

HepG2BD: A Novel and Versatile Cell Line with Inducible HDV Replication and Constitutive HBV Expression
 , , , and
, , , and 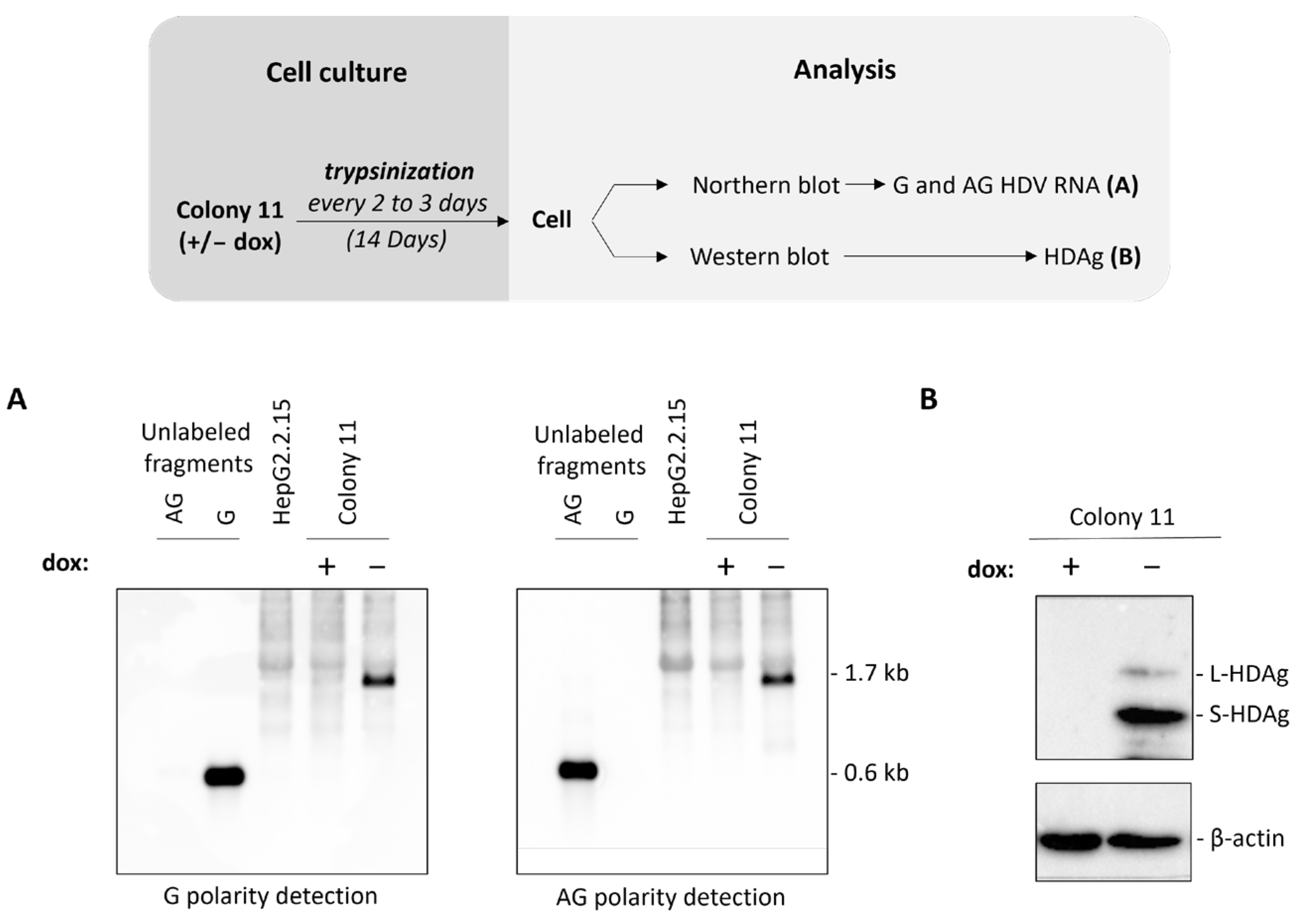{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagents
2.2. Plasmids and Cloning
2.3. Engineering of the Huh7-NTCP(L) Cell Line
2.4. HDV Infection Assay
2.5. Immunoprecipitation
2.6. ELISA
2.7. RT-qPCR and qPCR
2.8. Western Blot
2.9. Northern Blot
2.10. Contrast Phase and Confocal Immunofluorescence Microscopy
2.11. Antiviral Treatment
2.12. Statistical Analysis
3. Results
3.1. Generation of the HepG2BD Cell Line
3.2. HepG2BD Cell Line Allows Constitutive Expression of HBV and Inducible Replication of HDV
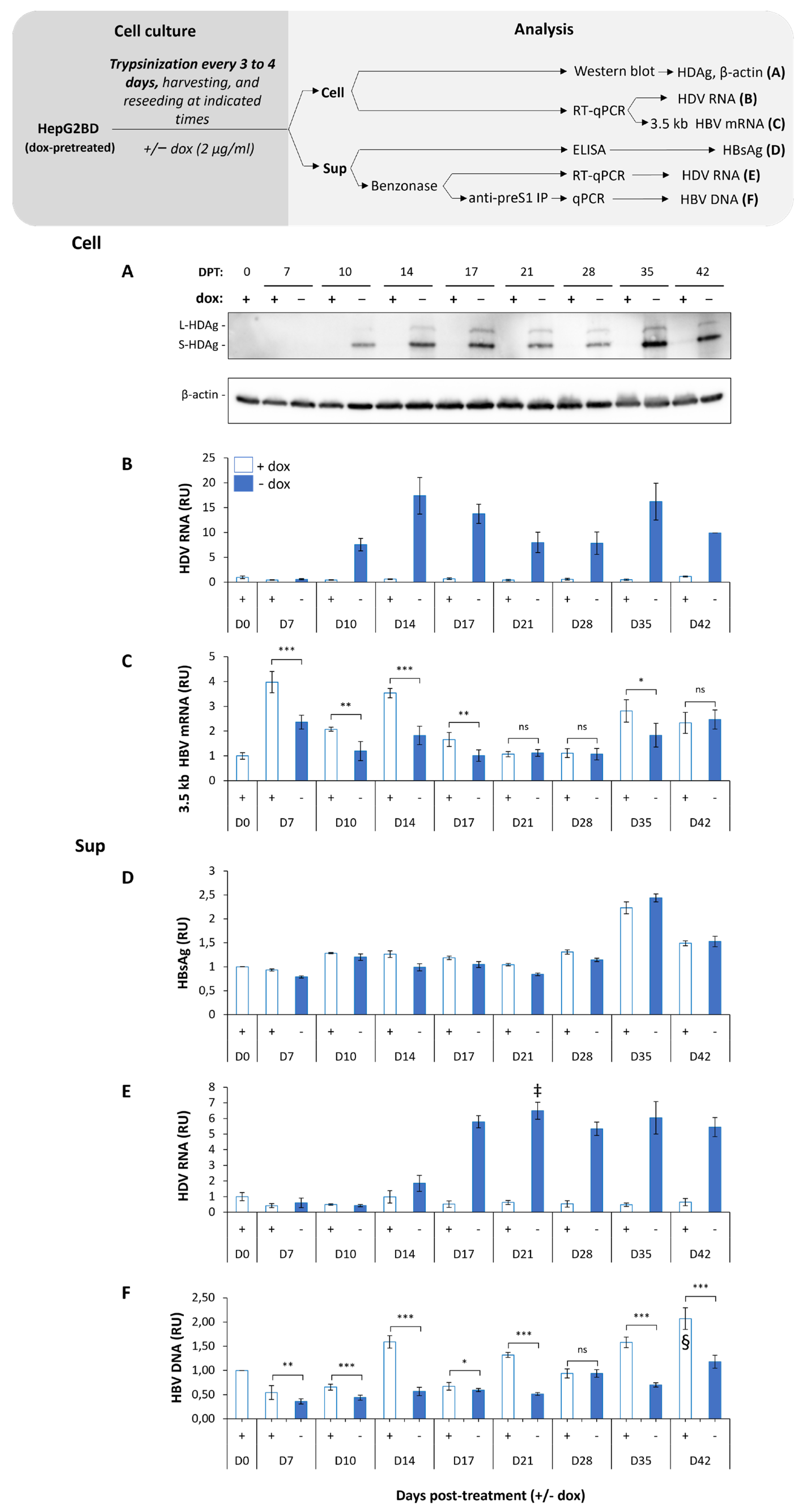
3.3. Ongoing HDV Replication in HepG2BD Is Efficiently Inhibited by the Addition of Doxycycline
3.4. Quiescence of HepG2BD Cells Enhances HDV Production
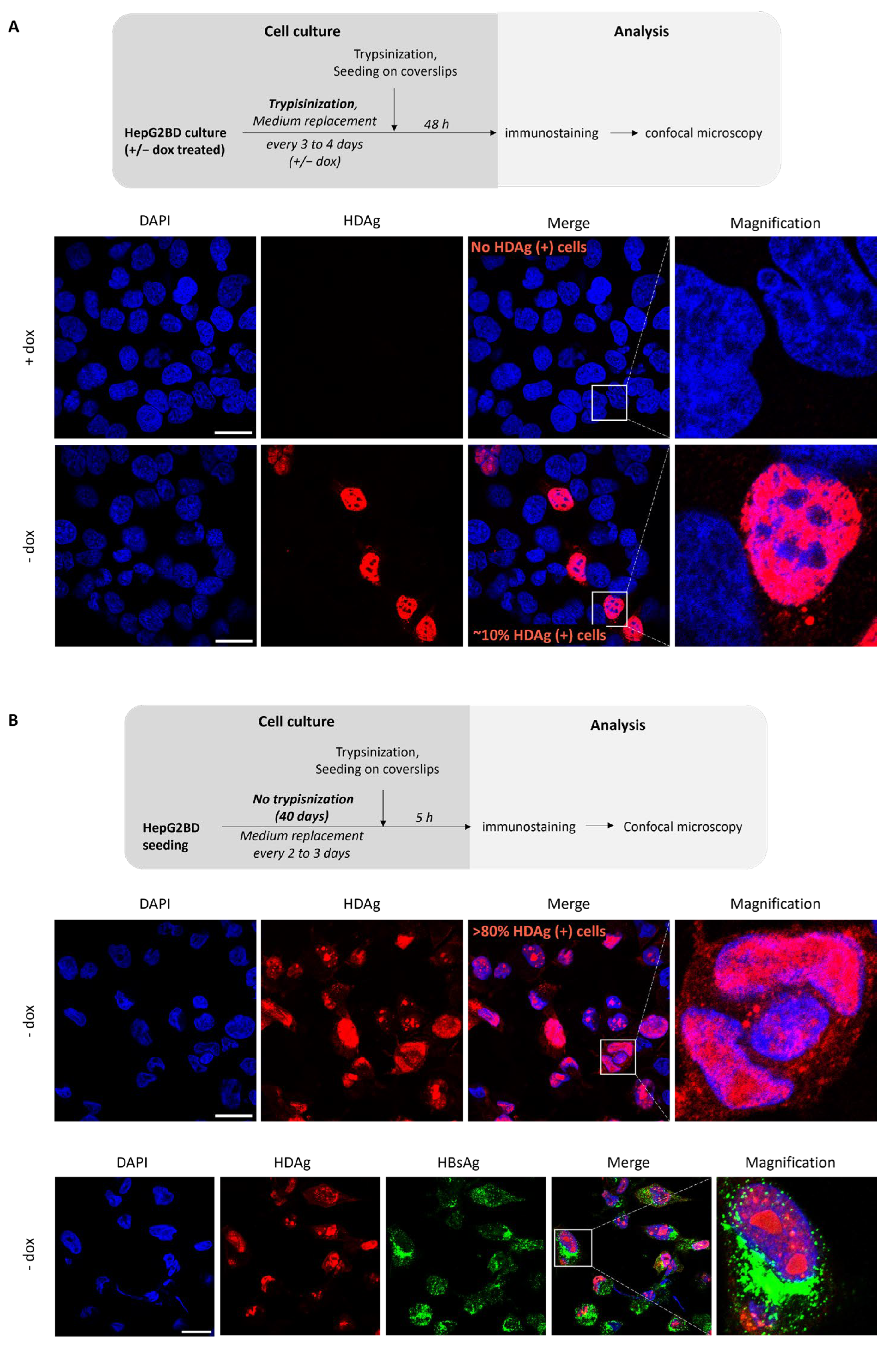
3.5. Effect of Culture Conditions on the Rate of HDV-Positive Cells
3.6. HepG2BD as a Tool for Antivirals Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Stephenne, J. Development and production aspects of a recombinant yeast-derived hepatitis B vaccine. Vaccine 1990, 8 (Suppl. S1), S69–S73; discussion S79–S80. [Google Scholar] [CrossRef]
- Polaris Observatory, C. Global prevalence, treatment, and prevention of hepatitis B virus infection in 2016: A modelling study. Lancet Gastroenterol. Hepatol. 2018, 3, 383–403. [Google Scholar] [CrossRef]
- Rizzetto, M.; Hamid, S.; Negro, F. The changing context of hepatitis D. J. Hepatol. 2021, 74, 1200–1211. [Google Scholar] [CrossRef]
