
Photodynamic Inactivation of Bovine Coronavirus with the Photosensitizer Toluidine Blue O

 ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cultivation of MDBK Cells
2.3. Evaluation of In Vitro Cytotoxicity with the MTT-Dye Assay
2.4. Determination of MDBK Specific Growth Rate
- Biomass balance:
- 2.
- Solution to the differential equation:
- 3.
- After rearrangement of the form of the Equation (3), SGR is obtained as follows:
2.5. Mathematical Model for Calculation of Median Inhibitory and Maximal Non-Toxic Concentration
2.6. Propagation of Bovine Coronavirus in MDBK Cells
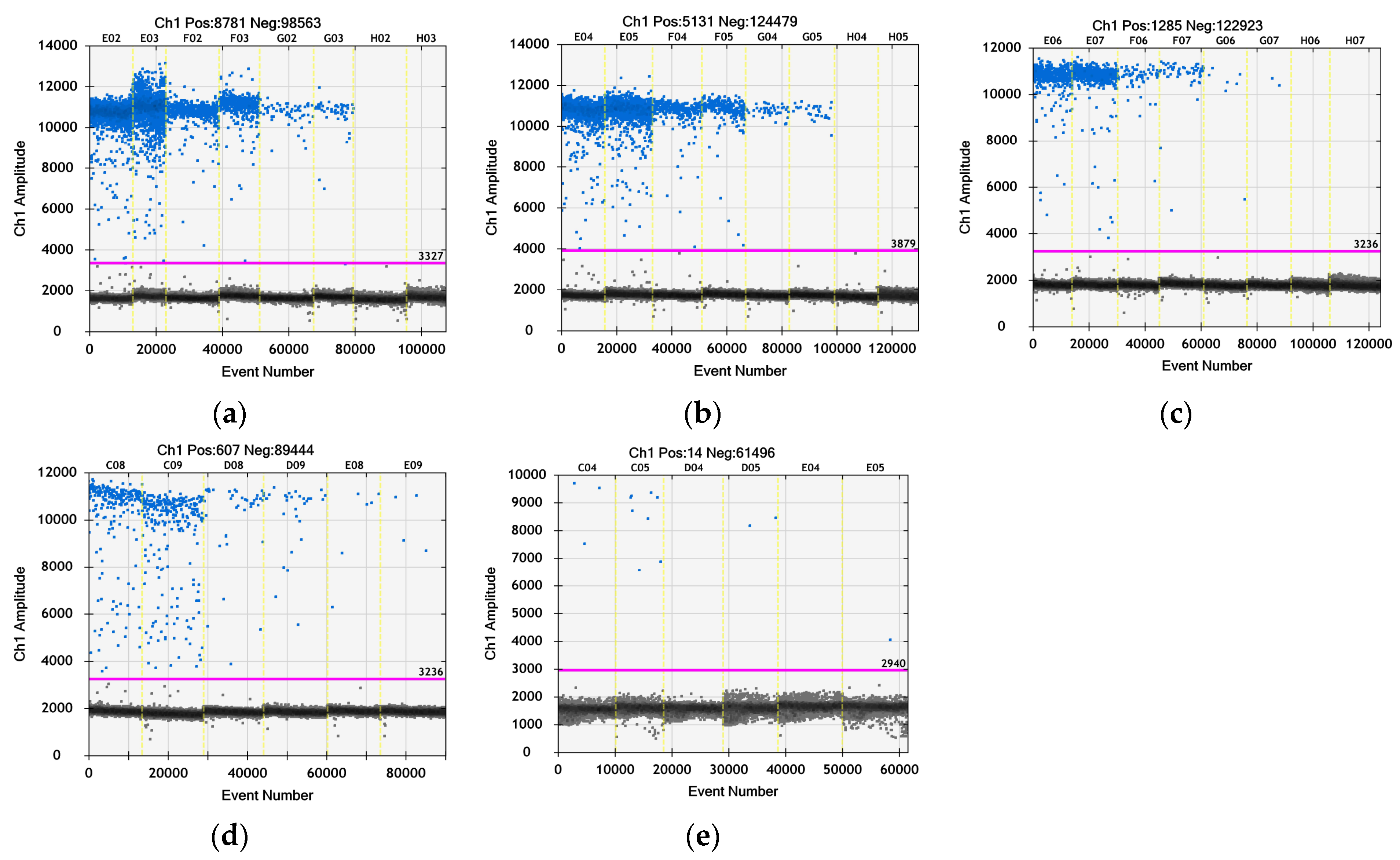
2.7. Determination of the Bovine Coronavirus Titer with Droplet Digital (ddPCR)
2.8. Determination of the Bovine Coronavirus Titer by TCID50 Assay
2.9. Determination of the Bovine Coronavirus Titer and Antiviral Activity of Photosensitizers by Plaque Assay
