
Brincidofovir Effectively Inhibits Proliferation of Pseudorabies Virus by Disrupting Viral Replication



Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Construction of Fluorescently Labeled Viruses
2.3. Cytotoxicity Test
2.4. Half Maximal Inhibitory Concentration (IC50) Determination
2.5. The 50% Tissue Culture Infectious Dose (TCID50) Assay
2.6. Quantitative PCR (qPCR) Detecting gE Gene
2.7. One-Step Growth Curve
2.8. In Vivo Assessment of Antiviral Effects of the Drugs
2.9. Time-of-Addition Assay
2.10. Statistical Analysis
3. Results
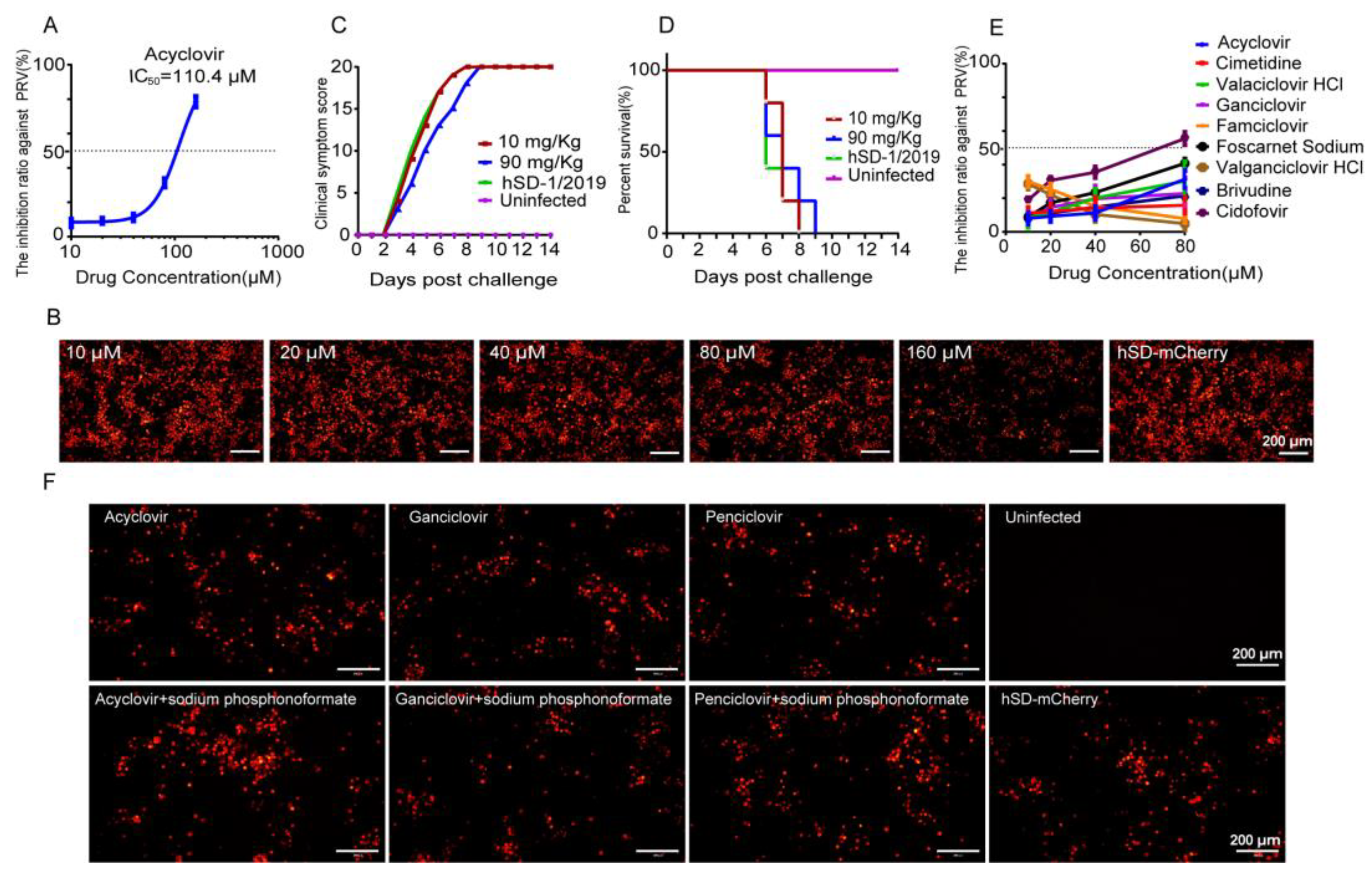
3.1. Fluorescence Intensity of the Recombinant PRV-mCherry Could Indicate Virus Proliferation Titers
3.2. The Anti-Herpesvirus Drugs Commonly Used in Clinical Practice Exhibited Limited Efficacy against the PRV Variant Strain hSD-1/2019
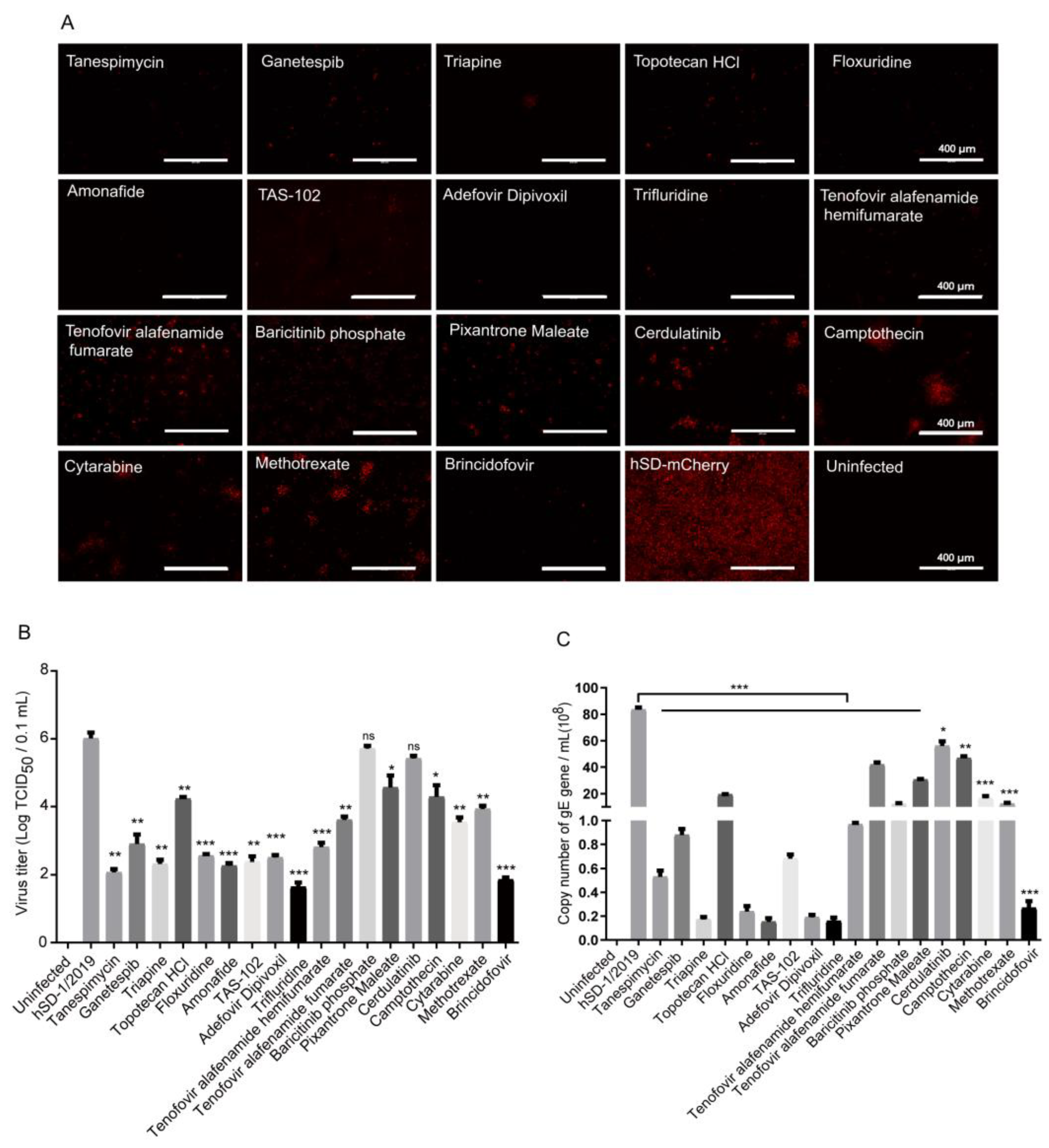
3.3. Eighteen Drugs Effectively Inhibiting PRV-mCherry Proliferation Were Screened out from 2104 FDA-Approved Drugs through the High-Throughput Screening Method
3.4. In Vitro Evaluation of the Anti-PRV Drug Candidates
3.5. In Vivo Evaluation of the 10 Drug Candidates
3.6. Brincidofovir Inhibits Virus Proliferation Mainly by Interfering with the Viral Replication Phase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. MMBR 2005, 69, 462–500. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Xie, C.; Ding, S.; Yang, H.; Guo, S.; Li, J.; Qin, L.; Ban, F.; Wang, D.; et al. A novel human acute encephalitis caused by pseudorabies virus variant strain. Clin. Infect. Dis. 2020, 73, 3690–3700. [Google Scholar] [CrossRef]
- Wu, T.; Fei, L.; Hao, Z.; Liang, C.; Tong, G.Z. Emergence of a Pseudorabies virus variant with increased virulence to piglets. Vet. Microbiol. 2015, 181, 236–240. [Google Scholar]
- Wang, C.H.; Yuan, J.; Qin, H.Y.; Luo, Y.; Cong, X.; Li, Y.; Chen, J.; Li, S.; Sun, Y.; Qiu, H.J. A novel gE-deleted pseudorabies virus (PRV) provides rapid and complete protection from lethal challenge with the PRV variant emerging in Bartha-K61-vaccinated swine population in China. Vaccine 2014, 32, 3379–3385. [Google Scholar] [CrossRef]