- Fattovich, G.; Boscaro, S.; Noventa, F.; Pornaro, E.; Stenico, D.; Alberti, A.; Ruol, A.; Realdi, G. Influence of hepatitis delta virus infection on progression to cirrhosis in chronic hepatitis type B. J. Infect. Dis. 1987, 155, 931–935. [Google Scholar] [CrossRef]
- Fattovich, G.; Giustina, G.; Christensen, E.; Pantalena, M.; Zagni, I.; Realdi, G.; Schalm, S.W. Influence of hepatitis delta virus infection on morbidity and mortality in compensated cirrhosis type B. The European Concerted Action on Viral Hepatitis (Eurohep). Gut 2000, 46, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, A.; Gerlich, W.; Starke, C.; Wend, U.; Donner, A.; Sagir, A.; Heintges, T.; Häussinger, D. Treatment of chronic hepatitis delta with pegylated interferon-alpha2b. Liver Int. 2006, 26, 805–810. [Google Scholar] [CrossRef]
- Wedemeyer, H.; Yurdaydìn, C.; Dalekos, G.N.; Erhardt, A.; Çakaloğlu, Y.; Değertekin, H.; Gürel, S.; Zeuzem, S.; Zachou, K.; Bozkaya, H.; et al. Peginterferon plus adefovir versus either drug alone for hepatitis delta. N. Engl. J. Med. 2011, 364, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Sureau, C.; Romet-Lemonne, J.L.; Mullins, J.I.; Essex, M. Production of hepatitis B virus by a differentiated human hepatoma cell line after transfection with cloned circular HBV DNA. Cell 1986, 47, 37–47. [Google Scholar] [CrossRef]
- Sells, M.A.; Chen, M.L.; Acs, G. Production of hepatitis B virus particles in Hep G2 cells transfected with cloned hepatitis B virus DNA. Proc. Natl. Acad. Sci. USA 1987, 84, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Ladner, S.K.; Otto, M.J.; Barker, C.S.; Zaifert, K.; Wang, G.H.; Guo, J.T.; Seeger, C.; King, R.W. Inducible expression of human hepatitis B virus (HBV) in stably transfected hepatoblastoma cells: A novel system for screening potential inhibitors of HBV replication. Antimicrob. Agents Chemother. 1997, 41, 1715–1720. [Google Scholar] [CrossRef]
