
Presence of Alphacoronavirus in Tree- and Crevice-Dwelling Bats from Portugal
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
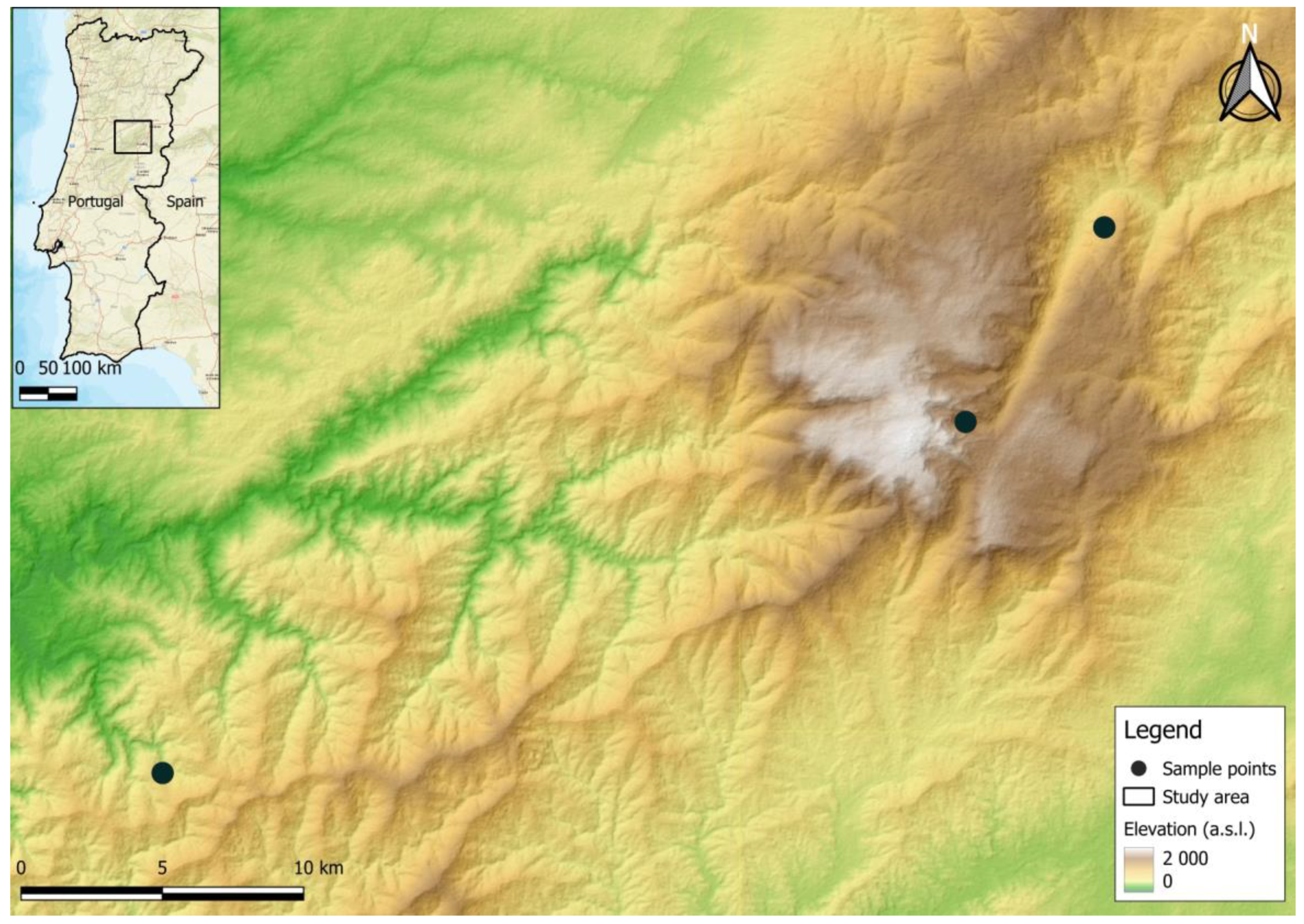
2.1. Sampling Location
2.2. Screening for Coronaviruses
2.3. Sanger Sequencing and Phylogenetic Analysis
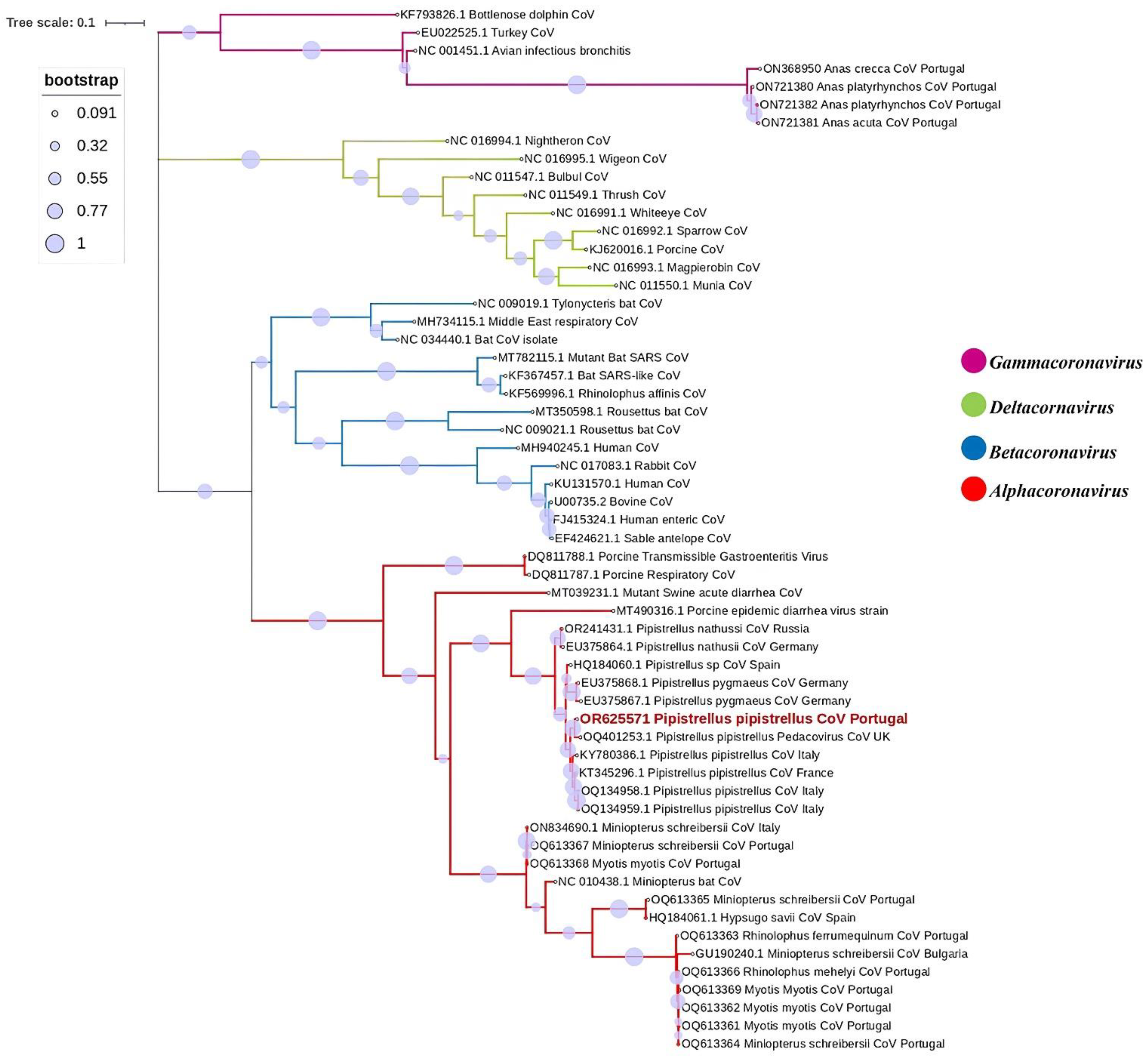
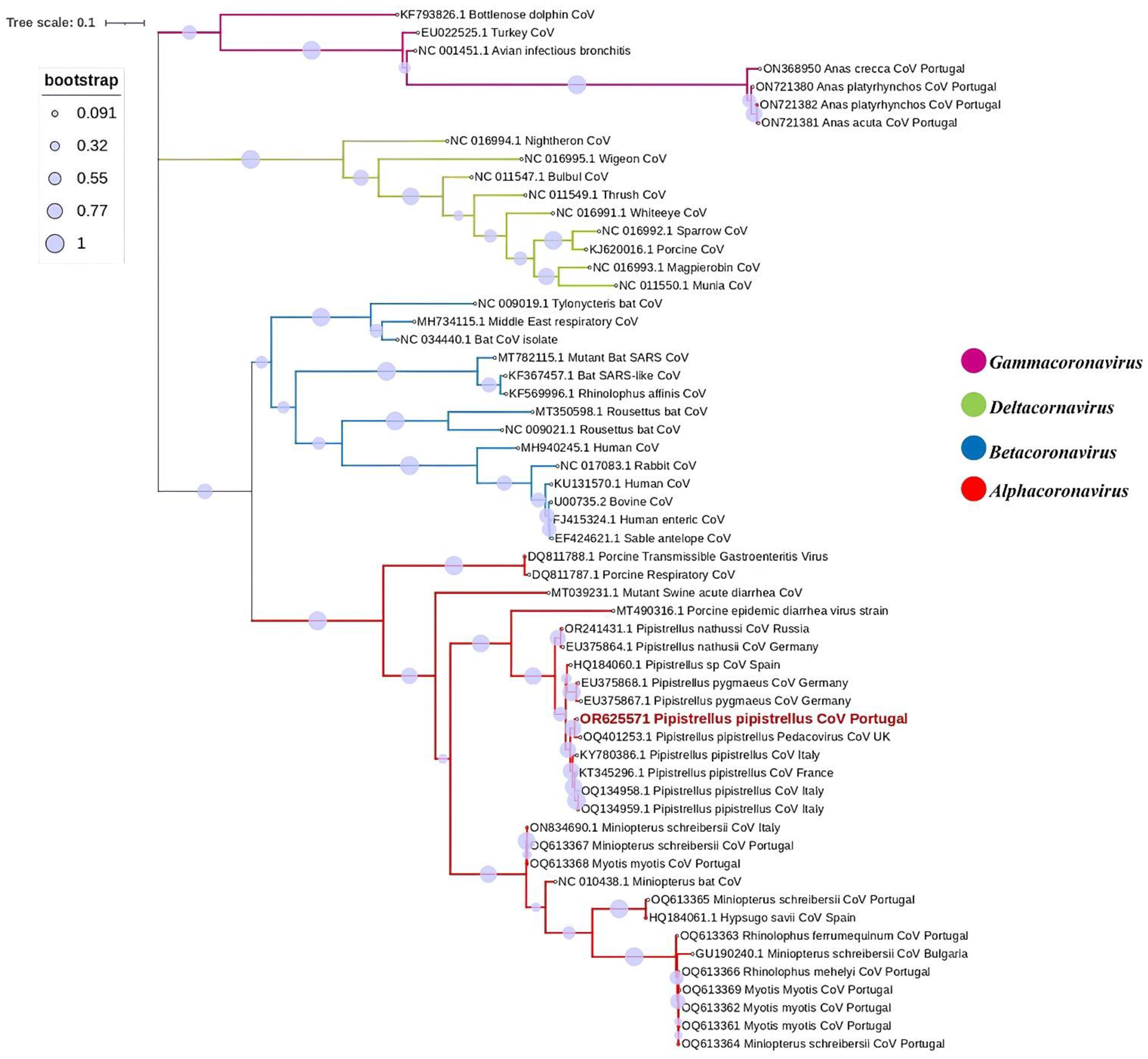
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the Largest RNA Virus Genome. Virus Res. 2006, 117, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.K.; Mifsud, J.C.O.; Costa, V.A.; Grimwood, R.M.; Kitson, J.; Baker, C.; Brosnahan, C.L.; Pande, A.; Holmes, E.C.; Gemmell, N.J.; et al. Slippery When Wet: Cross-Species Transmission of Divergent Coronaviruses in Bony and Jawless Fish and the Evolutionary History of the Coronaviridae. Virus Evol. 2021, 7, veab050. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; De Groot, R.J.; Haagmans, B.; Lau, S.K.P.; Benjamin, W. ICTV Virus Taxonomy Profile: Coronaviridae. J. Gen. Virol. 2023, 104, 001843. [Google Scholar] [CrossRef]
- Bukhari, K.; Mulley, G.; Gulyaeva, A.A.; Zhao, L.; Shu, G.; Jiang, J.; Neuman, B.W. Description and Initial Characterization of Metatranscriptomic Nidovirus-like Genomes from the Proposed New Family Abyssoviridae, and from a Sister Group to the Coronavirinae, the Proposed Genus Alphaletovirus. Virology 2018, 524, 160–171. [Google Scholar] [CrossRef]
- Mordecai, G.J.; Miller, K.M.; Di Cicco, E.; Schulze, A.D.; Kaukinen, K.H.; Ming, T.J.; Li, S.; Tabata, A.; Teffer, A.; Patterson, D.A.; et al. Endangered Wild Salmon Infected by Newly Discovered Viruses. eLife 2019, 8, e47615. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lai, K.K.Y.; Huang, Y.; Lee, P.; Luk, G.S.M.; Dyrting, K.C.; Chan, K.-H.; Yuen, K.-Y. Comparative Analysis of Complete Genome Sequences of Three Avian Coronaviruses Reveals a Novel Group 3c Coronavirus. J. Virol. 2009, 83, 908–917. [Google Scholar] [CrossRef]
- Jacob Machado, D.; Scott, R.; Guirales, S.; Janies, D.A. Fundamental Evolution of All Orthocoronavirinae Including Three Deadly Lineages Descendent from Chiroptera-Hosted Coronaviruses: SARS-CoV, MERS-CoV and SARS-CoV-2. Cladistics 2021, 37, 461–488. [Google Scholar] [CrossRef]