- Skinner, G.; Ahmad, A.; Davies, J.A. The infrequency of transmission of herpesviruses between humans and animals; postulation of an unrecognised protective host mechanism. Comp. Immunol. Microbiol. Infect. Dis. 2001, 24, 255–269. [Google Scholar] [CrossRef]
- Tan, L.; Shu, X.; Xu, K.; Liao, F.; Song, C.; Duan, D.; Yang, S.; Yao, J.; Wang, A. Homologous recombination technology generated recombinant pseudorabies virus expressing EGFP facilitates to evaluate its susceptibility to different cells and screen antiviral compounds. Res. Vet. Sci. 2022, 145, 125–134. [Google Scholar] [CrossRef]
- Fan, S.; Yuan, H.; Liu, L.; Li, H.; Wang, S.; Zhao, W.; Wu, Y.; Wang, P.; Hu, Y.; Han, J.; et al. Pseudorabies virus encephalitis in humans: A case series study. J. Neurovirol. 2020, 26, 556–564. [Google Scholar] [CrossRef]
- Zhao, W.; Wu, Y.; Li, H.; Li, S.; Fan, S.; Wu, H.; Li, Y.; Lü, Y.; Han, J.; Zhang, W.; et al. Clinical experience and next-generation sequencing analysis of encephalitis caused by pseudorabies virus. Zhonghua Yi Xue Za Zhi 2018, 98, 1152–1157. [Google Scholar] [CrossRef]
- Wang, D.; Tao, X.; Fei, M.; Chen, J.; Guo, W.; Li, P.; Wang, J. Human encephalitis caused by pseudorabies virus infection: A case report. J. Neurovirol. 2020, 26, 442–448. [Google Scholar] [CrossRef]
- Wang, Y.; Nian, H.; Li, Z.; Wang, W.; Wang, X.; Cui, Y. Human encephalitis complicated with bilateral acute retinal necrosis associated with pseudorabies virus infection: A case report. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2019, 89, 51–54. [Google Scholar] [CrossRef]
- Yang, X.; Guan, H.; Li, C.; Li, Y.; Wang, S.; Zhao, X.; Zhao, Y.; Liu, Y. Characteristics of human encephalitis caused by pseudorabies virus: A case series study. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2019, 87, 92–99. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, X.; Yuan, D.; Li, R.; Lu, J.; Li, X.; Tian, K.; Dai, E. Dynamic cerebrospinal fluid analyses of severe pseudorabies encephalitis. Transbound. Emerg. Dis. 2019, 66, 2562–2565. [Google Scholar] [CrossRef]