- Blanchet, M.; Sureau, C. Infectivity determinants of the hepatitis B virus pre-S domain are confined to the N-terminal 75 amino acid residues. J. Virol. 2007, 81, 5841–5849. [Google Scholar] [CrossRef]
- Blanchet, M.; Sureau, C. Analysis of the cytosolic domains of the hepatitis B virus envelope proteins for their function in viral particle assembly and infectivity. J. Virol. 2006, 80, 11935–11945. [Google Scholar] [CrossRef]
- Lempp, F.A.; Schlund, F.; Rieble, L.; Nussbaum, L.; Link, C.; Zhang, Z.; Ni, Y.; Urban, S. Recapitulation of HDV infection in a fully permissive hepatoma cell line allows efficient drug evaluation. Nat. Commun. 2019, 10, 2265. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Zhang, Z.; Engelskircher, L.; Verch, G.; Tu, T.; Lempp, F.A.; Urban, S. Generation and characterization of a stable cell line persistently replicating and secreting the human hepatitis delta virus. Sci. Rep. 2019, 9, 10021. [Google Scholar] [CrossRef] [PubMed]
- Bach, C.; Lucifora, J.; Delphin, M.; Heydmann, L.; Heuschkel, M.J.; Pons, C.; Goto, K.; Scheers, E.; Schuster, C.; Durantel, D.; et al. A stable hepatitis D virus-producing cell line for host target and drug discovery. Antivir. Res. 2023, 209, 105477. [Google Scholar] [CrossRef]
