
Development of Colloidal Gold-Based Immunochromatographic Strips for Rapid Detection and Surveillance of Japanese Encephalitis Virus in Dogs across Shanghai, China
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus and Serum Samples
2.2. Cloning, Expression, and Purification of Recombinant JEV EDIII Protein
2.3. Preparation of the Colloidal Gold-Labeled Suspensions
2.4. Preparation of the Immunochromatographic Strip
2.5. Working Principle of Immunochromatographic Strip (ICS)
2.6. Specificity, Sensitivity, and Stability of the ICS
2.7. Seroprevalence of JEV among Dogs in Shanghai, China
2.8. Statistical Analysis
3. Results
3.1. Expression and Purification of the Recombinant ED3 Protein
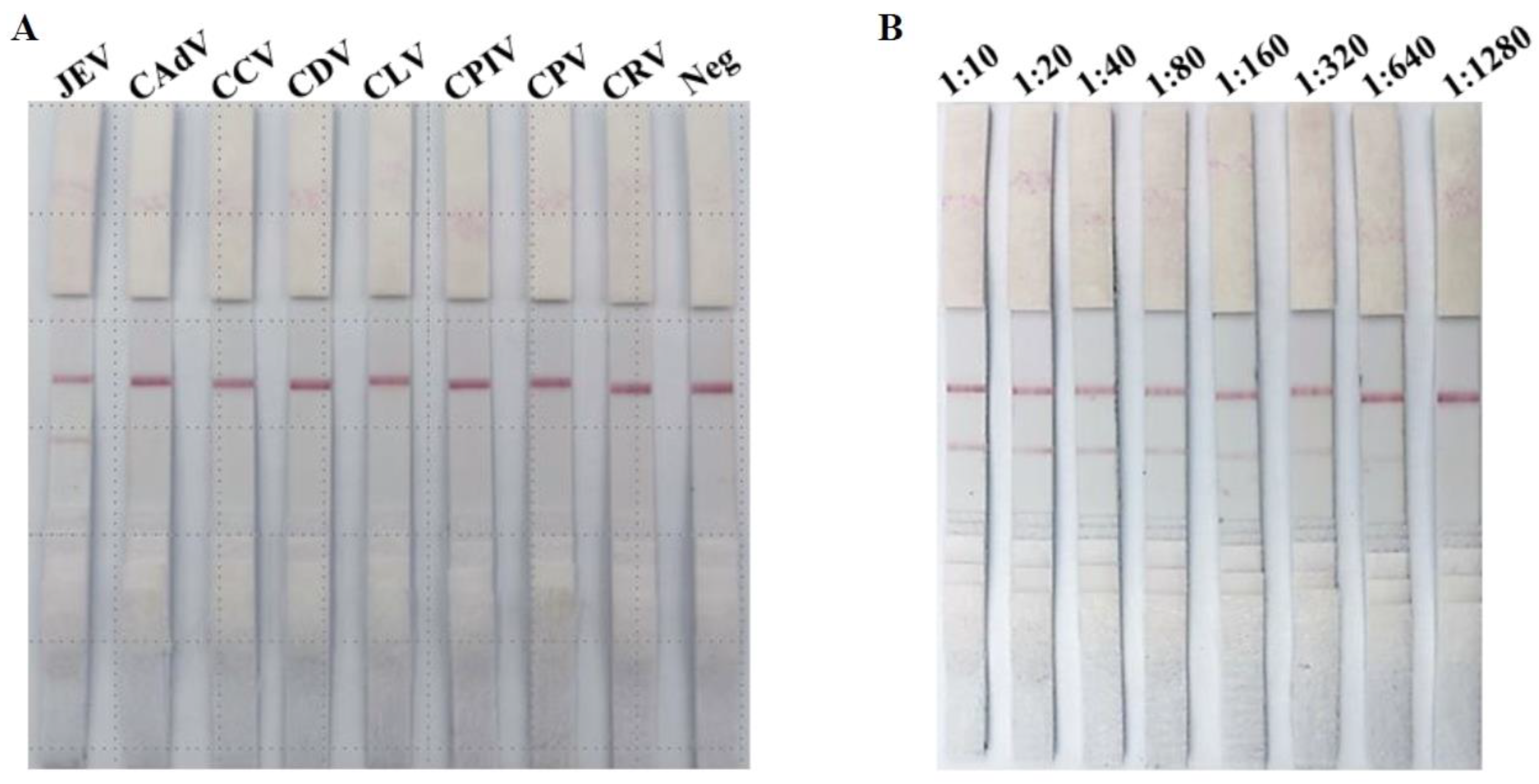
3.2. Specificity Evaluation of ICS
3.3. Sensitivity Evaluation of ICS
3.4. Stability Evaluation of ICS
3.5. Surveillance of JEV Antibodies in Dogs in Shanghai
3.6. Relationship between Dog JEV-Antibody-Positive Rate and Season in Shanghai
3.7. Relationship between Dog JEV-Antibody-Positive Rate and Living Environment in Shanghai
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774E. [Google Scholar] [CrossRef] [PubMed]
- Tarantola, A.; Goutard, F.; Newton, P.; de Lamballerie, X.; Lortholary, O.; Cappelle, J.; Buchy, P. Estimating the burden of Japanese encephalitis virus and other encephalitides in countries of the mekong region. PLoS Neglected Trop. Dis. 2014, 8, e2533. [Google Scholar] [CrossRef] [PubMed]
- Erlanger, T.E.; Weiss, S.; Keiser, J.; Utzinger, J.; Wiedenmayer, K. Past, present, and future of Japanese encephalitis. Emerg. Infect. Dis. 2009, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Turtle, L.; Solomon, T. Japanese encephalitis—The prospects for new treatments. Nat. Rev. Neurol. 2018, 14, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Wahaab, A.; Mustafa, B.E.; Hameed, M.; Stevenson, N.J.; Anwar, M.N.; Liu, K.; Wei, J.; Qiu, Y.; Ma, Z. Potential Role of Flavivirus NS2B-NS3 Proteases in Viral Pathogenesis and Anti-flavivirus Drug Discovery Employing Animal Cells and Models: A Review. Viruses 2021, 14, 44. [Google Scholar] [CrossRef] [PubMed]
- Brand, C.; Bisaillon, M.; Geiss, B.J. Organization of the Flavivirus RNA replicase complex. Wiley Interdiscip. Rev. RNA 2017, 8, e1437. [Google Scholar] [CrossRef] [PubMed]
- Wahaab, A.; Liu, K.; Hameed, M.; Anwar, M.N.; Kang, L.; Li, C.; Ma, X.; Wajid, A.; Yang, Y.; Khan, U.H.; et al. Identification of Cleavage Sites Proteolytically Processed by NS2B-NS3 Protease in Polyprotein of Japanese Encephalitis Virus. Pathogens 2021, 10, 102. [Google Scholar] [CrossRef]
- Wahaab, A.; Zhang, Y.; Rasgon, J.L.; Kang, L.; Hameed, M.; Li, C.; Anwar, M.N.; Zhang, Y.; Shoaib, A.; Liu, K.; et al. NS2B-D55E and NS2B-E65D Variations are Responsible for Differences in NS2B-NS3 Protease Activities Between Japanese Encephalitis Virus Genotype I and III in Fluorogenic Peptide Model. bioRxiv 2023. bioRxiv:2023.12.08.570834. [Google Scholar]
