

Detection of Acipenser European Iridovirus (AcIV-E) in Sturgeon Farms in Northern Italy between 2021–2023
 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Anatomopathological Examination
2.3. DNA Extraction and Real-Time-PCR
3. Results
3.1. Virological Analysis

{kind=link}
| Species | No. of Subjects | Positive (%) | Negative (%) |
|---|---|---|---|
| Acipenser gueldenstaedtii | 257 | 164 (63.81) | 93 (36.19) |
| Acipenser baerii | 50 | 6 (12.00) | 44 (88.00) |
| Acipenser ruthenus | 17 | 2 (11.76) | 15 (88.24) |
| Acipenser transmontanus | 22 | 0 | 22 (100) |
| Acipenser naccarii | 47 | 19 (40.43) | 28 (59.57) |
| Acipenser stellatus | 56 | 13 (23.21) | 43 (76.79) |
| Huso huso | 33 | 0 | 33 (100) |
| Species | Year | Total | Positive (%) |
|---|---|---|---|
| Acipenser gueldenstaedtii | 2021 | 136 | 86 (63.24) |
| 2022 | 85 | 52 (61.18) | |
| 2023 | 36 | 26 (72.22) | |
| Acipenser baerii | 2021 | 0 | 0 |
| 2022 | 20 | 4 (20.00) | |
| 2023 | 30 | 2 (6.67) | |
| Acipenser ruthenus | 2021 | 0 | 0 |
| 2022 | 10 | 2 (20.00) | |
| 2023 | 7 | 0 | |
| Acipenser transmontanus | 2021 | 2 | 0 |
| 2022 | 10 | 0 | |
| 2023 | 10 | 0 | |
| Acipenser naccarii | 2021 | 0 | 0 |
| 2022 | 36 | 13 (36.11) | |
| 2023 | 11 | 6 (54.55) | |
| Acipenser stellatus | 2021 | 10 | 2 (20.00) |
| 2022 | 21 | 10 (47.62) | |
| 2023 | 25 | 1 (4.00) | |
| Huso huso | 2021 | 10 | 0 |
| 2022 | 18 | 0 | |
| 2023 | 5 | 0 |
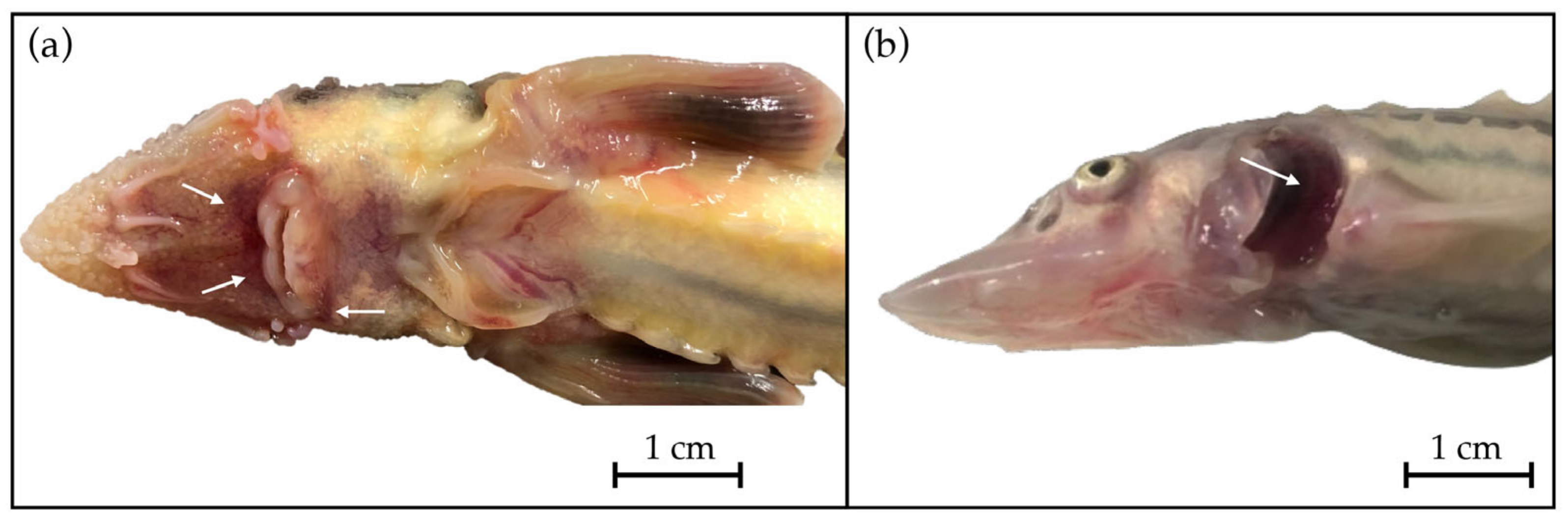
3.2. Anatomopathological Examination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciulli, S.; Volpe, E.; Sirri, R.; Passalacqua, P.L.; Cesa Bianchi, F.; Serratore, P.; Mandrioli, L. Outbreak of mortality in Russian (Acipenser gueldenstaedtii) and Siberian (Acipenser baerii) sturgeons associated with sturgeon nucleo-cytoplasmatic large DNA virus. Vet. Microbiol. 2016, 191, 27–34. [Google Scholar] [CrossRef]
- Bronzi, P.; Chebanov, M.; Michaels, J.T.; Wei, Q.; Rosenthal, H.; Gessner, J. Sturgeon meat and caviar production: Global update 2017. J. Appl. Ichthyol. 2019, 35, 257–266. [Google Scholar] [CrossRef]
- Pikitch, E.K.; Doukakis, P.; Lauck, L.; Chakrabarty, P.; Erickson, D.L. Status, trends and management of sturgeon and paddlefish fisheries. Fish Fish. 2005, 6, 233–265. [Google Scholar] [CrossRef]
- Ruban, G.I.; Khodorevskaya, R.P. Caspian Sea sturgeon fishery: A historic overview: Caspian Sea sturgeon fishery. J. Appl. Ichthyol. 2011, 27, 199–208. [Google Scholar] [CrossRef]
- Birstein, V.J.; Bemis, W.E.; Waldman, J.R. The threatened status of acipenseriform species: A summary. Environ. Biol. Fish. 1997, 48, 427–435. [Google Scholar] [CrossRef]
- Williot, P.; Sabeau, L.; Gessner, J.; Arlati, G.; Bronzi, P.; Gulyas, T.; Berni, P. Sturgeon farming in Western Europe: Recent developments and perspectives. Aquat. Living Resour. 2001, 14, 367–374. [Google Scholar] [CrossRef]