- Yang, H.; Han, H.; Wang, H.; Cui, Y.; Liu, H.; Ding, S. A Case of Human Viral Encephalitis Caused by Pseudorabies Virus Infection in China. Front. Neurol. 2019, 10, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Wang, J.; Peng, X. Bilateral Necrotizing Retinitis following Encephalitis Caused by the Pseudorabies Virus Confirmed by Next-Generation Sequencing. Ocul. Immunol. Inflamm. 2020, 29, 922–925. [Google Scholar] [CrossRef]
- Ai, J.W.; Weng, S.S.; Cheng, Q.; Cui, P.; Li, Y.J.; Wu, H.L.; Zhu, Y.M.; Xu, B.; Zhang, W.H. Human Endophthalmitis Caused By Pseudorabies Virus Infection, China, 2017. Emerg. Infect. Dis. 2018, 24, 1087–1090. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, T.X.; Wang, T.Y.; Tang, Y.D.; Wei, P. Isobavachalcone inhibits Pseudorabies virus by impairing virus-induced cell-to-cell fusion. Virol. J. 2020, 17, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Huan, C.; Xu, W.; Guo, T.; Pan, H.; Gao, S. (-)-Epigallocatechin-3-Gallate Inhibits the Life Cycle of Pseudorabies Virus In Vitro and Protects Mice Against Fatal Infection. Front. Cell. Infect. Microbiol. 2021, 10, 895–908. [Google Scholar] [CrossRef]
- Zhao, X.; Tong, W.; Song, X.; Jia, R.; Li, L.; Zou, Y.; He, C.; Liang, X.; Cheng, L.; Bo, J. Antiviral Effect of Resveratrol in Piglets Infected with Virulent Pseudorabies Virus. Viruses 2018, 10, 457. [Google Scholar] [CrossRef]
- Li, L.; Wang, R.; Hu, H.; Chen, X.; Yin, Z.; Liang, X.; He, C.; Yin, L.; Ye, G.; Zou, Y.; et al. The antiviral activity of kaempferol against pseudorabies virus in mice. BMC Vet. Res. 2021, 17, 247–258. [Google Scholar] [CrossRef]
- Sun, Y.; Li, C.; Li, Z.; Shangguan, A.; Jiang, J.; Zeng, W.; Zhang, S.; He, Q. Quercetin as an antiviral agent inhibits the Pseudorabies virus in vitro and in vivo. Virus Res. 2021, 305, 556–564. [Google Scholar] [CrossRef]
- Fang, L.; Gao, Y.; Lan, M.; Jiang, P.; Bai, J.; Li, Y.; Wang, X. Hydroquinone inhibits PRV infection in neurons in vitro and in vivo. Vet. Microbiol. 2020, 250, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chen, R.; Huang, P.; Hong, J.; Cao, J.; Wu, Q.; Zheng, W.; Lin, L.; Han, Q.; Chen, Y.; et al. Adefovir dipivoxil efficiently inhibits the proliferation of pseudorabies virus in vitro and in vivo. Antivir. Res. 2021, 186, 14–25. [Google Scholar] [CrossRef]