- Gao, X.; Liu, H.; Li, X.; Fu, S.; Cao, L.; Shao, N.; Zhang, W.; Wang, Q.; Lu, Z.; Lei, W.; et al. Changing Geographic Distribution of Japanese Encephalitis Virus Genotypes, 1935–2017. Vector Borne Zoonotic Dis. 2019, 19, 35–44. [Google Scholar] [CrossRef]
- Hameed, M.; Liu, K.; Anwar, M.N.; Wahaab, A.; Safdar, A.; Di, D.; Boruah, P.; Xu, J.; Wang, X.; Li, B.; et al. The emerged genotype I of Japanese encephalitis virus shows an infectivity similar to genotype III in Culex pipiens mosquitoes from China. PLoS Neglected Trop. Dis. 2019, 13, e0007716. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Li, K.; Mehmood, K.; Gui, R.; Li, J. Epidemiology of Japanese Encephalitis in China (2004–2015). Travel Med. Infect. Dis. 2019, 28, 109–110. [Google Scholar] [CrossRef]
- Endy, T.; Nisalak, A. Japanese encephalitis virus: Ecology and epidemiology. In Japanese Encephalitis and West Nile Viruses; Springer: Berlin/Heidelberg, Germany, 2002; pp. 11–48. [Google Scholar]
- Dye, C. The analysis of parasite transmission by bloodsucking insects. Annu. Rev. Entomol. 1992, 37, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.C.; Dhanze, H.; Bhilegaonkar, K.; Chakurkar, E.; Kumar, A.; Yathish, H. Serological evidence of Japanese encephalitis virus infection in pigs in a low human incidence state, Goa, India. Prev. Vet. Med. 2020, 175, 104882. [Google Scholar] [CrossRef] [PubMed]
- Ladreyt, H.; Auerswald, H.; Tum, S.; Ken, S.; Heng, L.; In, S.; Lay, S.; Top, C.; Ly, S.; Duong, V.; et al. Comparison of Japanese Encephalitis Force of Infection in Pigs, Poultry and Dogs in Cambodian Villages. Pathogens 2020, 9, 719. [Google Scholar] [CrossRef] [PubMed]
- Widiastuti, D.; Wijayanti, T.; Isnani, T.; Pramestuti, N.; Wijayanti, S.M.P. Seroprevalence of Japanese enchephalitis Infection in Pigs in Tulungagung, East Java. Balaba J. Litbang Pengendali. Penyakit Bersumber Binatang Banjarnegara 2019, 15, 125–132. [Google Scholar] [CrossRef]
- Ohno, Y.; Sato, H.; Suzuki, K.; Yokoyama, M.; Uni, S.; Shibasaki, T.; Sashika, M.; Inokuma, H.; Kai, K.; Maeda, K. Detection of antibodies against Japanese encephalitis virus in raccoons, raccoon dogs and wild boars in Japan. J. Vet. Med. Sci. 2009, 71, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.; Wang, Q.; Cao, S.; Zhao, Q.; Wen, Y.; Huang, X.; Wen, X.; Yan, Q.; Ma, X.; Wu, R. Serological and molecular epidemiology of Japanese encephalitis virus infections in swine herds in China, 2006–2012. J. Vet. Sci. 2018, 19, 151. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, H.; Ohno, Y.; Mochizuki, M.; Iwata, H.; Okuda, M.; Maeda, K. Dogs as sentinels for human infection with Japanese encephalitis virus. Emerg. Infect. Dis. 2010, 16, 1137–1139. [Google Scholar] [CrossRef]