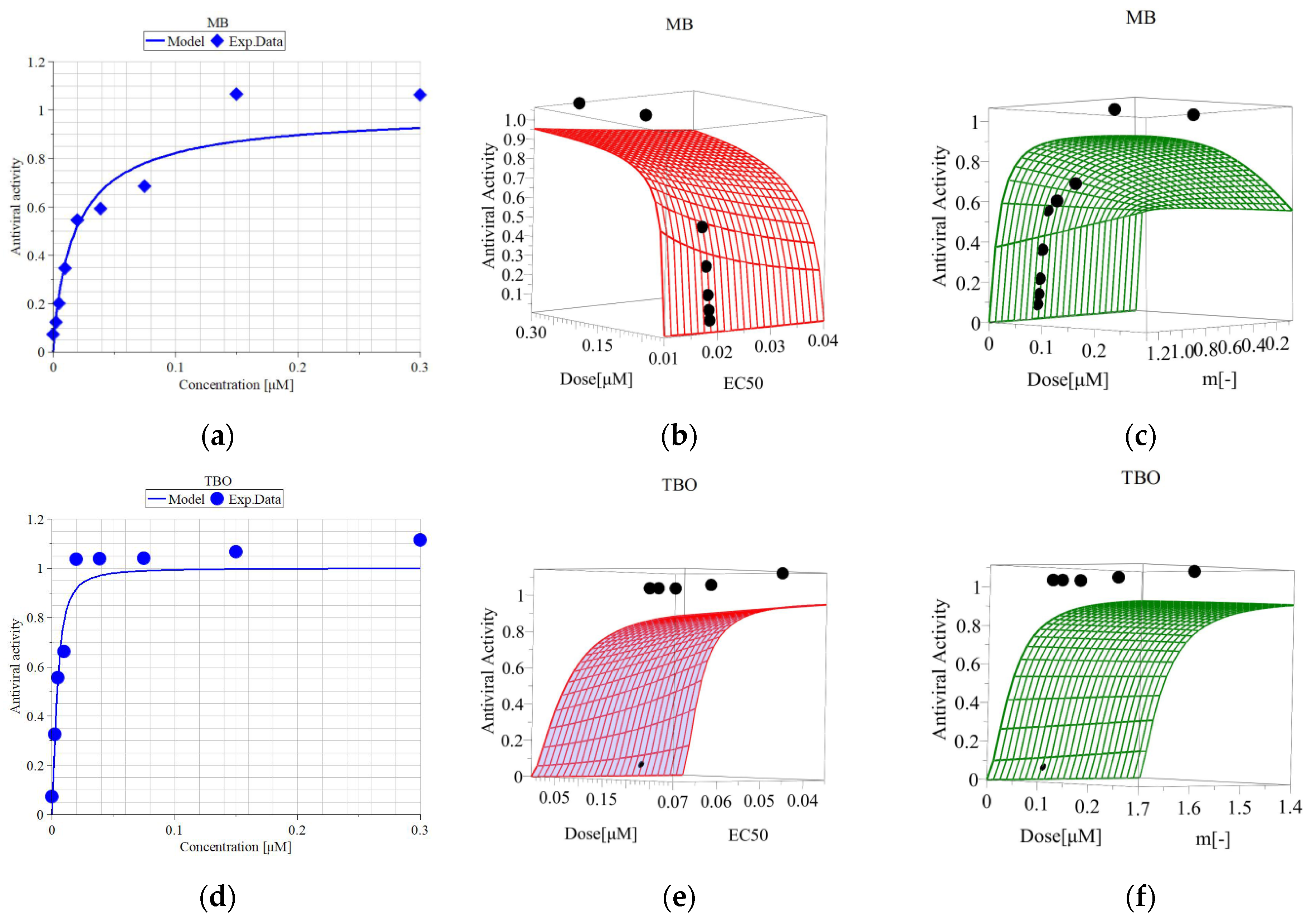
2.10. Determination of the Antiviral Activity of the Photosensitizers
2.11. Statistical Evaluation
3. Results
3.1. Specific Growth Rate of MDBK Cells
3.2. In Vitro Cytotoxicity of MB and TBO on MDBK Cells
3.3. Quantitative Evaluation of Virus Titer
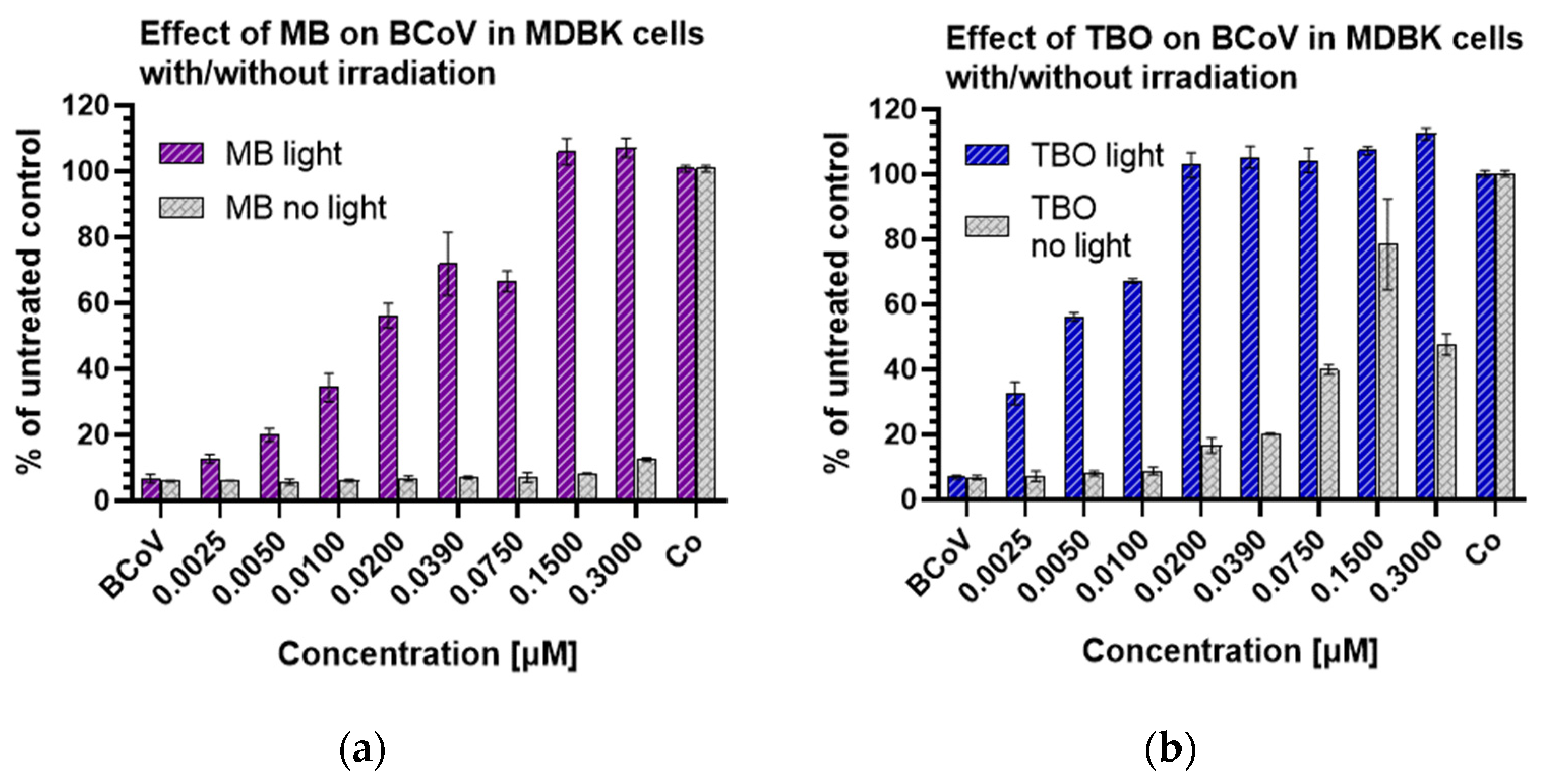
3.4. MB and TBO Inhibit the CPE of BCoV in MDBK Cells