- Smith, J.R.; Maguire, S.; Davis, L.A.; Alexander, M.; Yang, F.; Chandran, S.; ffrench-Constant, C.; Pedersen, R.A. Robust, persistent transgene expression in human embryonic stem cells is achieved with AAVS1-targeted integration. Stem Cells 2008, 26, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Papapetrou, E.P.; Schambach, A. Gene Insertion Into Genomic Safe Harbors for Human Gene Therapy. Mol. Ther. 2016, 24, 678–684. [Google Scholar] [CrossRef]
- Dalvai, M.; Loehr, J.; Jacquet, K.; Huard, C.C.; Roques, C.; Herst, P.; Côté, J.; Doyon, Y. A Scalable Genome-Editing-Based Approach for Mapping Multiprotein Complexes in Human Cells. Cell Rep. 2015, 13, 621–633. [Google Scholar] [CrossRef]
- Chatel-Chaix, L.; Cortese, M.; Romero-Brey, I.; Bender, S.; Neufeldt, C.J.; Fischl, W.; Scaturro, P.; Schieber, N.; Schwab, Y.; Fischer, B.; et al. Dengue Virus Perturbs Mitochondrial Morphodynamics to Dampen Innate Immune Responses. Cell Host Microbe 2016, 20, 342–356. [Google Scholar] [CrossRef]
- Reya, T.; Duncan, A.W.; Ailles, L.; Domen, J.; Scherer, D.C.; Willert, K.; Hintz, L.; Nusse, R.; Weissman, I.L. A role for Wnt signalling in self-renewal of haematopoietic stem cells. Nature 2003, 423, 409–414. [Google Scholar] [CrossRef]
- Blanchet, M.; Sureau, C.; Labonte, P. Use of FDA approved therapeutics with hNTCP metabolic inhibitory properties to impair the HDV lifecycle. Antivir. Res. 2014, 106, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Boulon, R.; Blanchet, M.; Lemasson, M.; Vaillant, A.; Labonté, P. Characterization of the antiviral effects of REP 2139 on the HBV lifecycle in vitro. Antivir. Res. 2020, 183, 104853. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, M.; Sinnathamby, V.; Vaillant, A.; Labonte, P. Inhibition of HBsAg secretion by nucleic acid polymers in HepG2.2.15 cells. Antivir. Res. 2019, 164, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Perez-Vargas, J.; Amirache, F.; Boson, B.; Mialon, C.; Freitas, N.; Sureau, C.; Fusil, F.; Cosset, F.L. Enveloped viruses distinct from HBV induce dissemination of hepatitis D virus in vivo. Nat. Commun. 