- Letko, M.; Seifert, S.N.; Olival, K.J.; Plowright, R.K.; Munster, V.J. Bat-Borne Virus Diversity, Spillover and Emergence. Nat. Rev. Microbiol. 2020, 18, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Joseph, A.A.; Fagbami, A.H. Coronaviruses: A Review of Their Properties and Diversity. Afr. J. Clin. Exp. Microbiol. 2020, 21, 258–271. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lau, C.C.Y.; Tsang, A.K.L.; Lau, J.H.N.; Bai, R.; Teng, J.L.L.; Tsang, C.C.C.; Wang, M.; et al. Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavi. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, F.; Shi, Z.L. Origin and Evolution of Pathogenic Coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef]
- Han, Y.; Du, J.; Su, H.; Zhang, J.; Zhu, G.; Zhang, S.; Wu, Z.; Jin, Q. Identification of Diverse Bat Alphacoronaviruses and Betacoronaviruses in China Provides New Insights into the Evolution and Origin of Coronavirus-Related Diseases. Front. Microbiol. 2019, 10, 1900. [Google Scholar] [CrossRef]
- Hayman, D.T.S. Bats as Viral Reservoirs. Annu. Rev. Virol. 2016, 3, 77–99. [Google Scholar] [CrossRef]
- Senthilkumar, K.; Marimuthu, G. Tree Roosting Fruit Bats (Chiroptera: Pteropodidae) in Southern Tamil Nadu. Int. J. Appl. BioRes. 2012, 14, 4–10. [Google Scholar]
- Banerjee, A.; Kulcsar, K.; Misra, V.; Frieman, M.; Mossman, K. Bats and Coronaviruses. Viruses 2019, 11, 41. [Google Scholar] [CrossRef]
- Kunz, T.H. Roosting Ecology of Bats. In Ecology of Bats; Springer: Boston, MA, USA, 1982; pp. 1–55. [Google Scholar] [CrossRef]
- Chaverri, G.; Ancillotto, L.; Russo, D. Social Communication in Bats. Biol. Rev. 2018, 93, 1938–1954. [Google Scholar] [CrossRef]
- Ruiz-Aravena, M.; McKee, C.; Gamble, A.; Lunn, T.; Morris, A.; Snedden, C.E.; Yinda, C.K.; Port, J.R.; Buchholz, D.W.; Yeo, Y.Y.; et al. Ecology, Evolution and Spillover of Coronaviruses from Bats; Springer: New York, NY, USA, 2022; Volume 20, ISBN 4157902100652. [Google Scholar]
- Jones, B.D.; Kaufman, E.J.; Peel, A.J. Viral Co-Infection in Bats: A Systematic Review. Viruses 2023, 15, 1860. [Google Scholar] [CrossRef]
- Subudhi, S.; Rapin, N.; Misra, V. Immune System Modulation and Viral Persistence in Bats: Understanding Viral Spillover. Viruses 2019, 11, 192. [Google Scholar] [CrossRef]
