandrashekar, R. Worldwide clinic-based serologic survey of FIV antibody and FeLV antigens in cats, ID14, 2017 ACVIM Forum Research Abstract Program. J. Vet. Intern. Med. 2017, 31, 1225–1361. [Google Scholar]
- Spada, E.; Perego, R.; Sgamma, E.A.; Proverbio, D. Survival time and effect of selected predictor variables on survival in owned pet cats seropositive for feline immunodeficiency and leukemia virus attending a referral clinic in northern Italy. Prev. Vet. Med. 2018, 150, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Van Nguyen, D.; Van Nguyen, C.; Bonsall, D.; Ngo, T.T.; Carrique-Mas, J.; Pham, A.H.; Bryant, J.E.; Thwaites, G.; Baker, S.; Woolhouse, M.; et al. Detection and Characterization of Homologues of Human Hepatitis Viruses and Pegiviruses in Rodents and Bats in Vietnam. Viruses 2018, 10, 102. [Google Scholar] [CrossRef]
- Choi, Y.R.; Chen, M.C.; Carrai, M.; Rizzo, F.; Chai, Y.; Tse, M.; Jackson, K.; Martella, V.; Steiner, J.; Pesavento, P.A.; et al. Hepadnavirus DNA Is Detected in Canine Blood Samples in Hong Kong but Not in Liver Biopsies of Chronic Hepatitis or Hepatocellular Carcinoma. Viruses 2022, 14, 1543. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, T.; Jarrett, O. Feline leukaemia virus proviral DNA detected by polymerase chain reaction in antigenaemic but non-viraemic (‘discordant’) cats. Arch. Virol. 1997, 142, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A computer program for analyzing recombination in, and removing signals of recombination from, nucleotide sequence datasets. Virus Evol. 2021, 7, veaa087. [Google Scholar] [CrossRef] [PubMed]
- Padidam, M.; Sawyer, S.; Fauquet, C.M. Possible emergence of new geminiviruses by frequent recombination. Virology 1999, 265, 218–225. [Google Scholar] [CrossRef]
- Martin, D.P.; Posada, D.; Crandall, K.A.; Williamson, C. A modified bootscan algorithm for automated identification of recombinant sequences and recombination breakpoints. AIDS Res. Hum. Retroviruses 2005, 21, 98–102. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Evaluation of methods for detecting recombination from DNA sequences: Computer simulations. Proc. Natl. Acad. Sci. USA 2001, 98, 13757–13762. [Google Scholar] [CrossRef]
- Gibbs, M.J.; Armstrong, J.S.; Gibbs, A.J. Sister-Scanning: A Monte Carlo procedure for assessing signals in recombinant sequences. Bioinformatics 2000, 16, 573–582. [Google Scholar] [CrossRef]