- Shimoda, H.; Tamaru, S.; Morimoto, M.; Hayashi, T.; Shimojima, M.; Maeda, K. Experimental infection of Japanese encephalitis virus in dogs. J. Vet. Med. Sci. 2011, 73, 1241–1242. [Google Scholar] [CrossRef][Green Version]
- Anderson, K.B.; Gibbons, R.V.; Thomas, S.J.; Rothman, A.L.; Nisalak, A.; Berkelman, R.L.; Libraty, D.H.; Endy, T.P. Preexisting Japanese encephalitis virus neutralizing antibodies and increased symptomatic dengue illness in a school-based cohort in Thailand. PLoS Neglected Trop. Dis. 2011, 5, e1311. [Google Scholar] [CrossRef]
- Chiou, S.S.; Tsai, K.H.; Huang, C.G.; Liao, Y.K.; Chen, W.J. High antibody prevalence in an unconventional ecosystem is related to circulation of a low-virulent strain of Japanese encephalitis virus. Vaccine 2007, 25, 1437–1443. [Google Scholar] [CrossRef]
- Ting, S.H.; Tan, H.C.; Wong, W.K.; Ng, M.L.; Chan, S.H.; Ooi, E.E. Seroepidemiology of neutralizing antibodies to Japanese encephalitis virus in Singapore: Continued transmission despite abolishment of pig farming? Acta Trop. 2004, 92, 187–191. [Google Scholar] [CrossRef]
- Yang, D.K.; Kim, B.H.; Lim, S.I.; Kwon, J.H.; Lee, K.W.; Choi, C.U.; Kweon, C.H. Development and evaluation of indirect ELISA for the detection of antibodies against Japanese encephalitis virus in swine. J. Vet. Sci. 2006, 7, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Cha, G.W.; Cho, J.E.; Ju, Y.R.; Hong, Y.-J.; Han, M.G.; Lee, W.-J.; Choi, E.Y.; Jeong, Y.E. Comparison of four serological tests for detecting antibodies to Japanese encephalitis virus after vaccination in children. Osong Public Health Res. Perspect. 2014, 5, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Counor, D.; Shen, D.; Sun, G.; He, H.; Deubel, V.; Zhang, S. Detection of Japanese encephalitis virus antibodies in bats in Southern China. Am. J. Trop. Med. Hyg. 2008, 78, 1007–1011. [Google Scholar] [CrossRef]
- Hamano, M.; Lim, C.K.; Takagi, H.; Sawabe, K.; Kuwayama, M.; Kishi, N.; Kurane, I.; Takasaki, T. Detection of antibodies to Japanese encephalitis virus in the wild boars in Hiroshima prefecture, Japan. Epidemiol. Infect. 2007, 135, 974–977. [Google Scholar] [CrossRef]
- Xinglin, J.; Huanchun, C.; Xiang, W.; Changming, Q. Quantitative and qualitative study of enzyme-linked immunosorbent assay to detect IgG against Japanese encephalitis virus in swine sera. Vet. Res. Commun. 2005, 29, 159–169. [Google Scholar] [CrossRef]
- Shimoda, H.; Inthong, N.; Noguchi, K.; Terada, Y.; Nagao, Y.; Shimojima, M.; Takasaki, T.; Rerkamnuaychoke, W.; Maeda, K. Development and application of an indirect enzyme-linked immunosorbent assay for serological survey of Japanese encephalitis virus infection in dogs. J. Virol. Methods 2013, 187, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Koraka, P.; Zeller, H.; Niedrig, M.; Osterhaus, A.D.; Groen, J. Reactivity of serum samples from patients with a flavivirus infection measured by immunofluorescence assay and ELISA. Microbes Infect. 2002, 4, 1209–1215. [Google Scholar] [CrossRef]
