
The Susceptibility of Chickens to Zika Virus: A Comprehensive Study on Age-Dependent Infection Dynamics and Host Responses

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Virus Propagation, Inactivation, and Quantification
2.3. Infection Experiments
2.4. Viral RNA Extraction
2.5. Total RNA Extraction from Spleen
2.6. Host Gene qPCR
2.7. Viral qRT-PCR
2.8. Serological Assays
3. Results
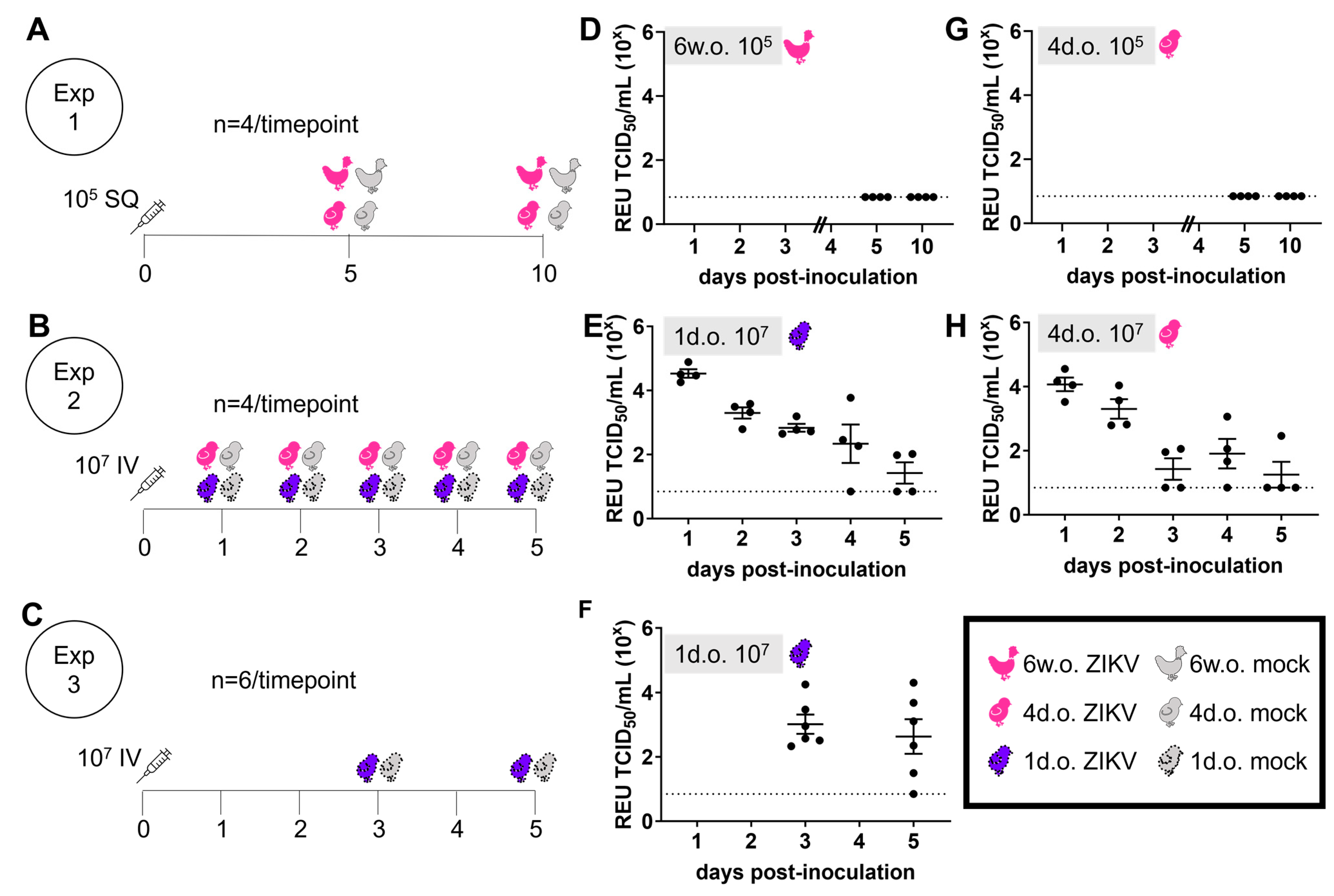
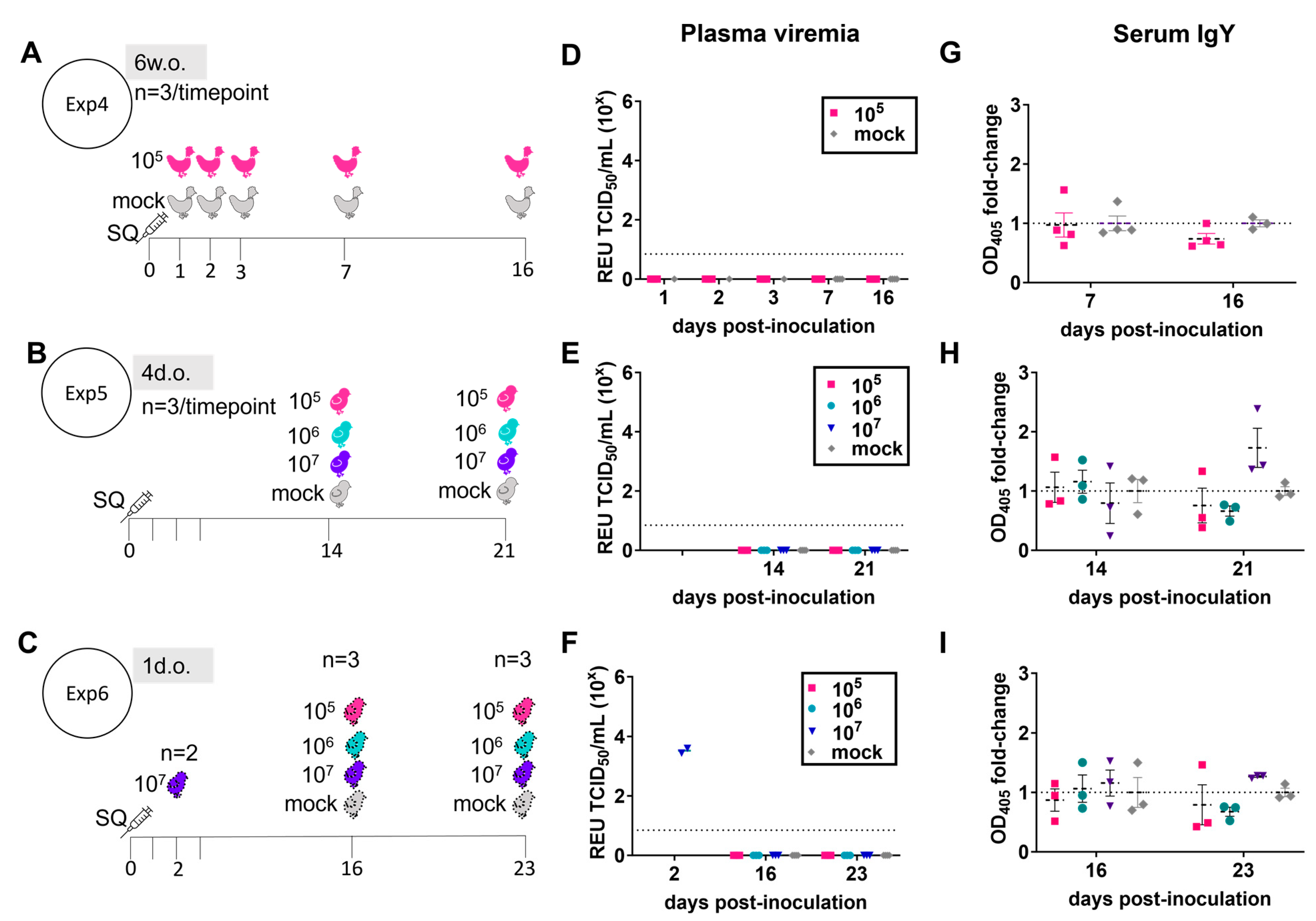
3.1. ZIKV Does Not Create a Productive Infection in Juvenile Chickens
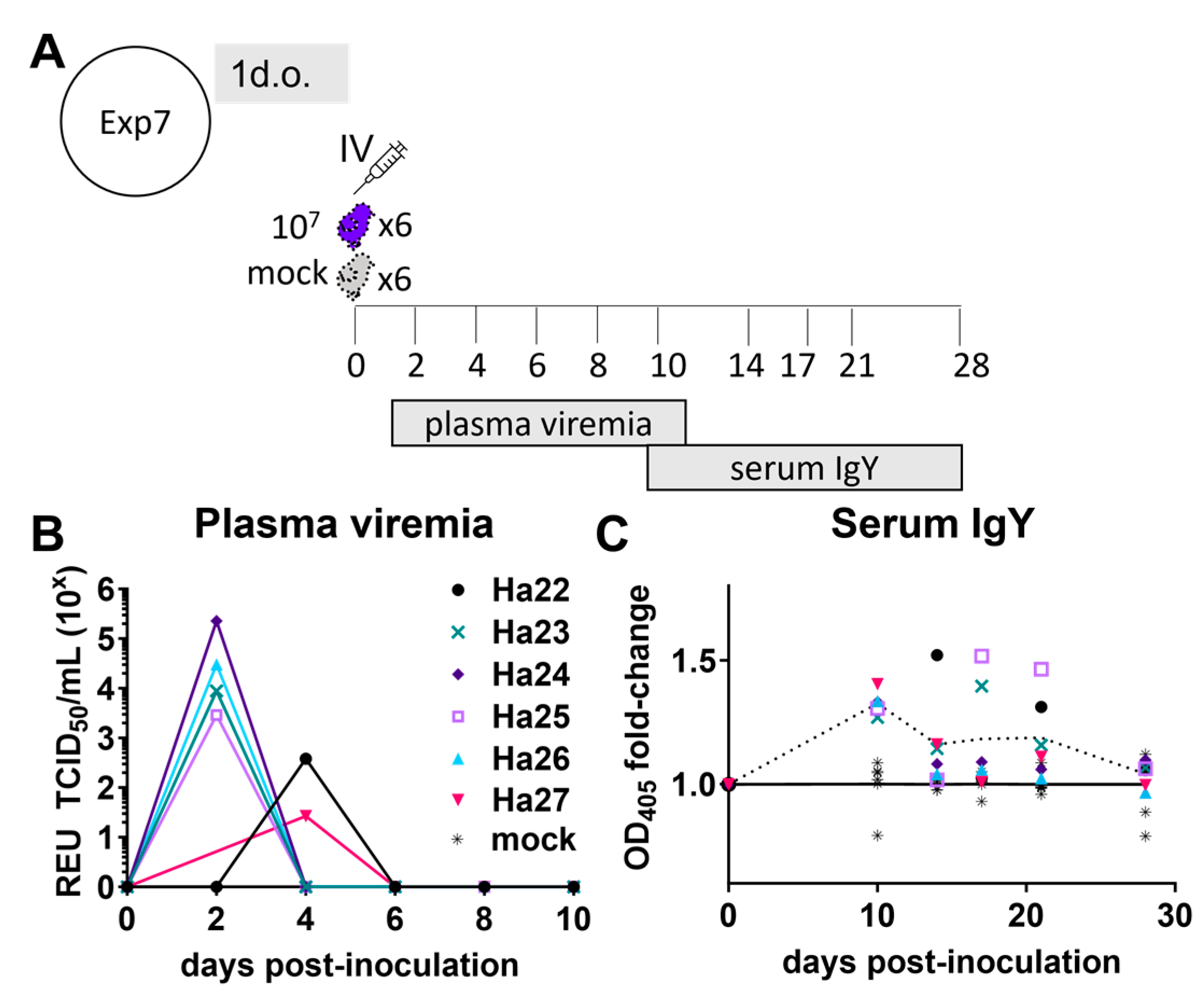
3.2. ZIKV Characterization in 1-Day-Old Chickens
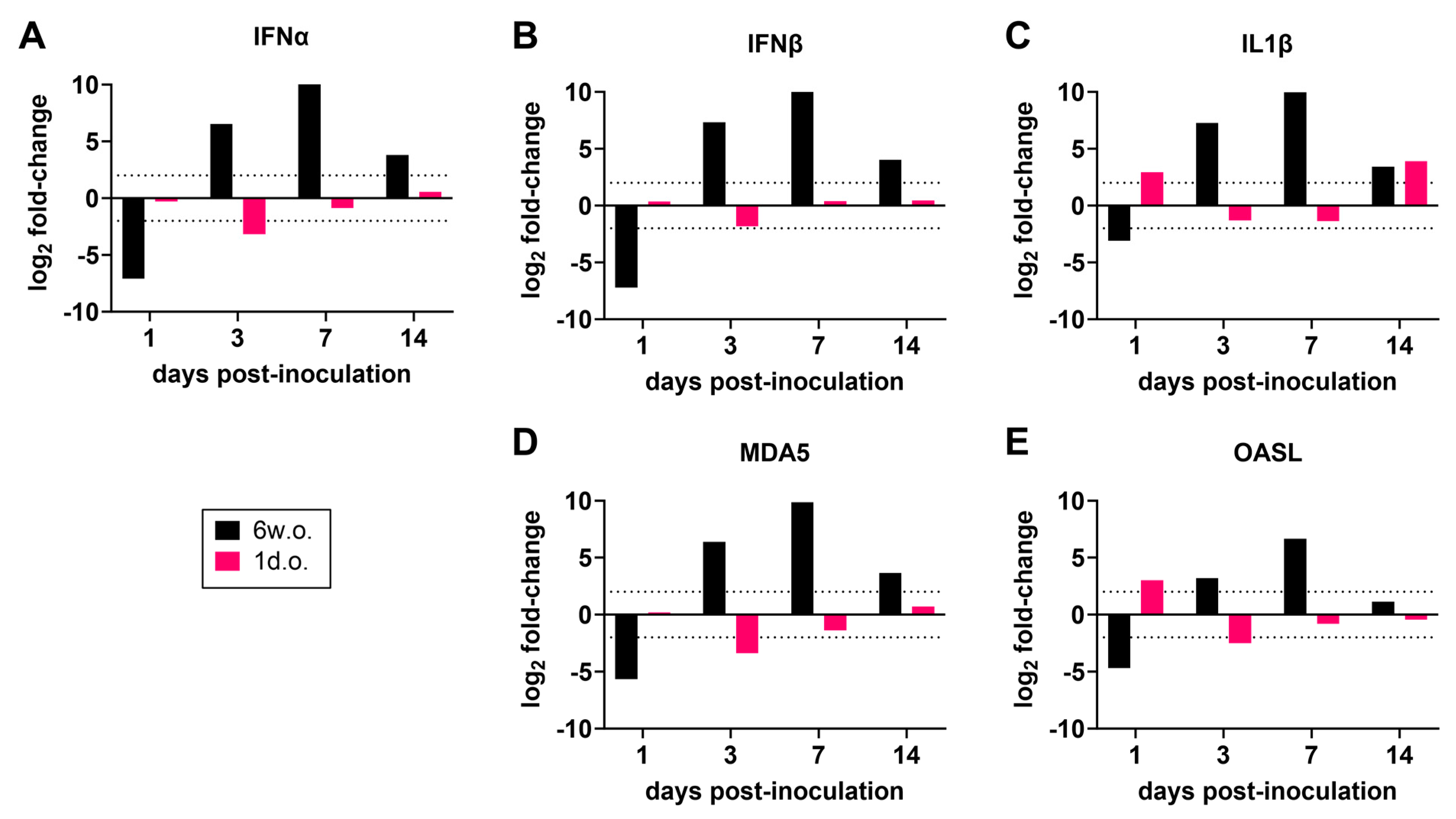
3.3. Differential Gene Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Postler, T.S.; Beer, M.; Blitvich, B.J.; Bukh, J.; de Lamballerie, X.; Drexler, J.F.; Imrie, A.; Kapoor, A.; Karganova, G.G.; Lemey, P.; et