- Bronzi, P.; Rosenthal, H. Present and future sturgeon and caviar production and marketing: A global market overview. J. Appl. Ichthyol. 2014, 30, 1536–1546. [Google Scholar] [CrossRef]
- Parisi, G.; Terova, G.; Gasco, L.; Piccolo, G.; Roncarati, A.; Moretti, V.M.; Centoducati, G.; Gatta, P.P.; Pais, A. Current status and future perspectives of Italian finfish aquaculture. Rev. Fish Biol. Fish. 2014, 24, 15–73. [Google Scholar] [CrossRef]
- Bauer, O.N.; Pugachev, O.N.; Voronin, V.N. Study of parasites and diseases of sturgeons in Russia: A review. J. Appl. Ichthyol. 2002, 18, 420–429. [Google Scholar] [CrossRef]
- Santi, M.; Pastorino, P.; Foglini, C.; Righetti, M.; Pedron, C.; Prearo, M. A survey of bacterial infections in sturgeon farming in Italy. J. Appl. Ichthyol. 2018, 35, 275–282. [Google Scholar] [CrossRef]
- Chebanov, M.S.; Galich, E.V. Sturgeon hatchery manual. FAO Fish. Aquac. Tech. Pap. 2018, 558, 1–17. [Google Scholar]
- Mugetti, D.; Pastorino, P.; Menconi, V.; Pedron, C.; Prearo, M. The old and the new on viral disease in sturgeon, Pathogens 2020, 9, 146. Pathogens 2020, 9, 146. [Google Scholar] [CrossRef]
- Clouthier, S.; Anderson, E.; Kurath, G.; Breyta, R. Molecular systematics of sturgeon Nucleocytoplasmic Large DNA viruses. Mol. Phylogen. Evol. 2018, 128, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Pallandre, L.; Lesne, M.; de Boisséson, C.; Charrier, A.; Daniel, P.; Tragnan, A.; Debeuf, B.; Chesneau, V.; Bigarré, L. Genetic identification of two Acipenser iridovirus-European variants using high-resolution melting analysis. J. Virol. Methods 2019, 265, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Bigarré, L.; Lesne, M.; Lautraite, A.; Chesneau, V.; Leroux, A.; Jamin, M.; Boitard, P.M.; Toffan, A.; Prearo, M.; Labrut, S.; et al. Molecular identification of iridoviruses infecting various sturgeon species in Europe. J. Fish Dis. 2017, 40, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Mugetti, D.; Pastorino, P.; Menconi, V.; Messina, M.; Masoero, L.; Ceresa, L.; Pedron, C.; Prearo, M. Two new sturgeon species are susceptible to Acipenser Iridovirus European (AcIV-E) infection. Pathogens 2020, 9, 156. [Google Scholar] [CrossRef] [PubMed]
- Quaglio, F.; Bocus, R.; Delgado, M.L.; Gamberini, L.; Nobile, L.; Minelli, C.; Galuppi, A.; Restani, R. Aeromonas hydrophila infection in sterlet (Acipenser ruthenus) in a farm in the Po Valley. Boll. Soc. It. Patol. Ittica 2000, 28, 17–32. [Google Scholar]
- Colussi, S.; Gasparri, F.; Brunetti, R.; Ferrari, A.; Marturano, S.; Prearo, M. Aeromonas hydrophila infection in farmed Siberian sturgeon (Acipenser baeri). Ittiopatologia 2005, 2, 105–110. [Google Scholar]
- Ma, Z.; Yang, H.; Li, T.; Luo, L.; Gao, J. Isolation and identification of pathogenic Aeromonas veronii isolated from infected Siberian sturgeon (Acipenser baerii). Acta Microbiol. Sin. 2009, 49, 1289–1294. [Google Scholar] [CrossRef]