- Litzba, N.; Klade, C.S.; Lederer, S.; Niedrig, M. Evaluation of serological diagnostic test systems assessing the immune response to Japanese encephalitis vaccination. PLoS Neglected Trop. Dis. 2010, 4, e883. [Google Scholar] [CrossRef]
- Niedrig, M.; Kürsteiner, O.; Herzog, C.; Sonnenberg, K. Evaluation of an indirect immunofluorescence assay for detection of immunoglobulin M (IgM) and IgG antibodies against yellow fever virus. Clin. Vaccine Immunol. CVI 2008, 15, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Feng, S.; Li, Y.; He, Y.; Jin, X.; Wang, X.; Zhou, Z.; Xiao, Y.; Bi, D. Development of immunochromatographic test strips for rapid, quantitative detection of H9AIV antibodies. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1095, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Li, X.S.; Fu, F.; Lang, Y.K.; Li, H.Z.; Wang, W.; Chen, X.; Tian, H.B.; Zhou, Y.J.; Tong, G.Z.; Li, X. Development and preliminary application of an immunochromatographic strip for rapid detection of infection with porcine reproductive and respiratory syndrome virus in swine. J. Virol. Methods 2011, 176, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Yang, J.; Lu, Q.; Guo, J.; Deng, R.; Wang, Y.; Wang, S.; Wang, S.; Chen, W.; Zhi, Y.; et al. Development of an immunochromatographic strip for the detection of antibodies against Porcine circovirus-2. J. Vet. Diagn. Investig. 2012, 24, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Cha, G.-W.; Lee, E.J.; Lim, E.-J.; Sin, K.S.; Park, W.W.; Jeon, D.Y.; Han, M.G.; Lee, W.-J.; Choi, W.-Y.; Jeong, Y.E. A novel immunochromatographic test applied to a serological survey of Japanese encephalitis virus on pig farms in Korea. PLoS ONE 2015, 10, e0127313. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Andres, J.; Rani, S.; Varjak, M.; Chase-Topping, M.E.; Beck, M.H.; Ferguson, M.C.; Schnettler, E.; Fragkoudis, R.; Barry, G.; Merits, A. Phenoloxidase activity acts as a mosquito innate immune response against infection with Semliki Forest virus. PLoS Pathog. 2012, 8, e1002977. [Google Scholar] [CrossRef]
- Sun, Q.; Xie, Y.; Guan, Z.; Zhang, Y.; Li, Y.; Yang, Y.; Zhang, J.; Li, Z.; Qiu, Y.; Li, B.; et al. Seroprevalence of Getah virus in Pigs in Eastern China Determined with a Recombinant E2 Protein-Based Indirect ELISA. Viruses 2022, 14, 2173. [Google Scholar] [CrossRef]
- Zhu, Z.; Shi, Z.; Yan, W.; Wei, J.; Shao, D.; Deng, X.; Wang, S.; Li, B.; Tong, G.; Ma, Z. Nonstructural protein 1 of influenza A virus interacts with human guanylate-binding protein 1 to antagonize antiviral activity. PLoS ONE 2013, 8, e55920. [Google Scholar] [CrossRef]
