
Sequencing, Analysis and Organization of the Complete Genome of a Novel Baculovirus Calliteara abietis Nucleopolyhedrovirus (CaabNPV)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Baculovirus Isolation and Host Identification
2.2. Electron Microscopy
2.3. Viral DNA Isolation, Sequencing, and Assembly
2.4. Genome Sequence Analysis
2.5. Phylogeny and Kimura 2-Parameter Analysis
3. Results and Discussion
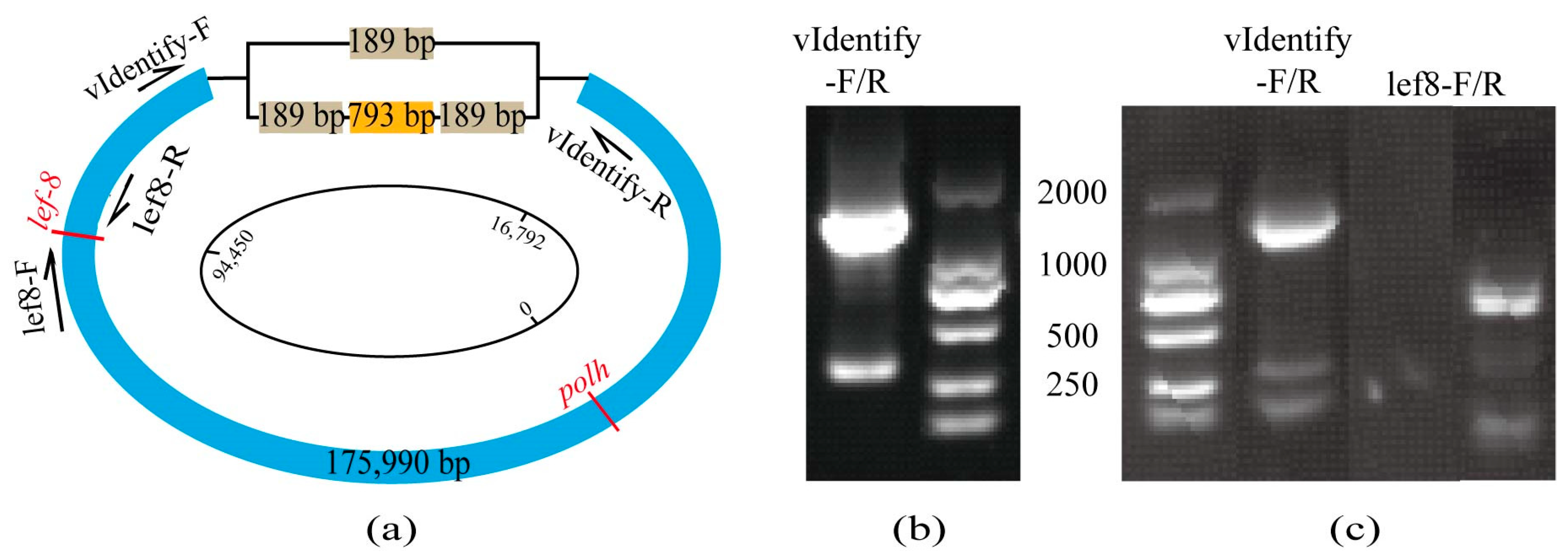
3.1. Host Determination and Virus Characterization
3.2. Nucleotide Sequence Analysis
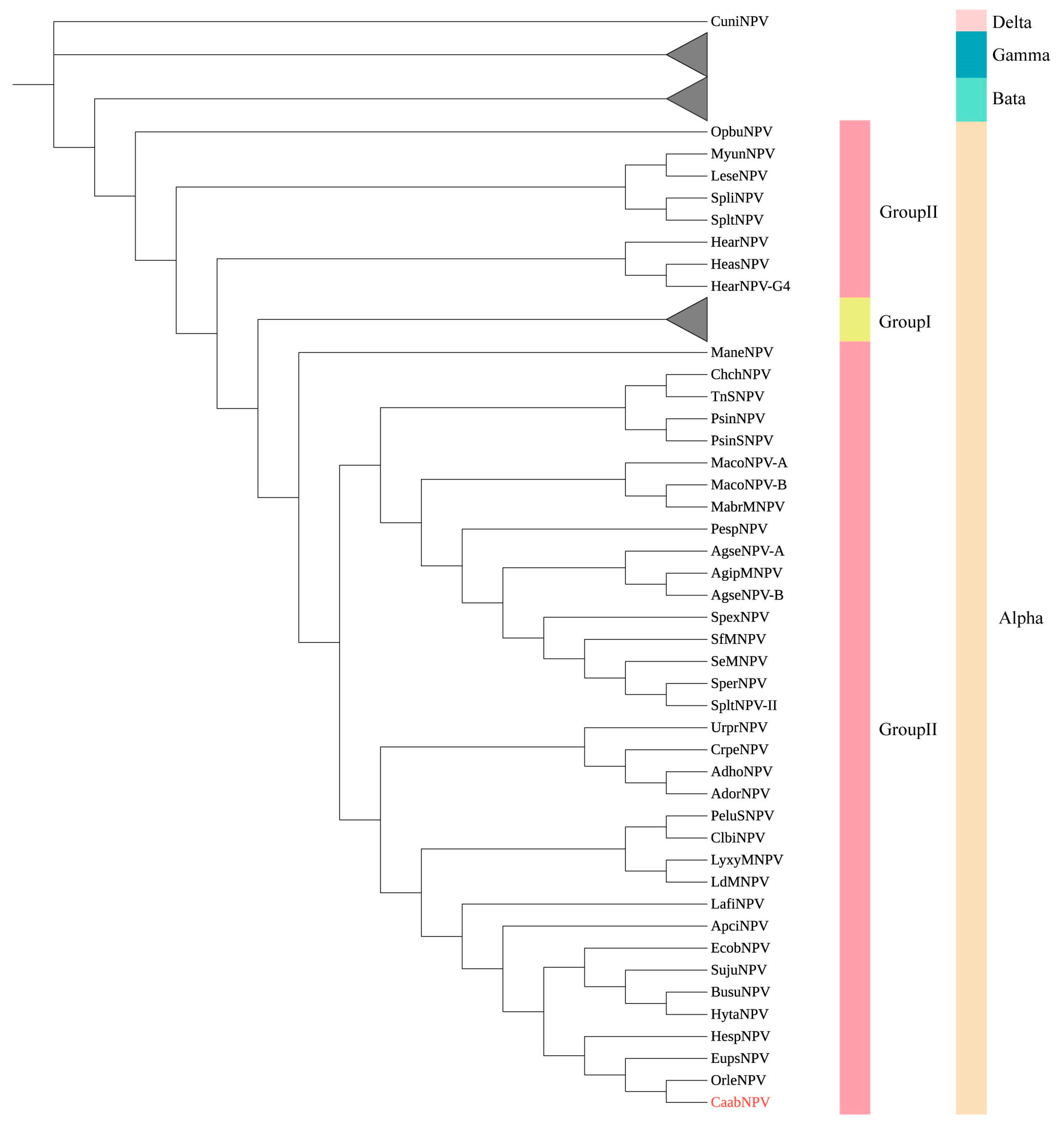
3.3. Phylogenetic Analysis
3.4. Repeated Sequences
3.5. Analysis of CaabNPV Multi-Copy Genes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
References
- Ribeiro, B.M.; Dos Santos, E.R.; Trentin, L.B.; da Silva, L.A.; de Melo, F.L.; Kitajima, E.W.; Ardisson-Araújo, D.M.P. A Nymphalid-Infecting Group I Alphabaculovirus Isolated from the Major Passion Fruit Caterpillar Pest Dione Juno Juno (Lepidoptera: Nymphalidae). Viruses 2019, 11, 602. [Google Scholar] [CrossRef]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus Insecticides in Latin America: Historical Overview, Current Status and Future Perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef]
- Harrison, R.L.; Herniou, E.A.; Jehle, J.A.; Theilmann, D.A.; Burand, J.P.; Becnel, J.J.; Krell, P.J.; van Oers, M.M.; Mowery, J.D.; Bauchan, G.R.; et al. ICTV Virus Taxonomy Profile: Baculoviridae. J. Gen. Virol. 2018, 99, 1185–1186. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, G.F. Baculovirus Molecular Biology, 4th ed.; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2019. [Google Scholar]
- Ackermann, H.-W.; Smirnoff, W.A. A Morphological Investigation of 23 Baculoviruses. J. Invertebr. Pathol. 1983, 41, 269–280. [Google Scholar] [CrossRef]
- Blissard, G.W.; Theilmann, D.A. Baculovirus Entry and Egress from Insect Cells. Annu. Rev. Virol. 2018, 5, 113–139. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, T.; Sako, Y. A Role for the Anti-Viral Host Defense Mechanism in the Phylogenetic Divergence in Baculovirus Evolution. PLoS ONE 2016, 11, e0156394. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.; Thickless, R.L. Baculovirus Molecular Evolution via Gene Turnover and Recurrent Positive Selection of Key Genes. J. Virol. 2017, 91, e01319-17. [Google Scholar] [CrossRef] [PubMed]
- Thézé, J.; Lopez-Vaamonde, C.; Cory, J.S.; Herniou, E.A. Biodiversity, Evolution and Ecological Specialization of Baculoviruses: A Treasure Trove for Future Applied Research. Viruses 2018, 10, 366. [Google Scholar] [CrossRef] [PubMed]