- Lv, C.; Liu, W.; Wang, B.; Dang, R.; Qiu, L.; Ren, J.; Yan, C.; Yang, Z.; Wang, X. Ivermectin inhibits DNA polymerase UL42 of pseudorabies virus entrance into the nucleus and proliferation of the virus in vitro and vivo. Antivir. Res. 2018, 159, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Nie, C.; Wen, H.; Long, Y.; Zhou, M.; Xie, Z.; Hong, D. Human viral encephalitis associated with suid herpesvirus 1. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2022, 43, 2681–2692. [Google Scholar] [CrossRef] [PubMed]
- Russ, P.; Schelling, P.; Scapozza, L.; Folkers, G.; Clercq, E.; Marquez, V. Synthesis and biological evaluation of 5-substituted derivatives of the potent antiherpes agent (north)-methanocarbathymine. J. Med. Chem. 2003, 46, 5045–5054. [Google Scholar] [CrossRef] [PubMed]
- Brancale, A.; McGuigan, C.; Algain, B.; Savy, P.; Benhida, R.; Fourrey, J.; Andrei, G.; Snoeck, R.; De Clercq, E.; Balzarini, J. Bicyclic anti-VZV nucleosides: Thieno analogues retain full antiviral activity. Bioorganic Med. Chem. Lett. 2001, 11, 2507–2510. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Turnage, C.T.; Gaston, W.D.; Arruda, P.; Alls, S.A.; Gidlewski, T. Pseudorabies detected in hunting dogs in Alabama and Arkansas after close contact with feral swine (Sus scrofa). BMC Vet. Res. 2018, 14, 388–395. [Google Scholar] [CrossRef]
- Chiari, M.; Ferrari, N.; Bertoletti, M.; Avisani, D.; Cerioli, M.; Zanoni, M.; Alborali, L.G.; Lanfranchi, P.; Lelli, D.; Martin, A.M. Long-Term Surveillance of Aujeszky’s Disease in the Alpine Wild Boar (Sus scrofa). EcoHealth 2015, 12, 563–570. [Google Scholar] [CrossRef]
- Sun, Y.; Luo, Y.; Wang, C.H.; Yuan, J.; Li, N.; Song, K.; Qiu, H.J. Control of swine pseudorabies in China: Opportunities and limitations. Vet. Microbiol. 2016, 183, 119–124. [Google Scholar] [CrossRef]
- Lu, J.J.; Yuan, W.Z.; Zhu, Y.P.; Hou, S.H.; Wang, X.J. Latent pseudorabies virus infection in medulla oblongata from quarantined pigs. Transbound. Emerg. Dis. 2021, 68, 543–551. [Google Scholar] [CrossRef]
- Tan, L.; Yao, J.; Yang, Y.; Luo, W.; Yuan, X.; Yang, L.; Wang, A. Current Status and Challenge of Pseudorabies Virus Infection in China. Virol. Sin. 2021, 36, 588–607. [Google Scholar] [CrossRef] [PubMed]