- Li, Y.; Hou, L.; Ye, J.; Liu, X.; Dan, H.; Jin, M.; Chen, H.; Cao, S. Development of a convenient immunochromatographic strip for the diagnosis of infection with Japanese encephalitis virus in swine. J. Virol. Methods 2010, 168, 51–56. [Google Scholar] [CrossRef]
- Xiang, L.; Li, L. Development and evaluation of an immunochromatographic strip for the detection of Human cytomegalovirus. Lett. Appl. Microbiol. 2011, 52, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Zhang, J.; Wang, G.; Chen, W.; Chen, L.; Zhang, X. Preparation of colloidal gold immunochromatographic strip for detection of Paragonimiasis skrjabini. PLoS ONE 2014, 9, e92034. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Maeki, T.; Tajima, S.; Kyaw, A.K.; Matsumoto, F.; Miura, K.; Yamashita, A.; Yoshikawa, A.; Negishi, K.; Noguchi, Y.; Tadokoro, K.; et al. Comparison of Neutralizing Antibody Titers against Japanese Encephalitis Virus Genotype V Strain with Those against Genotype I and III Strains in the Sera of Japanese Encephalitis Patients in Japan in 2016. Jpn. J. Infect. Dis. 2018, 71, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Chávez, J.H.; Silva, J.R.; Amarilla, A.A.; Moraes Figueiredo, L.T. Domain III peptides from flavivirus envelope protein are useful antigens for serologic diagnosis and targets for immunization. Biol. J. Int. Assoc. Biol. Stand. 2010, 38, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Philbert, A.; Ijumba, J.N. Preferred breeding habitats of Aedes aegypti (Diptera Culicidae) mosquito and its public health implications in Dares Salaam. J. Environ. Res. Manag. 2013, 4, 344–351. [Google Scholar]
- Tian, H.-Y.; Bi, P.; Cazelles, B.; Zhou, S.; Huang, S.-Q.; Yang, J.; Pei, Y.; Wu, X.-X.; Fu, S.-H.; Tong, S.-L. How environmental conditions impact mosquito ecology and Japanese encephalitis: An eco-epidemiological approach. Environ. Int. 2015, 79, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Hameed, M.; Khan, S.; Xu, J.; Zhang, J.; Wang, X.; Di, D.; Chen, Z.; Naveed Anwar, M.; Wahaab, A.; Ma, X.; et al. Detection of Japanese encephalitis virus in mosquitoes from Xinjiang during next-generation sequencing arboviral surveillance. Transbound. Emerg. Dis. 2021, 68, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Solomon, T. Pathogenic flaviviruses. Lancet 2008, 371, 500–509. [Google Scholar] [CrossRef]
- Huang, Y.-J.S.; Hettenbach, S.M.; Park, S.L.; Higgs, S.; Barrett, A.D.; Hsu, W.-W.; Harbin, J.N.; Cohnstaedt, L.W.; Vanlandingham, D.L. Differential infectivities among different Japanese encephalitis virus genotypes in Culex quinquefasciatus mosquitoes. PLoS Neglected Trop. Dis. 2016, 10, e0005038. [Google Scholar] [CrossRef]