al. Renaming of the genus Flavivirus to Orthoflavivirus and extension of binomial species names within the family Flaviviridae. Arch. Virol. 2023, 168, 224. [Google Scholar] [CrossRef] [PubMed]
- Brasil, P.; Calvet, G.A.; Siqueira, A.M.; Wakimoto, M.; de Sequeira, P.C.; Nobre, A.; Quintana Mde, S.; Mendonca, M.C.; Lupi, O.; de Souza, R.V.; et al. Zika Virus Outbreak in Rio de Janeiro, Brazil: Clinical Characterization, Epidemiological and Virological Aspects. PLoS Negl. Trop. Dis. 2016, 10, e0004636. [Google Scholar] [CrossRef] [PubMed]
- Lessler, J.; Chaisson, L.H.; Kucirka, L.M.; Bi, Q.; Grantz, K.; Salje, H.; Carcelen, A.C.; Ott, C.T.; Sheffield, J.S.; Ferguson, N.M.; et al. Assessing the global threat from Zika virus. Science 2016, 353, aaf8160. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Filho Dde, B.; Martelli, C.M.; Ximenes, R.A.; Araújo, T.V.; Rocha, M.A.; Ramos, R.C.; Dhalia, R.; França, R.F.; Marques Júnior, E.T.; Rodrigues, L.C. Initial Description of the Presumed Congenital Zika Syndrome. Am. J. Public Health 2016, 106, 598–600. [Google Scholar] [CrossRef] [PubMed]
- França, G.V.; Schuler-Faccini, L.; Oliveira, W.K.; Henriques, C.M.; Carmo, E.H.; Pedi, V.D.; Nunes, M.L.; Castro, M.C.; Serruya, S.; Silveira, M.F.; et al. Congenital Zika virus syndrome in Brazil: A case series of the first 1501 livebirths with complete investigation. Lancet 2016, 388, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; Choi, G.K.; Yip, C.C.; Cheng, V.C.; Yuen, K.Y. Zika fever and congenital Zika syndrome: An unexpected emerging arboviral disease. J. Infect. 2016, 72, 507–524. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Zika Virus. Available online: https://www.who.int/news-room/fact-sheets/detail/zika-virus (accessed on 23 February 2024).