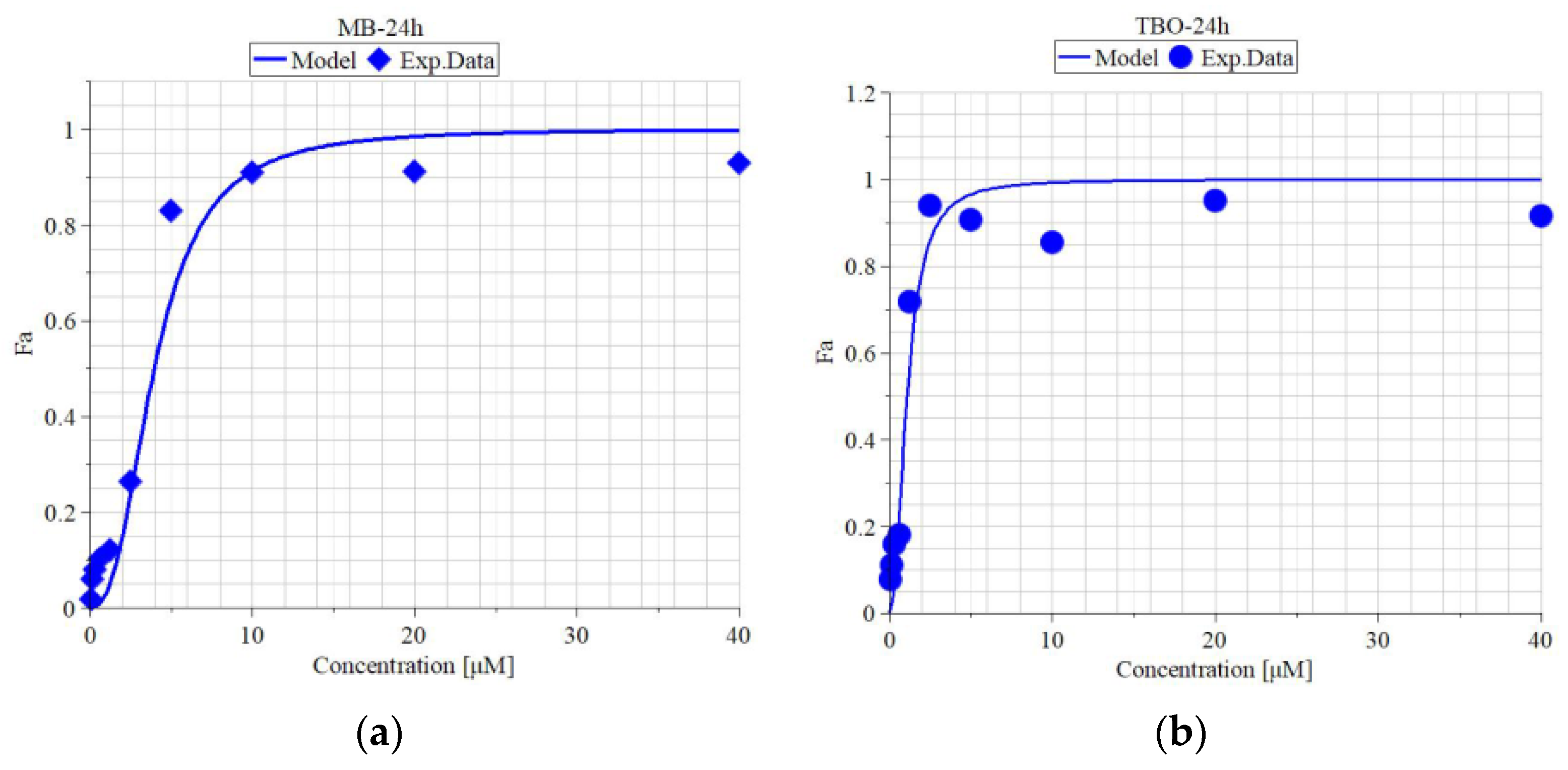
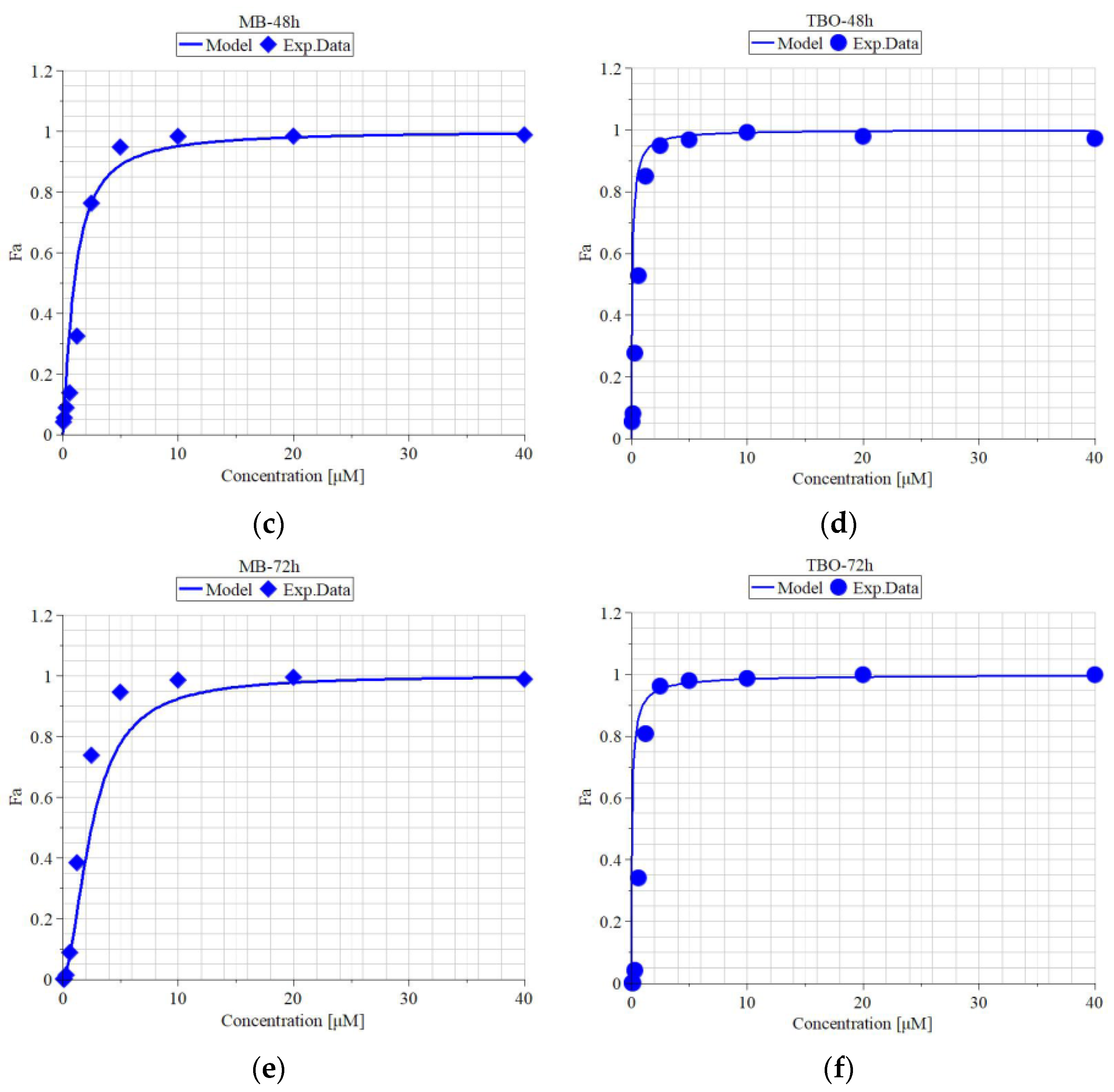
3.5. Metabolic Activity of MDBK Cells and Median Effective Concentrations of MB and TBO after Direct Inactivation of BCoV
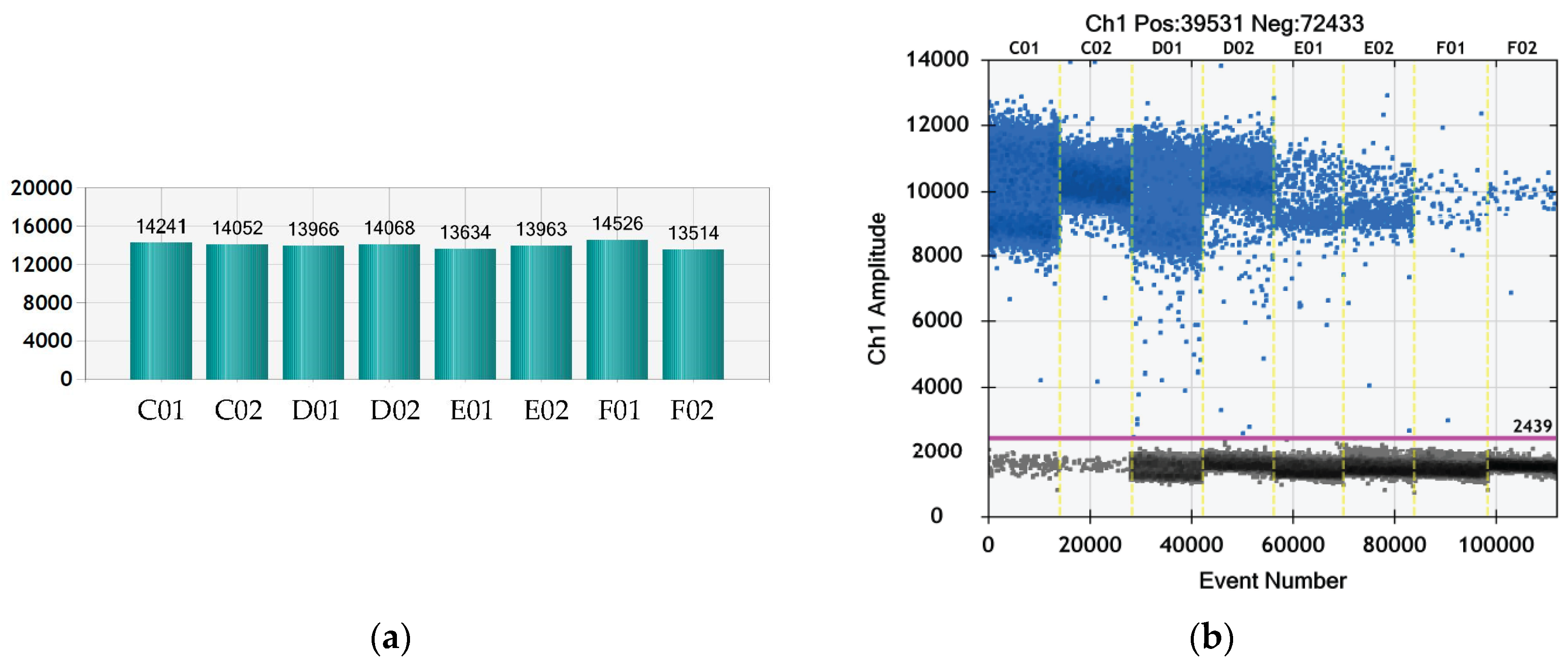
3.6. Quantitative Evaluation of the Virus Particles by ddPCR after Treatment with TBO
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 23 November 2023).