- Gninenko, Y.I.; Serdyukov, G.V. The Coniferous Tussock Moth (Calliteara abietis, Lepidoptera, Lymantriidae) in Buryatia. Entmol. Rev. 2010, 90, 999–1002. [Google Scholar] [CrossRef]
- Shu, R.; Meng, Q.; Miao, L.; Liang, H.; Chen, J.; Xu, Y.; Cheng, L.; Jin, W.; Qin, Q.; Zhang, H. Genome Analysis of a Novel Clade b Betabaculovirus Isolated from the Legume Pest Matsumuraeses phaseoli (Lepidoptera: Tortricidae). Viruses 2020, 12, 1068. [Google Scholar] [CrossRef]
- Stewart, D.; Zahiri, R.; Djoumad, A.; Freschi, L.; Lamarche, J.; Holden, D.; Cervantes, S.; Ojeda, D.I.; Potvin, A.; Nisole, A.; et al. A Multi-Species TaqMan PCR Assay for the Identification of Asian Gypsy Moths (Lymantria spp.) and Other Invasive Lymantriines of Biosecurity Concern to North America. PLoS ONE 2016, 11, e0160878. [Google Scholar] [CrossRef]
- Zhang, H.; Tong, Y.; Liu, H.; Guo, L.; Jin, W.; Li, X.; Meng, Q.; Yu, X.; Fang, F.; Qin, Q.; et al. Establishment of a Rapid Detection Method for Plutella Xylostella Granulovirus Based on qPCR. Heliyon 2023, 9, e15170. [Google Scholar] [CrossRef]
- Denman, A.M. Molecular Cloning: A Laboratory Manual. Immunology 1983, 49, 411. [Google Scholar]
- Solovyev, V.; Kosarev, P.; Seledsov, I.; Vorobyev, D. Automatic Annotation of Eukaryotic Genes, Pseudogenes and Promoters. Genome Biol. 2006, 7, S10. [Google Scholar] [CrossRef]
- van Oers, M.M.; Vlak, J.M. Baculovirus genomics. Curr. Drug Targets 2007, 8, 1051–1068. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.H.; Arif, B.M.; Jin, F.; Martens, J.W.; Chen, X.W.; Sun, J.S.; Zuidema, D.; Goldbach, R.W.; Vlak, J.M. Distinct Gene Arrangement in the Buzura Suppressaria Single-Nucleocapsid Nucleopolyhedrovirus Genome. J. Gen. Virol. 1998, 79 Pt 11, 2841–2851. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and Sequence Analysis Tools Services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem Repeats Finder: A Program to Analyze DNA Sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xie, Y.; Ma, J.; Luo, X.; Nie, P.; Zuo, Z.; Lahrmann, U.; Zhao, Q.; Zheng, Y.; Zhao, Y.; et al. IBS: An Illustrator for the Presentation and Visualization of Biological Sequences. Bioinformatics 2015, 31, 3359–3361. [Google Scholar] [CrossRef] [PubMed]
- Guarino, L.A.; Smith, M. Regulation of Delayed-Early Gene Transcription by Dual TATA Boxes. J. Virol. 1992, 66, 3733–3739. [Google Scholar] [CrossRef]