- Bialas, K.M.; Swamy, G.K.; Permar, S.R. Perinatal cytomegalovirus and varicella zoster virus infections: Epidemiology, prevention, and treatment. Clin. Perinatol. 2015, 42, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.M.; Kudesia, G.; Mckendrick, M. Management of herpesvirus infections. Int. J. Antimicrob. Agents 2013, 42, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cui, Q.; Fu, Q.; Song, X.; Jia, R.; Yang, Y.; Zou, Y.; Li, L.; He, C.; Liang, X. Antiviral properties of resveratrol against pseudorabies virus are associated with the inhibition of IκB kinase activation. Sci. Rep. 2017, 7, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Andreu, S.; Ripa, I.; Praena, B.; López-Guerrero, J.A.; Bello-Morales, R. The Valproic Acid Derivative Valpromide Inhibits Pseudorabies Virus Infection in Swine Epithelial and Mouse Neuroblastoma Cell Lines. Viruses 2021, 13, 2522. [Google Scholar] [CrossRef] [PubMed]
- Florescu, D.F.; Keck, M.A. Development of CMX001 (Brincidofovir) for the treatment of serious diseases or conditions caused by dsDNA viruses. Expert Rev. Anti-Infect. Ther. 2014, 12, 1171–1178. [Google Scholar] [CrossRef]
- Feghoul, L.; Mercier-Delarue, S.; Salmona, M.; Ntsiba, N.; Dalle, J.-H.; Baruchel, A.; Klonjkowski, B.; Richardson, J.; Simon, F.; LeGoff, J. Genetic diversity of the human adenovirus species C DNA polymerase. Antivir. Res. 2018, 159, 1–9. [Google Scholar] [CrossRef]
- Alvarez-Cardona, J.J.; Whited, L.K.; Chemaly, R.F. Brincidofovir: Understanding its unique profile and potential role against adenovirus and other viral infections. Future Microbiol. 2020, 15, 389–400. [Google Scholar] [CrossRef]
- Hutson, C.L.; Kondas, A.V.; Mauldin, M.R.; Doty, J.B.; Olson, V.A. Pharmacokinetics and Efficacy of a Potential Smallpox Therapeutic, Brincidofovir, in a Lethal Monkeypox Virus Animal Model. mSphere 2021, 6, 927–932. [Google Scholar] [CrossRef]
- Randall, L.E.; Scott, F.; Tom, B.; Sunwen, C.; Prichard, M.N.; Steven, K.; Chad, W.; Donella, C.; Herve, M.M. Analysis of Mutations in the Gene Encoding Cytomegalovirus DNA Polymerase in a Phase 2 Clinical Trial of Brincidofovir Prophylaxis. J. Infect. Dis. 2016, 214, 32–35. [Google Scholar]