- De Wispelaere, M.; Desprès, P.; Choumet, V. European Aedes albopictus and Culex pipiens are competent vectors for Japanese encephalitis virus. PLoS Neglected Trop. Dis. 2017, 11, e0005294. [Google Scholar] [CrossRef]
- Hameed, M.; Wahaab, A.; Nawaz, M.; Khan, S.; Nazir, J.; Liu, K.; Wei, J.; Ma, Z. Potential Role of Birds in Japanese Encephalitis Virus Zoonotic Transmission and Genotype Shift. Viruses 2021, 13, 357. [Google Scholar] [CrossRef]
- Liu, W.; Fu, S.; Ma, X.; Chen, X.; Wu, D.; Zhou, L.; Yin, Q.; Li, F.; He, Y.; Lei, W.; et al. An outbreak of Japanese encephalitis caused by genotype Ib Japanese encephalitis virus in China, 2018: A laboratory and field investigation. PLoS Neglected Trop. Dis. 2020, 14, e0008312. [Google Scholar] [CrossRef]
- Deng, X.; Yan, J.Y.; He, H.Q.; Yan, R.; Sun, Y.; Tang, X.W.; Zhou, Y.; Pan, J.H.; Mao, H.Y.; Zhang, Y.J.; et al. Serological and molecular epidemiology of Japanese Encephalitis in Zhejiang, China, 2015–2018. PLoS Neglected Trop. Dis. 2020, 14, e0008574. [Google Scholar] [CrossRef]
- Auerswald, H.; Ruget, A.S.; Ladreyt, H.; In, S.; Mao, S.; Sorn, S.; Tum, S.; Duong, V.; Dussart, P.; Cappelle, J.; et al. Serological Evidence for Japanese Encephalitis and West Nile Virus Infections in Domestic Birds in Cambodia. Front. Vet. Sci. 2020, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Arshad, S.S.; Selvarajah, G.T.; Abu, J.; Toung, O.P.; Abba, Y.; Bande, F.; Yasmin, A.R.; Sharma, R.; Ong, B.L.; et al. Prevalence and risk factors of Japanese encephalitis virus (JEV) in livestock and companion animal in high-risk areas in Malaysia. Trop. Anim. Health Prod. 2018, 50, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Pham-Thanh, L.; Nguyen-Tien, T.; Magnusson, U.; Bui-Nghia, V.; Bui-Ngoc, A.; Le-Thanh, D.; Lundkvist, Å.; Can-Xuan, M.; Nguyen-Thi Thu, T.; Vu-Thi Bich, H. Dogs as Sentinels for Flavivirus Exposure in Urban, Peri-Urban and Rural Hanoi, Vietnam. Viruses 2021, 13, 507. [Google Scholar] [CrossRef]
- Grant, D.C.; Troy, G.C. 69—Canine Infectious Diseases. In Canine Internal Medicine Secrets; Rubin, S.I., Carr, A.P., Eds.; Mosby: Saint Louis, MO, USA, 2007; pp. 394–422. [Google Scholar] [CrossRef]
- Wu, K.-P.; Wu, C.-W.; Tsao, Y.-P.; Kuo, T.-W.; Lou, Y.-C.; Lin, C.-W.; Wu, S.-C.; Cheng, J.-W. Structural basis of a flavivirus recognized by its neutralizing antibody: Solution structure of the domain III of the Japanese encephalitis virus envelope protein. J. Biol. Chem. 2003, 278, 46007–46013. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Hernández-Triana, L.M.; Banyard, A.C.; Fooks, A.R.; Johnson, N. Japanese encephalitis virus infection, diagnosis and control in domestic animals. Vet. Microbiol. 2017, 201, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wen, Y.; An, T.; Duan, G.; Sun, M.; Ge, J.; Li, X.; Yang, K.; Cai, X. Development of an Immunochromatographic Strip for Rapid Detection of Canine Adenovirus. Front. Microbiol. 2019, 10, 2882. [Google Scholar] [CrossRef]