- Bokelmann, M.; Balkema-buschmann, A. Coronaviruses in Bats. Berl. Munch. Tierarztl. Wochenschr. 2021, 134, 1–16. [Google Scholar] [CrossRef]
- Mandl, J.N.; Schneider, C.; Schneider, D.S.; Baker, M.L. Going to Bat(s) for Studies of Disease Tolerance. Front. Immunol. 2018, 9, 2112. [Google Scholar] [CrossRef] [PubMed]
- Gorbunova, V.; Seluanov, A.; Kennedy, B.K. The World Goes Bats: Living Longer and Tolerating Viruses. Cell Metab. 2020, 32, 31–43. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, T.J.; Cryan, P.M.; Cunningham, A.A.; Fooks, A.R.; Hayman, D.T.S.; Luis, A.D.; Peel, A.J.; Plowright, R.K.; Wood, J.L.N. Bat Flight and Zoonotic Viruses. Emerg. Infect. Dis. 2014, 20, 741–745. [Google Scholar] [CrossRef]
- Platto, S.; Zhou, J.; Wang, Y.; Wang, H.; Carafoli, E. Biodiversity Loss and COVID-19 Pandemic: The Role of Bats in the Origin and the Spreading of the Disease. Biochem. Biophys. Res. Commun. 2021, 538, 2–13. [Google Scholar] [CrossRef]
- Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Göttsche, M.; Panning, M.; Drexler, J.F.; Petersen, N.; Annan, A.; Grywna, K.; Müller, M.; et al. Detection and Prevalence Patterns of Group I Coronaviruses in Bats, Northern Germany. Emerg. Infect. Dis. 2008, 14, 626–631. [Google Scholar] [CrossRef]
- Annan, A.; Baldwin, H.J.; Corman, V.M.; Klose, S.M.; Owusu, M.; Nkrumah, E.E.; Badu, E.K.; Anti, P.; Agbenyega, O.; Meyer, B.; et al. Human Betacoronavirus 2c EMC/2012-Related Viruses in Bats, Ghana and Europe. Emerg. Infect. Dis. 2013, 19, 456–459. [Google Scholar] [CrossRef]
- Rihtarič, D.; Hostnik, P.; Steyer, A.; Grom, J.; Toplak, I. Identification of SARS-like Coronaviruses in Horseshoe Bats (Rhinolophus hipposideros) in Slovenia. Arch. Virol. 2010, 155, 507–514. [Google Scholar] [CrossRef]
- Lazov, C.M.; Chriél, M.; Baagøe, H.J.; Fjederholt, E.; Deng, Y.; Kooi, E.A.; Belsham, G.J.; Bøtner, A.; Rasmussen, T.B. Detection and Characterization of Distinct Alphacoronaviruses in Five Different Bat Species in Denmark. Viruses 2018, 10, 486. [Google Scholar] [CrossRef]
- Lecis, R.; Mucedda, M.; Pidinchedda, E.; Pittau, M.; Alberti, A. Molecular Identification of Betacoronavirus in Bats from Sardinia (Italy): First Detection and Phylogeny. Virus Genes 2019, 55, 60–67. [Google Scholar] [CrossRef]