2019, 10, 2098. [Google Scholar] [CrossRef] [PubMed]
- Sureau, C.; Fournier-Wirth, C.; Maurel, P. Role of N glycosylation of hepatitis B virus envelope proteins in morphogenesis and infectivity of hepatitis delta virus. J. Virol. 2003, 77, 5519–5523. [Google Scholar] [CrossRef] [PubMed]
- Legrand, A.F.; Lucifora, J.; Lacombe, B.; Ménard, C.; Michelet, M.; Foca, A.; Abrial, P.; Salvetti, A.; Rivoire, M.; Lotteau, V.; et al. Farnesoid X receptor alpha ligands inhibit HDV in vitro replication and virion infectivity. Hepatol. Commun. 2023, 7, e0078. [Google Scholar] [CrossRef] [PubMed]
- Marcellin, P.; Zoulim, F.; Hézode, C.; Causse, X.; Roche, B.; Truchi, R.; Pauwels, A.; Ouzan, D.; Dumortier, J.; Pageaux, G.P.; et al. Effectiveness and Safety of Tenofovir Disoproxil Fumarate in Chronic Hepatitis B: A 3-Year, Prospective, Real-World Study in France. Dig. Dis. Sci. 2016, 61, 3072–3083. [Google Scholar] [CrossRef]
- Moini, M.; Fung, S. HBsAg Loss as a Treatment Endpoint for Chronic HBV Infection: HBV Cure. Viruses. 2022, 14, 657. [Google Scholar] [CrossRef]
- Koh, C.; Da, B.L.; Glenn, J.S. HBV/HDV Coinfection: A Challenge for Therapeutics. Clin. Liver Dis. 2019, 23, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Lucifora, J.; Delphin, M. Current knowledge on Hepatitis Delta Virus replication. Antivir. Res. 2020, 179, 104812. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Cornillez-Ty, C.; Lazinski, D.W. By inhibiting replication, the large hepatitis delta antigen can indirectly regulate amber/W editing and its own expression. J. Virol. 2004, 78, 8120–8134. [Google Scholar] [CrossRef] [PubMed]