- Islam, A.; Ferdous, J.; Islam, S.; Sayeed, M.A.; Dutta Choudhury, S.; Saha, O.; Hassan, M.M.; Shirin, T. Evolutionary Dynamics and Epidemiology of Endemic and Emerging Coronaviruses in Humans, Domestic Animals, and Wildlife. Viruses 2021, 13, 1908. [Google Scholar] [CrossRef] [PubMed]
- Velavan, T.P.; Meyer, C.G. The COVID-19 epidemic. Trop. Med. Int. Health 2020, 25, 278–280. [Google Scholar] [CrossRef] [PubMed]
- Piret, J.; Boivin, G. Pandemics Throughout History. Front. Microbiol. 2021, 11, 631736. [Google Scholar] [CrossRef] [PubMed]
- David, D.; Storm, N.; Ilan, W.; Sol, A. Characterization of Winter Dysentery Bovine Coronavirus Isolated from Cattle in Israel. Viruses 2021, 13, 1070. [Google Scholar] [CrossRef] [PubMed]
- Abi, K.-m.; Zhang, Q.; Zhang, B.; Zhou, L.; Yue, H.; Tang, C. An emerging novel bovine coronavirus with a 4-amino-acid insertion in the receptor-binding domain of the hemagglutinin-esterase gene. Arch. Virol. 2020, 165, 3011–3015. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.F.S.; Santos, I.P.; Santos, G.M.P.; Bastos, T.M.; Rocha, V.P.C.; Meira, C.S.; Soares, M.B.P.; Pitta, I.R.; Pinheiro, A.L.B. Anti–Trypanosoma cruzi effect of the photodynamic antiparasitic chemotherapy using phenothiazine derivatives as photosensitizers. Lasers Med. Sci. 2020, 35, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Fekrazad, R.; Asefi, S.; Pourhajibagher, M.; Vahdatinia, F.; Fekrazad, S.; Bahador, A.; Abrahamse, H.; Hamblin, M.R. Photobiomodulation and Antiviral Photodynamic Therapy in COVID-19 Management. In Coronavirus Disease—COVID-19; Rezaei, N., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 517–547. [Google Scholar]
- Hamblin, M.R.; Hasan, T. Photodynamic therapy: A new antimicrobial approach to infectious disease? Photochem. Photobiol. Sci. 2004, 3, 436–450. [Google Scholar] [CrossRef]
- Jori, G.; Fabris, C.; Soncin, M.; Ferro, S.; Coppellotti, O.; Dei, D.; Fantetti, L.; Chiti, G.; Roncucci, G. Photodynamic therapy in the treatment of microbial infections: Basic principles and perspective applications. Lasers Surg. Med. 2006, 38, 468–481. [Google Scholar] [CrossRef]
- Moor, A.C.E. Signaling pathways in cell death and survival after photodynamic therapy. J. Photochem. Photobiol. B Biol. 2000, 57, 1–13. [Google Scholar] [CrossRef]
- Plotino, G.; Grande, N.M.; Mercade, M. Photodynamic therapy in endodontics. Int. Endod. J. 2019, 52, 760–774. [Google Scholar] [CrossRef]
- Costa, L.; Faustino, M.A.F.; Neves, M.G.P.M.S.; Cunha, Â.; Almeida, A. Photodynamic Inactivation of Mammalian Viruses and Bacteriophages. Viruses 2012, 4, 1034–1074. [Google Scholar] [CrossRef] [PubMed]
- Wiehe, A.; O’Brien, J.M.; Senge, M.O. Trends and targets in antiviral phototherapy. Photochem. Photobiol. Sci. 2019, 18, 2565–2612. [Google Scholar] [CrossRef] [PubMed]
- Remichkova, M.; Mukova, L.; Nikolaeva-Glomb, L.; Nikolova, N.; Doumanova, L.; Mantareva, V.; Angelov, I.; Kussovski, V.; Galabov, A.S. Virus inactivation under the photodynamic effect of phthalocyanine zinc(II) complexes. Z. Naturforschung C J. Biosci. 2017, 72, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Nikolaeva-Glomb, L.; Mukova, L.; Nikolova, N.; Kussovski, V.; Doumanova, L.; Mantareva, V.; Angelov, I.; Wöhrle, D.; Galabov, A.S. Photodynamic Effect of some Phthalocyanines on Enveloped and Naked Viruses. Acta Virol. 2017, 61, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Sadraeian, M.; da Cruz, E.F.; Boyle, R.W.; Bahou, C.; Chudasama, V.; Janini, L.M.R.; Diaz, R.S.; Guimarães, F.E.G. Photoinduced Photosensitizer–Antibody Conjugates Kill HIV Env-Expressing Cells, Also Inactivating HIV. ACS Omega 2021, 6, 16524–16534. [Google Scholar] [CrossRef] [PubMed]
- Korneev, D.; Kurskaya, O.; Sharshov, K.; Eastwood, J.; Strakhovskaya, M. Ultrastructural Aspects of Photodynamic Inactivation of Highly Pathogenic Avian H5N8 Influenza Virus. Viruses 2019, 11, 955. [Google Scholar] [CrossRef] [PubMed]
- Sharshov, K.; Solomatina, M.; Kurskaya, O.; Kovalenko, I.; Kholina, E.; Fedorov, V.; Meerovich, G.; Rubin, A.; Strakhovskaya, M. The Photosensitizer Octakis(cholinyl)zinc Phthalocyanine with Ability to Bind to a Model Spike Protein Leads to a Loss of SARS-CoV-2 Infectivity In Vitro When Exposed to Far-Red LED. Viruses 2021, 13, 643. [Google Scholar] [CrossRef]
- Cieplik, F.; Tabenski, L.; Buchalla, W.; Maisch, T. Antimicrobial photodynamic therapy for inactivation of biofilms formed by oral key pathogens. Front. Microbiol. 2014, 5, 405. [Google Scholar] [CrossRef]
- Svyatchenko, V.A.; Nikonov, S.D.; Mayorov, A.P.; Gelfond, M.L.; Loktev, V.B. Antiviral photodynamic therapy: Inactivation and inhibition of SARS-CoV-2 in vitro using methylene blue and Radachlorin. Photodiagn. Photodyn. Ther. 2021, 33, 102112. [Google Scholar] [CrossRef]