- Meng, Y.; Xiao, H.B.; Zeng, L.B. Isolation and identification of the hemorrhagic septicemia pathogen from Amur sturgeon, Acipenser schrenckii. J. Appl. Ichthyol. 2011, 27, 799–803. [Google Scholar] [CrossRef]
- Kayiş, Ş.; Er, A.; Kangel, P.; Kurtoğlu, İ.Z. Bacterial pathogens and health problems of Acipenser gueldenstaedtii and Acipenser baerii sturgeons reared in the eastern Black Sea region of Turkey. Iranian J. Vet. Res. 2017, 18, 18–24. [Google Scholar]
- Ture, M.; Ozcelep, T.; Akbulut, B.; Kutlu, I. Disease of Russian sturgeon (Acipenser gueldenstaedtii) caused by Aeromonas sp. Gen. Aquat. Org. 2018, 2, 43–47. [Google Scholar] [CrossRef]
- Vuillaume, A.; Brun, R.; Chene, P.; Sochon, E.; Lesel, R. First isolation of Yersinia ruckeri from sturgeon, Acipenser baerii Brandt, in south west of France. Bull. Eur. Ass. Fish Pathol. 1987, 7, 18–19. [Google Scholar]
- Li, S.; Wang, D.; Liu, H.; Lu, T. Isolation of Yersinia ruckeri strain H01 from farm-raised Amur Sturgeon Acipenser schrenckii in China. J. Aquat. An. Health 2013, 25, 9–14. [Google Scholar] [CrossRef]
- Li, S.; Zhang, J.; Cao, Y.; Wang, D.; Liu, H.; Lu, T. Transcriptome profiles of Amur sturgeon spleen in response to Yersinia ruckeri infection. Fish Shellfish Immunol. 2017, 70, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, R.; Gasparri, F.; Marturano, S.; Prearo, M. Pseudomonas fluorescens infection in farmed Siberian sturgeon (Acipenser baeri). Ittiopatologia 2006, 3, 221–226. [Google Scholar]
- Yang, J.; Tian, T.; Xiao, K.; Zeng, Q.; Tan, C.; Du, H. Pathogenic infection and immune-related gene expression of Chinese sturgeon (Acipenser sinensis) challenged by Citrobacter freundii. Dev. Comp. Immunol. 2021, 114, 103872. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Mu, Z.; Lv, X.; Jiang, X.; Zhou, J.; Guo, H.; Zhang, W.; Wang, B.; Lu, Y.; Wu, J.; et al. Pathogenicity of Plesiomonas shigelloides and Citrobacter freundii isolated from the endangered Chinese sturgeon (Acipenser sinensis). Microb. Pathog. 2022, 173, 105818. [Google Scholar] [CrossRef]
- Jiang, J.-Z.; Liu, Y.; Yan, L.-H.; Yan, Q.-G.; Wen, X.-T.; Cao, S.-J.; Huang, Y.; Huang, X.-B.; Ma, X.-P.; Han, X.-F.; et al. Identification and pathogenicity of Plesiomonas shigelloides from Acipenser dabryanus in China. Aquac. Res. 2021, 52, 2286–2293. [Google Scholar] [CrossRef]
- Pelkola, K.; Vennerström, P.; Viljamaa-Dirks, S.; Kuronen, H. Bacterial infections of farmed sturgeon in Finland. Fin. Food Saf. Auth. Evira 2012, 12, 122–127. [Google Scholar]
- Chinchilla, B.; Vázquez-Fernández, E.; Rebollada-Merino, A.; Pérez-Sancho, M.; Domínguez, L.; Rodríguez-Bertos, A. First detection of Flavobacterium psychrophilum in juvenile Siberian sturgeons (Acipenser baerii) and description of the pathological findings. J. Fish Dis. 2023, 46, 887–894. [Google Scholar] [CrossRef]