- Sharma, C.; Singh, M.; Upmanyu, V.; Chander, V.; Verma, S.; Chakrovarty, S.; Sharma, G.K.; Dhanze, H.; Singh, P.; Shrivastava, S.; et al. Development and evaluation of a gold nanoparticle-based immunochromatographic strip test for the detection of canine parvovirus. Arch. Virol. 2018, 163, 2359–2368. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, Y.; Zhou, Z.-B.; Xia, S.; Shi, W.-Q.; Xue, J.-B.; Li, Y.-Y.; Wu, J.-T. New strains of Japanese encephalitis virus circulating in Shanghai, China after a ten-year hiatus in local mosquito surveillance. Parasites Vectors 2019, 12, 22. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| ICS | NT | Total | |
|---|---|---|---|
| Positive | Negative | ||
| Positive | 172 | 7 | 179 |
| Negative | 13 | 394 | 407 |
| Total | 185 | 401 | 586 |
| Variable | Category | No. Examined | No. Positive | Positive Rate | |
|---|---|---|---|---|---|
| Season | Spring | March | 32 | 8 (25.0%) | 16.4% a |
| April | 41 | 6 (14.6%) | |||
| May | 52 | 5 (9.6%) | |||
| Summer | June | 56 | 10 (17.9%) | 21.5% a | |
| July | 63 | 15 (23.8%) | |||
| August | 57 | 13 (22.8%) | |||
| Autumn | September | 53 | 22 (41.5%) | 46.3% b | |
| October | 71 | 36 (50.7%) | |||
| November | 60 | 28 (46.7%) | |||
| Winter | December | 30 | 12 (40.0%) | 35.7% b | |
| January | 33 | 10 (30.3%) | |||
| February | 38 | 14 (36.8%) | |||
| LivingEnvironment | Domestic dogs | 355 | 73 | 20.6% a | |
| Breeding dogs | 134 | 58 | 43.3% b | ||
| Stray dogs | 97 | 48 | 49.5% b | ||
| Total | 586 | 179 | 29.98% | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, D.; Wahaab, A.; Zheng, J.; Zhang, J.; Ma, Z.; Wei, J. Development of Colloidal Gold-Based Immunochromatographic Strips for Rapid Detection and Surveillance of Japanese Encephalitis Virus in Dogs across Shanghai, China. Viruses 2024, 16, 258. https://doi.org/10.3390/v16020258
Zhong D, Wahaab A, Zheng J, Zhang J, Ma Z, Wei J. Development of Colloidal Gold-Based Immunochromatographic Strips for Rapid Detection and Surveillance of Japanese Encephalitis Virus in Dogs across Shanghai, China. Viruses. 2024; 16(2):258. https://doi.org/10.3390/v16020258
Chicago/Turabian StyleZhong, Dengke, Abdul Wahaab, Jiayang Zheng, Junjie Zhang, Zhiyong Ma, and Jianchao Wei. 2024. "Development of Colloidal Gold-Based Immunochromatographic Strips for Rapid Detection and Surveillance of Japanese Encephalitis Virus in Dogs across Shanghai, China" Viruses 16, no. 2: 258. https://doi.org/10.3390/v16020258
APA StyleZhong, D., Wahaab, A., Zheng, J., Zhang, J., Ma, Z., & Wei, J. (2024). Development of Colloidal Gold-Based Immunochromatographic Strips for Rapid Detection and Surveillance of Japanese Encephalitis Virus in Dogs across Shanghai, China. Viruses, 16(2), 258. https://doi.org/10.3390/v16020258