- Bhatti, M.; MacRobert, A.; Meghji, S.; Henderson, B.; Wilson, M. A Study of the Uptake of Toluidine Blue O by Porphyromonas gingivalis and the Mechanism of Lethal Photosensitization. Photochem. Photobiol. 1998, 68, 370–376. [Google Scholar] [CrossRef]
- Wainwright, M.; O’Kane, C.; Rawthore, S. Phenothiazinium photosensitisers XI. Improved toluidine blue photoantimicrobials. J. Photochem. Photobiol. B Biol. 2016, 160, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Fekrazad, R.; Zare, H.; Vand, S.M.S. Photodynamic therapy effect on cell growth inhibition induced by Radachlorin and toluidine blue O on Staphylococcus aureus and Escherichia coli: An in vitro study. Photodiagn. Photodyn. Ther. 2016, 15, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Pourhajibagher, M.; Kazemian, H.; Chiniforush, N.; Hosseini, N.; Pourakbari, B.; Azizollahi, A.; Rezaei, F.; Bahador, A. Exploring different photosensitizers to optimize elimination of planktonic and biofilm forms of Enterococcus faecalis from infected root canal during antimicrobial photodynamic therapy. Photodiagn. Photodyn. Ther. 2018, 24, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Kömerik, N.; Nakanishi, H.; MacRobert, A.J.; Henderson, B.; Speight, P.; Wilson, M. In Vivo Killing of Porphyromonas gingivalis by Toluidine Blue-Mediated Photosensitization in an Animal Model. Antimicrob. Agents Chemother. 2003, 47, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Najm, M.; Pourhajibagher, M.; Badirzadeh, A.; Razmjou, E.; Alipour, M.; Khoshmirsafa, M.; Bahador, A.; Hadighi, R. Photodynamic Therapy Using Toluidine Blue O (TBO) Dye as a Photosensitizer against Leishmania major. Iran. J. Public Health 2021, 50, 2111–2120. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, S.; Rosin, J.L.A.; Matuck, B.F.; Schröter, G.; Rodrigues, M.F.S.D.; Ramalho, K.M.; Raggio, D.P.; Moreira, M.S.; da Silva, L.F.F. aPDT for oral decontamination of hospitalized patients with COVID 19. Photodiagn. Photodyn. Ther. 2022, 38, 102762. [Google Scholar] [CrossRef] [PubMed]
- Sachet, P.; Rocha, B.A.; Lima, F.S.; Pedrosa, M.d.S.; Guollo, A.; Melo Filho, M.R.d.; Horta, M.C.R.; Simões, A. Management of orofacial lesions with antimicrobial photodynamic therapy and photobiomodulation protocols in patients with COVID-19: A multicenter case series. Photodiagn. Photodyn. Ther. 2022, 38, 102743. [Google Scholar] [CrossRef] [PubMed]
- ISO 10993-5:2009; Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity (ICS 11.100.20). ISO: Geneva, Switzerland, 2009.
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Chou, T.C.; Martin, N. (Eds.) CompuSyn for Drug Combinations: PC Software and User’s Guide: A Computer Program for Quantitation of Synergism and Antagonism in Drug Combinations, and the Determination of IC50and ED50 and LD50 Values; ComboSyn Inc.: Paramus, NJ, USA, 2005; p. 68. [Google Scholar]
- Zaharieva, M.M.; Trochopoulos, A.; Dimitrova, L.; Berger, M.R.; Najdenski, H.; Konstantinov, S.; Kroumov, A.D. New Insights in Routine Procedure for Mathematical Evaluation of in vitro Cytotoxicity Data from Cancer Cell Lines. Int. J. Bioautom. 2018, 22, 87–106. [Google Scholar] [CrossRef]
- Horwood, P.F.; Mahony, T.J. Multiplex real-time RT-PCR detection of three viruses associated with the bovine respiratory disease complex. J. Virol. Methods 2011, 171, 360–363. [Google Scholar] [CrossRef]
- Souf, S. Recent advances in diagnostic testing for viral infections. Biosci. Horiz. Int. J. Stud. Res. 2016, 9, hzw010. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Lei, C.; Yang, J.; Hu, J.; Sun, X. On the Calculation of TCID50 for Quantitation of Virus Infectivity. Virol. Sin. 2021, 36, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Burleson, F.G.; Chambers, T.M.; Wiedbrauk, D.L. Virology: A Laboratory Manual; Academic Press: London, UK, 1992. [Google Scholar]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Spearman, C. The method of right and wrong cases (constant stimuli) without Gauss’s formulae. Br. J. Psychol. 1908, 2, 227. [Google Scholar] [CrossRef]
- Indrayanto, G.; Putra, G.S.; Suhud, F. Chapter Six—Validation of in-vitro bioassay methods: Application in herbal drug research. In Profiles of Drug Substances, Excipients and Related Methodology; Al-Majed, A.A., Ed.; Academic Press: Cambridge, MA, USA, 2021; Volume 46, pp. 273–307. [Google Scholar]