- De Sabato, L.; Lelli, D.; Faccin, F.; Canziani, S.; Di Bartolo, I.; Vaccari, G.; Moreno, A. Full Genome Characterization of Two Novel Alpha-Coronavirus Species from Italian Bats. Virus Res. 2019, 260, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Kivistö, I.; Tidenberg, E.M.; Lilley, T.; Suominen, K.; Forbes, K.M.; Vapalahti, O.; Huovilainen, A.; Sironen, T. First Report of Coronaviruses in Northern European Bats. Vector-Borne Zoonotic Dis. 2020, 20, 155–158. [Google Scholar] [CrossRef]
- Lwande, O.W.; Thalin, T.; de Jong, J.; Sjödin, A.; Näslund, J.; Evander, M.; Ecke, F. Alphacoronavirus in a Daubenton’s Myotis Bat (Myotis daubentonii) in Sweden. Viruses 2022, 14, 556. [Google Scholar] [CrossRef]
- Reusken, C.B.E.M.; Lina, P.H.C.; Pielaat, A.; de Vries, A.; Dam-Deisz, C.; Adema, J.; Drexler, J.F.; Drosten, C.; Kooi, E.A. Circulation of Group 2 Coronaviruses in a Bat Species Common to Urban Areas in Western Europe. Vector-Borne Zoonotic Dis. 2010, 10, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Gloza-Rausch, F.; Glende, J.; Corman, V.M.; Muth, D.; Goettsche, M.; Seebens, A.; Niedrig, M.; Pfefferle, S.; Yordanov, S.; et al. Genomic Characterization of Severe Acute Respiratory Syndrome-Related Coronavirus in European Bats and Classification of Coronaviruses Based on Partial RNA-Dependent RNA Polymerase Gene Sequences. J. Virol. 2010, 84, 11336–11349. [Google Scholar] [CrossRef]
- Balboni, A.; Palladini, A.; Bogliani, G.; Battilani, M. Detection of a Virus Related to Betacoronaviruses in Italian Greater Horseshoe Bats. Epidemiol. Infect. 2011, 139, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Corman, V.M.; Wegner, T.; Tateno, A.F.; Zerbinati, R.M.; Gloza-Rausch, F.; Seebens, A.; Müller, M.A.; Drosten, C. Amplification of Emerging Viruses in a Bat Colony. Emerg. Infect. Dis. 2011, 17, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Dallos, B.; Görföl, T.; Boldogh, S.; Estók, P.; Kurucz, K.; Kutas, A.; Földes, F.; Oldal, M.; Németh, V.; et al. Molecular Survey of RNA Viruses in Hungarian Bats: Discovering Novel Astroviruses, Coronaviruses, and Caliciviruses. Vector-Borne Zoonotic Dis. 2014, 14, 846–855. [Google Scholar] [CrossRef]