- Pollicino, T.; Raffa, G.; Santantonio, T.; Gaeta, G.B.; Iannello, G.; Alibrandi, A.; Squadrito, G.; Cacciola, I.; Calvi, C.; Colucci, G.; et al. Replicative and transcriptional activities of hepatitis B virus in patients coinfected with hepatitis B and hepatitis delta viruses. J. Virol. 2011, 85, 432–439. [Google Scholar] [CrossRef]
- Lucifora, J.; Alfaiate, D.; Pons, C.; Michelet, M.; Ramirez, R.; Fusil, F.; Amirache, F.; Rossi, A.; Legrand, A.F.; Charles, E.; et al. Hepatitis D virus interferes with hepatitis B virus RNA production via interferon-dependent and-independent mechanisms. J. Hepatol. 2023, 78, 958–970. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanchet, M.; Angelo, L.; Tétreault, Y.; Khabir, M.; Sureau, C.; Vaillant, A.; Labonté, P. HepG2BD: A Novel and Versatile Cell Line with Inducible HDV Replication and Constitutive HBV Expression. Viruses 2024, 16, 532. https://doi.org/10.3390/v16040532
Blanchet M, Angelo L, Tétreault Y, Khabir M, Sureau C, Vaillant A, Labonté P. HepG2BD: A Novel and Versatile Cell Line with Inducible HDV Replication and Constitutive HBV Expression. Viruses. 2024; 16(4):532. https://doi.org/10.3390/v16040532
Chicago/Turabian StyleBlanchet, Matthieu, Léna Angelo, Yasmine Tétreault, Marwa Khabir, Camille Sureau, Andrew Vaillant, and Patrick Labonté. 2024. "HepG2BD: A Novel and Versatile Cell Line with Inducible HDV Replication and Constitutive HBV Expression" Viruses 16, no. 4: 532. https://doi.org/10.3390/v16040532