- Quenelle, D.C.; Lampert, B.; Collins, D.J.; Rice, T.L.; Painter, G.R.; Kern, E.R. Efficacy of CMX001 against Herpes Simplex Virus Infections in Mice and Correlations with Drug Distribution Studies. J. Infect. Dis. 2010, 202, 1492–1499. [Google Scholar] [CrossRef] [PubMed]
- Gosert, R.; Rinaldo, C.H.; Wernli, M.; Major, E.O.; Hirsch, H.H. CMX001 (1-O-Hexadecyloxypropyl-Cidofovir) Inhibits Polyomavirus JC Replication in Human Brain Progenitor-Derived Astrocytes. Antimicrob. Agents Chemother. 2011, 55, 2129–2136. [Google Scholar] [CrossRef] [PubMed]
- Olson, V.A.; Smith, S.K.; Foster, S.; Li, Y.; Lanier, E.R.; Gates, I.; Trost, L.C.; Damon, I.K. In Vitro Efficacy of Brincidofovir against Variola Virus. Antimicrob. Agents Chemother. 2014, 58, 5570–5571. [Google Scholar] [CrossRef]
- Lanier, R.; Trost, L.; Tippin, T.; Lampert, B.; Robertson, A.; Foster, S.; Rose, M.; Painter, W.; O’Mahony, R.; Almond, M.; et al. Development of CMX001 for the Treatment of Poxvirus Infections. Viruses 2010, 2, 2740–2762. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Sequences | Primer | Sequences (5′-3′) |
|---|---|---|
| Upstream homologous arm | PRV-HindIII-F | CCCAAGCTTAGGCCGCGTACCCTCCG |
| PRV-KpnI-R | CGGGGTACCGGGCGAGGGGCGAGGG | |
| mCherry | mCherry-KpnI-F | CGGGGTACCGGTGGAGGCGGTTCAGGCGGAG GTGGCTCTATGGTGAGCAAGGGCGAGGA |
| mCherry-BamHI-R | CGCGGATCCCTTGTACAGCTCGTCCATGC | |
| Downstream homologous arm | PRV-BamHI-F | CGCGGATCCTAGCCCCGCGCGATCAATAAAG |
| PRV-EcoRI-R | CCGGAATTCCCGCGCGTGGTGGAGTCG |
| Score | Clinical Symptoms |
|---|---|
| 0 | No symptoms |
| 1 | Excitement, restlessness, occasional itching and scratching |
| 2 | Ataxia, severe itching, persistent gnawing on hind limbs |
| 3 | Gnawing on the hind limbs resulting in bone disruption and tissue necrosis |
| 4 | Dead or dying |
| Number | Drug | Target |
|---|---|---|
| 1 | Tanespimycin (17-AAG) | Cytoskeletal Signaling |
| 2 | Ganetespib (STA-9090) | Cytoskeletal Signaling |
| 3 | Triapine | DNA/RNA Synthesis |
| 4 | Topotecan HCl | DNA Damage |
| 5 | Floxuridine | DNA Damage |