- Teymouri, M.; Mollazadeh, S.; Mortazavi, H.; Ghale-Noie, Z.N.; Keyvani, V.; Aghababaei, F.; Hamblin, M.R.; Abbaszadeh-Goudarzi, G.; Pourghadamyari, H.; Hashemian, S.M.R.; et al. Recent advances and challenges of RT-PCR tests for the diagnosis of COVID-19. Pathol. Res. Pract. 2021, 221, 153443. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, V.; Kholina, E.; Khruschev, S.; Kovalenko, I.; Rubin, A.; Strakhovskaya, M. What Binds Cationic Photosensitizers Better: Brownian Dynamics Reveals Key Interaction Sites on Spike Proteins of SARS-CoV, MERS-CoV, and SARS-CoV-2. Viruses 2021, 13, 1615. [Google Scholar] [CrossRef] [PubMed]
- Seghatchian, J.; Struff, W.G.; Reichenberg, S. Main Properties of the THERAFLEX MB-Plasma System for Pathogen Reduction. Transfus. Med. Hemother. 2011, 38, 55–64. [Google Scholar] [CrossRef]
- Cagno, V.; Medaglia, C.; Cerny, A.; Cerny, T.; Zwygart, A.C.-A.; Cerny, E.; Tapparel, C. Methylene Blue has a potent antiviral activity against SARS-CoV-2 and H1N1 influenza virus in the absence of UV-activation in vitro. Sci. Rep. 2021, 11, 14295. [Google Scholar] [CrossRef]
- Li, Z.; Lang, Y.; Sakamuru, S.; Samrat, S.; Trudeau, N.; Kuo, L.; Rugenstein, N.; Tharappel, A.; D’Brant, L.; Koetzner, C.A.; et al. Methylene blue is a potent and broad-spectrum inhibitor against Zika virus in vitro and in vivo. Emerg. Microbes Infect. 2020, 9, 2404–2416. [Google Scholar] [CrossRef]
- Fryk, J.J.; Marks, D.C.; Hobson-Peters, J.; Prow, N.A.; Watterson, D.; Hall, R.A.; Young, P.R.; Reichenberg, S.; Sumian, C.; Faddy, H.M. Dengue and chikungunya viruses in plasma are effectively inactivated after treatment with methylene blue and visible light. Transfusion 2016, 56, 2278–2285. [Google Scholar] [CrossRef] [PubMed]
- Zhukhovitsky, V.; Shevlyagina, N.; Zubasheva, M.; Russu, L.; Gushchin, V.; Meerovich, G.; Strakhovskaya, M. Infectivity and Morphology of Bovine Coronavirus Inactivated In Vitro by Cationic Photosensitizers. Viruses 2022, 14, 1053. [Google Scholar] [CrossRef] [PubMed]
- Arentz, J.; von der Heide, H.J. Evaluation of methylene blue based photodynamic inactivation (PDI) against intracellular B-CoV and SARS-CoV2 viruses under different light sources in vitro as a basis for new local treatment strategies in the early phase of a COVID 19 infection. Photodiagn. Photodyn. Ther. 2022, 37, 102642. [Google Scholar] [CrossRef] [PubMed]
- Diederich, J.; Lodemann, E.; Wacker, A. New type of interferon inducer. Naturwissenschaften 1972, 59, 172–173. [Google Scholar] [CrossRef] [PubMed]
- Diederich, J.; Lodemann, E.; Wacker, A. Basic dyes as inducers of interferon-like activity in mice. Arch. Gesamte Virusforsch. 1973, 40, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Jamison, J.M.; Krabill, K.; Allen, K.A.; Stuart, S.H.; Tsai, C.C. RNA–Intercalating Agent Interactions: In vitro Antiviral Activity Studies. Antivir. Chem. Chemother. 1990, 1, 333–347. [Google Scholar] [CrossRef]
- Kaplan, C.; Turner, G.S. Observations on photodynamic inactivation of vaccinia virus and its effect on immunogenicity. J. Hyg. 1965, 63, 395–410. [Google Scholar] [CrossRef]
- de Oliveira, P.; Cunha, K.; Neves, P.; Muniz, M.; Gatto, G.; Salgado Filho, N.; Guedes, F.; Silva, G. Renal Morphology in Coronavirus Disease: A Literature Review. Medicina 2021, 57, 258. [Google Scholar] [CrossRef]
- Schrom, M.; Bablanian, R. Altered cellular morphology resulting from cytocidal virus infection. Arch. Virol. 1981, 70, 173–187. [Google Scholar] [CrossRef]
- Zhu, N.; Wang, W.; Liu, Z.; Liang, C.; Wang, W.; Ye, F.; Huang, B.; Zhao, L.; Wang, H.; Zhou, W.; et al. Morphogenesis and cytopathic effect of SARS-CoV-2 infection in human airway epithelial cells. Nat. Commun. 2020, 11, 3910. [Google Scholar] [CrossRef]
- Ruggieri, A.; Di Trani, L.; Gatto, I.; Franco, M.; Vignolo, E.; Bedini, B.; Elia, G.; Buonavoglia, C. Canine coronavirus induces apoptosis in cultured cells. Vet. Microbiol. 2007, 121, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Matsui, Y.; Miura, Y.; Sentsui, H. Equine coronavirus induces apoptosis in cultured cells. Vet. Microbiol. 2008, 129, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhou, H.; Huang, W.; Zhou, J.; Qiu, M.; Deng, Z.; Chen, L.; Weng, Y.; Cai, L.; Gu, Y.; et al. Cell morphological analysis of SARS-CoV-2 infection by transmission electron microscopy. J. Thorac. Dis. 2020, 12, 4368–4373. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Nyodu, R.; Maurya, V.K.; Saxena, S.K. Morphology, Genome Organization, Replication, and Pathogenesis of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). In Coronavirus Disease 2019 (COVID-19): Epidemiology, Pathogenesis, Diagnosis, and Therapeutics; Saxena, S.K., Ed.; Springer: Singapore, 2020; pp. 23–31. [Google Scholar]