- Lam, H.M.; Ratmann, O.; Boni, M.F. Improved Algorithmic Complexity for the 3SEQ Recombination Detection Algorithm. Mol. Biol. Evol. 2018, 35, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Ito, G.; Goto-Koshino, Y.; Sakamoto, M.; Nishimura, R.; Momoi, Y. Detection of domestic cat hepadnavirus by next-generation sequencing and epidemiological survey in Japan. J. Vet. Med. Sci. 2023, 85, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.S.Y.; Covert, E.; Wilson, E.; Kottilil, S. Chronic Hepatitis B Infection. JAMA 2018, 319, 1802. [Google Scholar] [CrossRef] [PubMed]
- Bencic, S.; Orel, B.; Surca, A.; Stangar, U.L. Structural and electrochromic properties of nanosized Fe/V-oxide films with FeVo(4) and Fe2V4O13 grains: Comparative studies with crystalline V2O5. Sol. Energy 2000, 68, 499–515. [Google Scholar] [CrossRef]
- Fruci, P.; Di Profio, F.; Palombieri, A.; Massirio, I.; Lanave, G.; Diakoudi, G.; Pellegrini, F.; Marsilio, F.; Martella, V.; Di Martino, B. Detection of antibodies against domestic cat hepadnavirus using baculovirus-expressed core protein. Transbound. Emerg. Dis. 2022, 69, 2980–2986. [Google Scholar] [CrossRef] [PubMed]
- Gleich, S.E.; Krieger, S.; Hartmann, K. Prevalence of feline immunodeficiency virus and feline leukaemia virus among client-owned cats and risk factors for infection in Germany. J. Feline Med. Surg. 2009, 11, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Azócar-Aedo, L.; Monti, G. Risk factors for seropositivity to feline retroviruses among owned domestic cats in Valdivia, southern Chile. Curr. Sci. 2018, 114, 1548–1553. [Google Scholar] [CrossRef]
- Sacristán, I.; Sieg, M.; Acuña, F.; Aguilar, E.; García, S.; López, M.J.; Cevidanes, A.; Hidalgo-Hermoso, E.; Cabello, J.; Vahlenkamp, T.W.; et al. Molecular and serological survey of carnivore pathogens in free-roaming domestic cats of rural communities in southern Chile. J. Vet. Med. Sci. 2019, 81, 1740–1748. [Google Scholar] [CrossRef]
- Mora, M.; Napolitano, C.; Ortega, R.; Poulin, E.; Pizarro-Lucero, J. Feline immunodeficiency virus and feline leukemia virus infection in free-ranging guignas (Leopardus guigna) and sympatric domestic cats in human perturbed landscapes on chiloé island, chile. J. Wildl. Dis. 2015, 51, 199–208. [Google Scholar] [CrossRef]