- Falcón, A.; Vázquez-Morón, S.; Casas, I.; Aznar, C.; Ruiz, G.; Pozo, F.; Perez-Breña, P.; Juste, J.; Ibáñez, C.; Garin, I.; et al. Detection of Alpha and Betacoronaviruses in Multiple Iberian Bat Species. Arch. Virol. 2011, 156, 1883–1890. [Google Scholar] [CrossRef]
- August, T.A.; Mathews, F.; Nunn, M.A. Alphacoronavirus Detected in Bats in the United Kingdom. Vector-Borne Zoonotic Dis. 2012, 12, 530–533. [Google Scholar] [CrossRef]
- Lelli, D.; Papetti, A.; Sabelli, C.; Rosti, E.; Moreno, A.; Boniotti, M.B. Detection of Coronaviruses in Bats of Various Species in Italy. Viruses 2013, 5, 2679–2689. [Google Scholar] [CrossRef]
- De Benedictis, P.; Marciano, S.; Scaravelli, D.; Priori, P.; Zecchin, B.; Capua, I.; Monne, I.; Cattoli, G. Alpha and Lineage C BetaCoV Infections in Italian Bats. Virus Genes 2014, 48, 366–371. [Google Scholar] [CrossRef]
- Goffard, A.; Demanche, C.; Arthur, L.; Pinçon, C.; Michaux, J.; Dubuisson, J. Alphacoronaviruses Detected in French Bats Are Phylogeographically Linked to Coronaviruses of European Bats. Viruses 2015, 7, 6279–6290. [Google Scholar] [CrossRef]
- Fischer, K.; Zeus, V.; Kwasnitschka, L.; Kerth, G.; Haase, M.; Groschup, M.H.; Balkema-Buschmann, A. Insectivorous Bats Carry Host Specific Astroviruses and Coronaviruses across Different Regions in Germany. Infect. Genet. Evol. 2016, 37, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Monchatre-Leroy, E.; Boué, F.; Boucher, J.M.; Renault, C.; Moutou, F.; Gouilh, M.A.; Umhang, G. Identification of Alpha and Beta Coronavirus in Wildlife Species in France: Bats, Rodents, Rabbits, and Hedgehogs. Viruses 2017, 9, 364. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, O.; Dubina, D.; Sokolovskyi, D.; Gaidash, O. Possible Zoonotic Viral Threats Associated with Bats in Southern Ukraine. Proc. Theriol. Sch. 2017, 2017, 150–153. [Google Scholar] [CrossRef]
- Pauly, M.; Pir, J.B.; Loesch, C.; Sausy, A.; Snoeck, C.J.; Hübschen, J.M.; Muller, C.P. Novel Alphacoronaviruses and Paramyxoviruses Cocirculate with Type 1 and Severe Acute Respiratory System (SARS)-Related Betacoronaviruses in Synanthropic Bats of Luxembourg. Appl. Environ. Microbiol. 2017, 83, e01326-17. [Google Scholar] [CrossRef]
- Moreno, A.; Lelli, D.; De Sabato, L.; Zaccaria, G.; Boni, A.; Sozzi, E.; Prosperi, A.; Lavazza, A.; Cella, E.; Castrucci, M.R.; et al. Detection and Full Genome Characterization of Two Beta CoV Viruses Related to Middle East Respiratory Syndrome from Bats in Italy. Virol. J. 2017, 14, 239. [Google Scholar] [CrossRef] [PubMed]