- Arlati, G.; Poliakova, L. Restoration of Adriatic sturgeon (Acipenser naccarii) in Italy: Situation and perspectives. In Biology, Conservation and Sustainable Development of Sturgeons; Carmona, R., Domezain, A., García-Gallego, M., Hernando, J.A., Rodríguez, F., Ruiz-Rejón, M., Eds.; Fish & Fisheries Series; Springer: Dordrecht, The Netherlands, 2009; Volume 29, pp. 237–245. [Google Scholar] [CrossRef]
- Boscari, E.; Congiu, L. The need for genetic support in restocking activities and ex situ conservation programmers: The case of the Adriatic sturgeon (Acipenser naccarii Bonaparte, 1836) in the Ticino River Park. J. Appl. Ichthyol. 2014, 30, 1416–1422. [Google Scholar] [CrossRef]
- Congiu, L.; Boscari, E.; Pagani, S.; Gazzola, M.; Bronzi, P. Resumption of natural reproduction of the Adriatic sturgeon in the River Po. Oryx 2021, 55, 816. [Google Scholar] [CrossRef]
- Barca, F.; Dalle Palle, S.; Schiavon, L.; Samassa, C.; Castaldelli, G.; Boscari, E.; Congiu, L. Characterization of captive breeders to preserve the residual genetic diversity of Adriatic sturgeon (Acipenser naccarii). Diversity 2022, 14, 829. [Google Scholar] [CrossRef]
- Axén, C.; Vendramin, N.; Toffan, A. Outbreak of mortality associated with Acipenser Iridovirus European (AcIV-E) detection in Siberian Sturgeon (Acipenser baerii) farmed in Sweden. Fishes 2018, 3, 42. [Google Scholar] [CrossRef]
- Radosavljević, V.; Milićević, V.; Maksimović-Zorić, J.; Veljović, L.; Nešić, K.; Pavlović, M.; Ljubojević Pelić, D.; Marković, Z. Sturgeon diseases in aquaculture. Arch. Vet. Med. 2019, 12, 5–20. [Google Scholar] [CrossRef]
- Billard, R.; Lecointre, G. Biology and conservation of sturgeon and paddlefish. Rev. Fish Biol. Fisheries 2001, 10, 355–392. [Google Scholar] [CrossRef]
- EUMOFA. Sturgeon meat and other by-products of caviar—production, trade, and consumption in and outside the EU. Publ. Off. Eur. Union Luxemb. 2023, 1–27. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bondavalli, F.; Schleicherová, D.; Pastorino, P.; Mugetti, D.; Pedron, C.; Prearo, M. Detection of Acipenser European Iridovirus (AcIV-E) in Sturgeon Farms in Northern Italy between 2021–2023. Viruses 2024, 16, 465. https://doi.org/10.3390/v16030465
Bondavalli F, Schleicherová D, Pastorino P, Mugetti D, Pedron C, Prearo M. Detection of Acipenser European Iridovirus (AcIV-E) in Sturgeon Farms in Northern Italy between 2021–2023. Viruses. 2024; 16(3):465. https://doi.org/10.3390/v16030465
Chicago/Turabian StyleBondavalli, Fabio, Dáša Schleicherová, Paolo Pastorino, Davide Mugetti, Claudio Pedron, and Marino Prearo. 2024. "Detection of Acipenser European Iridovirus (AcIV-E) in Sturgeon Farms in Northern Italy between 2021–2023" Viruses 16, no. 3: 465. https://doi.org/10.3390/v16030465
APA StyleBondavalli, F., Schleicherová, D., Pastorino, P., Mugetti, D., Pedron, C., & Prearo, M. (2024). Detection of Acipenser European Iridovirus (AcIV-E) in Sturgeon Farms in Northern Italy between 2021–2023. Viruses, 16(3), 465. https://doi.org/10.3390/v16030465