- Blissard, G.W.; Kogan, P.H.; Wei, R.; Rohrmann, G.F. A Synthetic Early Promoter from a Baculovirus: Roles of the TATA Box and Conserved Start Site CAGT Sequence in Basal Levels of Transcription. Virology 1992, 190, 783–793. [Google Scholar] [CrossRef]
- Wormleaton, S.; Kuzio, J.; Winstanley, D. The Complete Sequence of the Adoxophyes Orana Granulovirus Genome. Virology 2003, 311, 350–365. [Google Scholar] [CrossRef]
- Morris, T.D.; Miller, L.K. Mutational Analysis of a Baculovirus Major Late Promoter. Gene 1994, 140, 147–153. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic Orthology Inference for Comparative Genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A Fast, Scalable and User-Friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Jehle, J.A.; Lange, M.; Wang, H.; Hu, Z.; Wang, Y.; Hauschild, R. Molecular Identification and Phylogenetic Analysis of Baculoviruses from Lepidoptera. Virology 2006, 346, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Friesen, P.D.; Nissen, M.S. Gene Organization and Transcription of TED, a Lepidopteran Retrotransposon Integrated within the Baculovirus Genome. Mol. Cell Biol. 1990, 10, 3067–3077. [Google Scholar] [CrossRef] [PubMed]
- Beames, B.; Summers, M.D. Sequence Comparison of Cellular and Viral Copies of Host Cell DNA Insertions Found in Autographa Californica Nuclear Polyhedrosis Virus. Virology 1990, 174, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, G.; Williams, T.; Muñoz, D.; Caballero, P.; López-Ferber, M. Mixed Genotype Transmission Bodies and Virions Contribute to the Maintenance of Diversity in an Insect Virus. Proc. R. Soc. B Biol. Sci. 2010, 277, 943–951. [Google Scholar] [CrossRef]
- Miele, S.A.B.; Garavaglia, M.J.; Belaich, M.N.; Ghiringhelli, P.D. Baculovirus: Molecular Insights on Their Diversity and Conservation. Int. J. Evol. Biol. 2011, 2011, 379424. [Google Scholar] [CrossRef] [PubMed]
- Thumbi, D.K.; Eveleigh, R.J.M.; Lucarotti, C.J.; Lapointe, R.; Graham, R.I.; Pavlik, L.; Lauzon, H.A.M.; Arif, B.M. Complete Sequence, Analysis and Organization of the Orgyia Leucostigma Nucleopolyhedrovirus Genome. Viruses 2011, 3, 2301–2327. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, M.J.; Miele, S.A.B.; Iserte, J.A.; Belaich, M.N.; Ghiringhelli, P.D. The Ac53, Ac78, Ac101, and Ac103 Genes Are Newly Discovered Core Genes in the Family Baculoviridae. J. Virol. 2012, 86, 12069–12079. [Google Scholar] [CrossRef]