- Hemnani, M.; da Silva, P.G.; Thompson, G.; Poeta, P.; Rebelo, H.; Mesquita, J.R. First Report of Alphacoronavirus Circulating in Cavernicolous Bats from Portugal. Viruses 2023, 15, 1521. [Google Scholar] [CrossRef]
- da Luz Mathias, M.; Fonseca, C.; Rodrigues, L.; Grilo, C.; Lopes-Fernandes, M.; Palmeirim, J.M.; Santos-Reis, M.; Alves, P.C.; Cabral, J.A.; Ferreira, M.; et al. Livro Vermelho Dos Mamíferos de Portugal Continental, 2023rd ed.; FCiências.ID, ICNF: Lisboa, Portugal, 2023. [Google Scholar]
- Drzewnioková, P.; Festa, F.; Panzarin, V.; Lelli, D.; Moreno, A.; Zecchin, B.; De Benedictis, P.; Leopardi, S. Best Molecular Tools to Investigate Coronavirus Diversity in Mammals: A Comparison. Viruses 2021, 13, 1975. [Google Scholar] [CrossRef]
- Hemnani, M.; Rodrigues, D.; Santos, N.; Santos-Silva, S.; Figueiredo, M.E.; Henriques, P.; Ferreira-e-Silva, J.; Rebelo, H.; Poeta, P.; Thompson, G.; et al. Molecular Detection and Characterization of Coronaviruses in Migratory Ducks from Portugal Show the Circulation of Gammacoronavirus and Deltacoronavirus. Animals 2022, 12, 3283. [Google Scholar] [CrossRef]
- Wilkinson, D.A.; Joffrin, L.; Lebarbenchon, C.; Mavingui, P. Analysis of Partial Sequences of the RNA-Dependent RNA Polymerase Gene as a Tool for Genus and Subgenus Classification of Coronaviruses. J. Gen. Virol. 2021, 101, 1261–1269. [Google Scholar] [CrossRef]
- Zhang, D.; Kan, X.; Huss, S.E.; Jiang, L.; Chen, L.Q.; Hu, Y. Using Phylogenetic Analysis to Investigate Eukaryotic Gene Origin. J. Vis. Exp. 2018, 2018, e56684. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Bittar, C.; Machado, R.R.G.; Comelis, M.T.; Bueno, L.M.; Beguelini, M.R.; Morielle-Versute, E.; Nogueira, M.L.; Rahal, P. Alphacoronavirus Detection in Lungs, Liver, and Intestines of Bats from Brazil. Microb. Ecol. 2020, 79, 203–212. [Google Scholar] [CrossRef]