- Sacristán, I.; Acuña, F.; Aguilar, E.; García, S.; José López, M.; Cabello, J.; Hidalgo-Hermoso, E.; Sanderson, J.; Terio, K.A.; Barrs, V.; et al. Cross-species transmission of retroviruses among domestic and wild felids in human-occupied landscapes in Chile. Evol. Appl. 2021, 14, 1070–1082. [Google Scholar] [CrossRef]

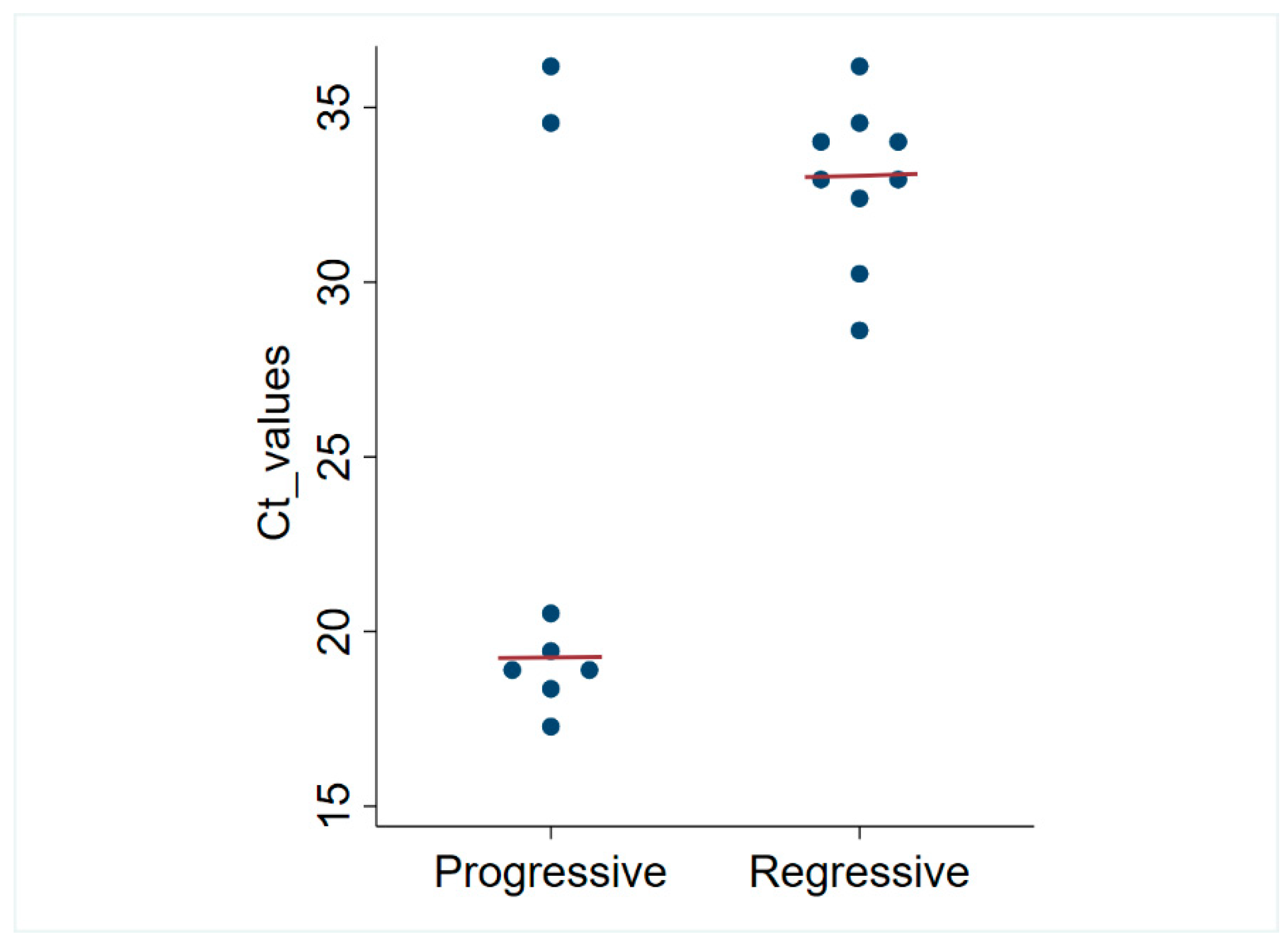
| Combined Testing Results | |||||
|---|---|---|---|---|---|
| FeLV p27 Antigen | FeLV qPCR | FIV Antibody | DCH cPCR | Frequency (%) | Interpretation of Test Results |
| Positive | Positive | Negative | Negative | 6/120 (5%) | Progressive FeLV infection |
| Positive | Positive | Positive | Negative | 2/120 (1.7%) | Dual FeLV/FIV infected |
| Negative | Positive | Negative | Negative | 9/120 (7.5%) | Regressive FeLV infection |
| Positive | Negative | Negative | Negative | 1/120 (0.8%) | False positive FeLV antigen result |
| Negative | Negative | Positive | Negative | 2/120 (1.7%) | FIV infected |
| Negative | Negative | Negative | Positive | 2 */120 (1.7%) | DCH infected |
| Negative | Negative | Negative | Negative | 98/120 (81.7%) | None of the infectious agents detected |
| Variable | Categories | Progressive FeLV Infection(qPCR and p27 Positive) (%) | p Value |
|---|---|---|---|
| Age | ≤3 years old | 2/26 (7.69%) | 1.000 |
| >3 years old | 6/82 (7.32%) | ||
| Sex | Female | 0/66 (0.00%) | 0.001 * |
| Male | 8/54 (14.81%) | ||
| Environment | access to outdoors | 7/94 (7.45%) | 1.000 |
| Indoors only | 1/26 (3.85%) | ||
| Source | Client-owned | 8/95 (8.42%) | 0.202 |
| Shelter-owned | 0/25 (0.00%) | ||
| Health status | Healthy | 5/83 (6.02%) | 0.699 |
| Sick | 3/37 (8.11%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.R.; Iturriaga, M.P.; Nekouei, O.; Tu, T.; Van Brussel, K.; Barrs, V.R.; Beatty, J.A. Domestic Cat Hepadnavirus and Pathogenic Retroviruses; A Sero-Molecular Survey of Cats in Santiago, Chile. Viruses 2024, 16, 46. https://doi.org/10.3390/v16010046
Choi YR, Iturriaga MP, Nekouei O, Tu T, Van Brussel K, Barrs VR, Beatty JA. Domestic Cat Hepadnavirus and Pathogenic Retroviruses; A Sero-Molecular Survey of Cats in Santiago, Chile. Viruses. 2024; 16(1):46. https://doi.org/10.3390/v16010046
Chicago/Turabian StyleChoi, Yan Ru, María Paz Iturriaga, Omid Nekouei, Thomas Tu, Kate Van Brussel, Vanessa R. Barrs, and Julia A. Beatty. 2024. "Domestic Cat Hepadnavirus and Pathogenic Retroviruses; A Sero-Molecular Survey of Cats in Santiago, Chile" Viruses 16, no. 1: 46. https://doi.org/10.3390/v16010046