| 6 | Amonafide | DNA Damage |
| 7 | TAS-102 | DNA/RNA Synthesis |
| 8 | Adefovir Dipivoxil | Microbiology |
| 9 | Trifluridine | DNA Damage |
| 10 | Tenofovir alafenamide hemifumarate | Reverse Transcriptase |
| 11 | Tenofovir alafenamide fumarate | Reverse Transcriptase |
| 12 | Baricitinib phosphate | JAK/STAT |
| 13 | Pixantrone Maleate | DNA Damage |
| 14 | Cerdulatinib (PRT062070) | JAK/STAT |
| 15 | Camptothecin | Topoisomerase |
| 16 | Cytarabine hydrochloride | DNA Damage |
| 17 | Methotrexate | Metabolism |
| 18 | Brincidofovir | DNA/RNA Synthesis |
| Number | Drug | CC50 (μM) | IC50 (μM) | SI (CC50/IC50) |
|---|---|---|---|---|
| 1 | Tanespimycin (17-AAG) | 48.89 | 0.46 | 106.08 |
| 2 | Ganetespib (STA-9090) | 294.51 | 0.07 | 3937.17 |
| 3 | Triapine | 78.66 | 2.99 | 26.29 |
| 4 | Floxuridine | 375.43 | 0.23 | 1653.74 |
| 5 | Amonafide | 241.71 | 1.24 | 194.76 |
| 6 | TAS-102 | 1124.00 | 0.95 | 1186.66 |
| 7 | Adefovir Dipivoxil | 227.61 | 4.08 | 55.84 |
| 8 | Trifluridine | 352.40 | 1.43 | 246.09 |
| 9 | Tenofovir hemifumarate | 225.95 | 2.79 | 81.09 |
| 10 | Brincidofovir | 22.32 | 0.54 | 41.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Liu, Q.; Yang, D.; Zhang, H.; Kuang, Y.; Li, Y.; Chen, H.; Wang, X. Brincidofovir Effectively Inhibits Proliferation of Pseudorabies Virus by Disrupting Viral Replication. Viruses 2024, 16, 464. https://doi.org/10.3390/v16030464
Guo H, Liu Q, Yang D, Zhang H, Kuang Y, Li Y, Chen H, Wang X. Brincidofovir Effectively Inhibits Proliferation of Pseudorabies Virus by Disrupting Viral Replication. Viruses. 2024; 16(3):464. https://doi.org/10.3390/v16030464
Chicago/Turabian StyleGuo, Huihui, Qingyun Liu, Dan Yang, Hao Zhang, Yan Kuang, Yafei Li, Huanchun Chen, and Xiangru Wang. 2024. "Brincidofovir Effectively Inhibits Proliferation of Pseudorabies Virus by Disrupting Viral Replication" Viruses 16, no. 3: 464. https://doi.org/10.3390/v16030464
APA StyleGuo, H., Liu, Q., Yang, D., Zhang, H., Kuang, Y., Li, Y., Chen, H., & Wang, X. (2024). Brincidofovir Effectively Inhibits Proliferation of Pseudorabies Virus by Disrupting Viral Replication. Viruses, 16(3), 464. https://doi.org/10.3390/v16030464