- Bueno, L.M.; Rizotto, L.S.; de Oliveira Viana, A.; Silva, L.M.N.; de Moraes, M.V.d.S.; Benassi, J.C.; Scagion, G.P.; Dorlass, E.G.; Lopes, B.L.T.; Cunha, I.N.; et al. High Genetic Diversity of Alphacoronaviruses in Bat Species (Mammalia: Chiroptera) from the Atlantic Forest in Brazil. Transbound. Emerg. Dis. 2022, 69, e2863–e2875. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, F.; Edenborough, K.M.; Toffoli, R.; Culasso, P.; Zoppi, S.; Dondo, A.; Robetto, S.; Rosati, S.; Lander, A.; Kurth, A.; et al. Coronavirus and Paramyxovirus in Bats from Northwest Italy. BMC Vet. Res. 2017, 13, 396. [Google Scholar] [CrossRef] [PubMed]
- Hulva, P.; Horáček, I.; Strelkov, P.P.; Benda, P. Molecular Architecture of Pipistrellus pipistrellus/Pipistrellus pygmaeus Complex (Chiroptera: Vespertilionidae): Further Cryptic Species and Mediterranean Origin of the Divergence. Mol. Phylogenet. Evol. 2004, 32, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Frick, W.F.; Kingston, T.; Flanders, J. A Review of the Major Threats and Challenges to Global Bat Conservation. Ann. N. Y. Acad. Sci. 2020, 1469, 5–25. [Google Scholar] [CrossRef]
- Dietz, M.; Brombacher, M.; Erasmy, M.; Fenchuk, V.; Simon, O. Bat Community and Roost Site Selection of Tree-Dwelling Bats in a Well-Preserved European Lowland Forest. Acta Chiropterologica 2018, 20, 117–127. [Google Scholar] [CrossRef]
- Bryja, J.; Kaňuch, P.; Fornůsková, A.; Bartonička, T.; Řehák, Z. Low Population Genetic Structuring of Two Cryptic Bat Species Suggests Their Migratory Behaviour in Continental Europe. Biol. J. Linn. Soc. 2009, 96, 103–114. [Google Scholar] [CrossRef]
- Nusová, G.; Uhrin, M.; Voigt, C.C.; Kaňuch, P. Tracing the Geographic Origin of Common Pipistrelles (Pipistrellus pipistrellus) Swarming at a Mass Hibernaculum. Mamm. Biol. 2020, 100, 601–610. [Google Scholar] [CrossRef]
- Brook, C.E.; Dobson, A.P. Bats as “special” Reservoirs for Emerging Zoonotic Pathogens. Trends Microbiol. 2015, 23, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Kasso, M.; Balakrishnan, M. Ecological and Economic Importance of Bats (Order Chiroptera). ISRN Biodivers. 2013, 2013, 187415. [Google Scholar] [CrossRef]
- Moussy, C.; Hosken, D.J.; Mathews, F.; Smith, G.C.; Aegerter, J.N.; Bearhop, S. Migration and Dispersal Patterns of Bats and Their Influence on Genetic Structure. Mamm. Rev. 2013, 43, 183–195. [Google Scholar] [CrossRef]
- Wong, S.; Lau, S.; Woo, P.; Yuen, K.Y. Bats as a Continuing Source of Emerging Infections in Humans. Rev. Med. Virol. 2007, 17, 67–91. [Google Scholar] [CrossRef]