- Ardisson-Araújo, D.M.P.; da Silva, A.M.R.; Melo, F.L.; Dos Santos, E.R.; Sosa-Gómez, D.R.; Ribeiro, B.M. A Novel Betabaculovirus Isolated from the Monocot Pest Mocis latipes (Lepidoptera: Noctuidae) and the Evolution of Multiple-Copy Genes. Viruses 2018, 10, 134. [Google Scholar] [CrossRef] [PubMed]
- Guarino, L.A.; Gonzalez, M.A.; Summers, M.D. Complete Sequence and Enhancer Function of the Homologous DNA Regions of Autographa Californica Nuclear Polyhedrosis Virus. J. Virol. 1986, 60, 224–229. [Google Scholar] [CrossRef]
- Kang, W.; Suzuki, M.; Zemskov, E.; Okano, K.; Maeda, S. Characterization of Baculovirus Repeated Open Reading Frames (Bro) in Bombyx Mori Nucleopolyhedrovirus. J. Virol. 1999, 73, 10339–10345. [Google Scholar] [CrossRef]
- Kang, W.K.; Imai, N.; Suzuki, M.; Iwanaga, M.; Matsumoto, S.; Zemskov, E.A. Interaction of Bombyx Mori Nucleopolyhedrovirus BRO-A and Host Cell Protein Laminin. Arch. Virol. 2003, 148, 99–113. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Z.; Liu, C.; Qu, L.; Wang, D. Genome Analysis of an Alphabaculovirus Isolated from the Larch Looper, Erannis ankeraria. Viruses 2021, 14, 34. [Google Scholar] [CrossRef] [PubMed]
- Mikhailov, V.S.; Vanarsdall, A.L.; Rohrmann, G.F. Isolation and Characterization of the DNA-Binding Protein (DBP) of the Autographa Californica Multiple Nucleopolyhedrovirus. Virology 2008, 370, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, W.; Byambasuren, M.; Ganbold, U.; Shi, H.; Liang, H.; Li, M.; Wang, H.; Qin, Q.; Zhang, H. Sequencing, Analysis and Organization of the Complete Genome of a Novel Baculovirus Calliteara abietis Nucleopolyhedrovirus (CaabNPV). Viruses 2024, 16, 252. https://doi.org/10.3390/v16020252
Jin W, Byambasuren M, Ganbold U, Shi H, Liang H, Li M, Wang H, Qin Q, Zhang H. Sequencing, Analysis and Organization of the Complete Genome of a Novel Baculovirus Calliteara abietis Nucleopolyhedrovirus (CaabNPV). Viruses. 2024; 16(2):252. https://doi.org/10.3390/v16020252
Chicago/Turabian StyleJin, Wenyi, Mijidsuren Byambasuren, Uranbileg Ganbold, Huixian Shi, Hongbin Liang, Miaomiao Li, Hongtuo Wang, Qilian Qin, and Huan Zhang. 2024. "Sequencing, Analysis and Organization of the Complete Genome of a Novel Baculovirus Calliteara abietis Nucleopolyhedrovirus (CaabNPV)" Viruses 16, no. 2: 252. https://doi.org/10.3390/v16020252
APA StyleJin, W., Byambasuren, M., Ganbold, U., Shi, H., Liang, H., Li, M., Wang, H., Qin, Q., & Zhang, H. (2024). Sequencing, Analysis and Organization of the Complete Genome of a Novel Baculovirus Calliteara abietis Nucleopolyhedrovirus (CaabNPV). Viruses, 16(2), 252. https://doi.org/10.3390/v16020252