- Schikora, D.; Hepburn, J.; Plavin, S. Reduction of the Viral Load by Non-Invasive Photodynamic Therapy in Early Stages of COVID-19 infection. Am. J. Virol. Dis. 2020, 2, 1–5. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 h | 24 h | 48 h | 72 h | 96 h | Doubling Time [h] | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cell Number | Abs | SD | Abs | SD | Abs | SD | Abs | SD | Abs | SD | |
| 0.15 × 106/mL | 0.381 | 0.023 | 0.514 | 0.009 | 0.778 | 0.020 | 1.280 | 0.021 | 1.818 | 0.075 | 42.4 |
| 0.125 × 106/mL | 0.337 | 0.015 | 0.443 | 0.010 | 0.617 | 0.015 | 1.200 | 0.024 | 1.594 | 0.061 | 42.6 |
| 0.1 × 106/mL | 0.277 | 0.014 | 0.385 | 0.013 | 0.532 | 0.038 | 1.129 | 0.049 | 1.518 | 0.062 | 38.9 |
| 0.08 × 106/mL | 0.232 | 0.016 | 0.325 | 0.018 | 0.447 | 0.018 | 0.986 | 0.045 | 1.356 | 0.091 | 37.5 |
| 0.06 × 106/mL | 0.209 | 0.029 | 0.275 | 0.012 | 0.372 | 0.036 | 0.835 | 0.081 | 1.200 | 0.107 | 37.9 |
| 0.04 × 106/mL | 0.145 | 0.004 | 0.196 | 0.012 | 0.296 | 0.008 | 0.615 | 0.008 | 1.055 | 0.005 | 33.4 |
| Parameters and Time of Incubation | Methylene Blue | Toluidine Blue O |
|---|---|---|
| 24 h incubation: | ||
| IC50 [µM] | 2.54 | 0.97 |
| Hill slope (m) | 3.98 | 1.88 |
| R2 | 0.99 | 0.95 |
| MNC [µM] | 2.14 | 0.66 |
| 48 h incubation: | ||
| IC50 | 1.28 | 0.52 |
| Hill slope (m) | 1.00 | 0.098 |
| R2 | 0.98 | 0.97 |
| MNC [µM] | 1.04 | 0.35 |
| 72 h incubation: | ||
| IC50 | 1.80 | 0.85 |
| Hill slope (m) | 2.52 | 0.075 |
| R2 | 0.98 | 0.96 |
| MNC [µM] | 1.1 | 0.59 |
| Sample | Concentration/20 µL * | Average Concentration/20 µL | Average Concentration of the Viral Stock/mL |
|---|---|---|---|
| C01 = dilution 10−3 | 1.07 × 105 | 1.19 × 105 | 1.06 × 108/reaction (20 µL) ↓ 2.24 × 1010/mL ** |
| C02 = dilution 10−3 | 1.31 × 105 | ||
| D01 = dilution 10−4 | 1.1 × 104 | 1.08 × 104 | |
| D02 = dilution 10−4 | 1.05 × 104 | ||
| E01 = dilution 10−5 | 1 × 103 | 0.94 × 103 | |
| E02 = dilution 10−5 | 0.88 × 103 | ||
| F01 = dilution 10−6 | 1.2 × 102 | 1.03 × 102 | |
| F02 = dilution 10−6 | 0.86 × 102 |
| EC50 [µM] (50% Viable Cells) | EC100 [µM] (100% Viable Cells) | Hill Slope | R2 | SI (IC50/EC50) | |
|---|---|---|---|---|---|
| MB | 0.018 | 0.15 | 0.900 | 0.970 | 100 |
| TBO | 0.005 | 0.02 | 1.596 | 0.982 | 170 |
| Šídák’s Multiple Comparisons Test MB vs. TBO | Mean Difference | 95.00% CI of Difference | Below Threshold? | Summary | “Adjusted p Value” |
|---|---|---|---|---|---|
| BCoV | 0.01719 | −5.908 to 5.942 | No | ns | >0.9999 |
| 0.0025 | −20.54 | −26.47 to −14.62 | Yes | **** | <0.0001 |
| 0.0050 | −35.60 | −41.53 to −29.68 | Yes | **** | <0.0001 |
| 0.0100 | −33.03 | −38.96 to −27.11 | Yes | **** | <0.0001 |
| 0.0200 | −48.96 | −54.89 to −43.04 | Yes | **** | <0.0001 |
| 0.0390 | −31.23 | −37.15 to −25.30 | Yes | **** | <0.0001 |
| 0.0750 | −36.42 | −42.35 to −30.50 | Yes | **** | <0.0001 |
| 0.1500 | −0.4070 | −6.332 to 5.518 | No | ns | >0.9999 |
| 0.3000 | −6.087 | −12.01 to −0.1619 | Yes | * | 0.0402 |
| Co | 0.01364 | −5.912 to 5.939 | No | ns | >0.9999 |
| Sample/Dilution of the cDNA | BCoV cDNA Concentration/Reaction * | BCoV cDNA Concentration/mL | Δlog cDNA/mL vs. Virus Control |
|---|---|---|---|
| Virus control: | |||
| 10−4 | 9.5 × 103 | 1.3 × 1010 | - |
| 10−5 | 8.9 × 102 | ||
| 10−6 | 9.6 × 101 | ||
| TBO 0.005 µM: | |||
| 10−3 | 3.5 × 103 | 5.8 × 108 | 2.24 × 101 |
| 10−4 | 3.9 × 102 | ||
| 10−5 | 5.1 × 101 | ||
| TBO 0.02 µM: | |||
| 10−2 | 9.2 × 102 | 1.2 × 107 | 1.08 × 103 |
| 10−3 | 8.9 × 101 | ||
| 10−4 | 6.8 × 100 | ||
| TBO 0.15 µM: | |||
| - | 4.3 × 102 | 8.2 × 104 | 1.59 × 105 |
| 10−1 | 5.2 × 101 | ||
| 10−2 | 8.2 × 100 | ||
| TBO 0.30 µM: | |||
| - | 1.4 × 101 | 2.2 × 103 | 5.91 × 106 |
| 10−1 | 2.4 × 100 | ||
| 10−2 | 1.0 × 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaharieva, M.M.; Foka, P.; Karamichali, E.; Kroumov, A.D.; Philipov, S.; Ilieva, Y.; Kim, T.C.; Podlesniy, P.; Manasiev, Y.; Kussovski, V.; et al. Photodynamic Inactivation of Bovine Coronavirus with the Photosensitizer Toluidine Blue O. Viruses 2024, 16, 48. https://doi.org/10.3390/v16010048
Zaharieva MM, Foka P, Karamichali E, Kroumov AD, Philipov S, Ilieva Y, Kim TC, Podlesniy P, Manasiev Y, Kussovski V, et al. Photodynamic Inactivation of Bovine Coronavirus with the Photosensitizer Toluidine Blue O. Viruses. 2024; 16(1):48. https://doi.org/10.3390/v16010048
Chicago/Turabian StyleZaharieva, Maya Margaritova, Pelagia Foka, Eirini Karamichali, Alexander Dimitrov Kroumov, Stanislav Philipov, Yana Ilieva, Tanya Chan Kim, Petar Podlesniy, Yordan Manasiev, Vesselin Kussovski, and et al. 2024. "Photodynamic Inactivation of Bovine Coronavirus with the Photosensitizer Toluidine Blue O" Viruses 16, no. 1: 48. https://doi.org/10.3390/v16010048