- Watanabe, S.; Masangkay, J.S.; Nagata, N.; Morikawa, S.; Mizutani, T.; Fukushi, S.; Alviola, P.; Omatsu, T.; Ueda, N.; Iha, K.; et al. Bat Coronaviruses and Experimental Infection of Bats, the Philippines. Emerg. Infect. Dis. 2010, 16, 1217–1223. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Location | Species | Number of Individuals Found in Each Location | Stool Samples Collected from Each Species |
|---|---|---|---|
| Serra da Estrela, day one | Pipistrellus pipistrellus | 11 | 10 |
| Eptesicus serotinus | 2 | 2 | |
| Myotis emarginatus | 2 | 2 | |
| Nyctalus leisleri | 5 | 4 | |
| Barbastella barbastellus | 1 | 0 | |
| Nyctalus lasiopterus | 1 | 0 | |
| Serra da Estrela, day two | Eptesicus serotinus/isabellinus | 2 | 2 |
| Myotis daubentonii | 1 | 1 | |
| Plecotus austriacus | 1 | 1 | |
| Plecotus auritus | 6 | 6 | |
| Nyctalus leisleri | 1 | 1 | |
| Hypsugo savii | 4 | 3 | |
| Pipistrellus pipistrellus | 11 | 11 | |
| Myotis mystacinus | 3 | 3 | |
| Eptesicus serotinus | 1 | 0 | |
| Barbastella barbastellus | 1 | 1 | |
| Serra do Açor | Nyctalus leisleri | 13 | 12 |
| Nyctalus lasiopterus | 10 | 10 | |
| Pipistrellus pipistrellus | 2 | 2 | |
| Plecotus auritus | 15 | 13 | |
| Eptesicus serotinus | 1 | 1 | |
| Myotis bechsteinii | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hemnani, M.; da Silva, P.G.; Thompson, G.; Poeta, P.; Rebelo, H.; Mesquita, J.R. Presence of Alphacoronavirus in Tree- and Crevice-Dwelling Bats from Portugal. Viruses 2024, 16, 434. https://doi.org/10.3390/v16030434
Hemnani M, da Silva PG, Thompson G, Poeta P, Rebelo H, Mesquita JR. Presence of Alphacoronavirus in Tree- and Crevice-Dwelling Bats from Portugal. Viruses. 2024; 16(3):434. https://doi.org/10.3390/v16030434
Chicago/Turabian StyleHemnani, Mahima, Priscilla Gomes da Silva, Gertrude Thompson, Patrícia Poeta, Hugo Rebelo, and João R. Mesquita. 2024. "Presence of Alphacoronavirus in Tree- and Crevice-Dwelling Bats from Portugal" Viruses 16, no. 3: 434. https://doi.org/10.3390/v16030434
APA StyleHemnani, M., da Silva, P. G., Thompson, G., Poeta, P., Rebelo, H., & Mesquita, J. R. (2024). Presence of Alphacoronavirus in Tree- and Crevice-Dwelling Bats from Portugal. Viruses, 16(3), 434. https://doi.org/10.3390/v16030434