- Sah, R.; Mohanty, A.; Paul, D.; Padhi, B.K. Recent outbreak of zika virus in India amid ongoing COVID-19 and monkeypox outbreak: A call for action. Int. J. Surg. 2023, 109, 601–603. [Google Scholar] [CrossRef]
- Gubler, D.J.; Vasilakis, N.; Musso, D. History and Emergence of Zika Virus. J. Infect. Dis. 2017, 216 (Suppl. 10), S860–S867. [Google Scholar] [CrossRef]
- Pielnaa, P.; Al-Saadawe, M.; Saro, A.; Dama, M.F.; Zhou, M.; Huang, Y.; Huang, J.; Xia, Z. Zika virus-spread, epidemiology, genome, transmission cycle, clinical manifestation, associated challenges, vaccine and antiviral drug development. Virology 2020, 543, 34–42. [Google Scholar] [CrossRef]
- Hasan, S.; Saeed, S.; Panigrahi, R.; Choudhary, P. Zika Virus: A Global Public Health Menace: A Comprehensive Update. J. Int. Soc. Prev. Community Dent. 2019, 9, 316–327. [Google Scholar] [CrossRef]
- Benzarti, E.; Garigliany, M. In Vitro and In Vivo Models to Study the Zoonotic Mosquito-Borne Usutu Virus. Viruses 2020, 12, 1116. [Google Scholar] [CrossRef] [PubMed]
- Duggal, N.K.; Langwig, K.E.; Ebel, G.D.; Brault, A.C. On the Fly: Interactions Between Birds, Mosquitoes, and Environment That Have Molded West Nile Virus Genomic Structure Over Two Decades. J. Med. Entomol. 2019, 56, 1467–1474. [Google Scholar] [CrossRef] [PubMed]
- Curren, E.J.; Lindsey, N.P.; Fischer, M.; Hills, S.L. St. Louis Encephalitis Virus Disease in the United States, 2003–2017. Am. J. Trop. Med. Hyg. 2018, 99, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Aliota, M.T.; Peinado, S.A.; Osorio, J.E.; Bartholomay, L.C. Culex pipiens and Aedes triseriatus Mosquito Susceptibility to Zika Virus. Emerg. Infect. Dis. 2016, 22, 1857–1859. [Google Scholar] [CrossRef] [PubMed]
- Lourenço-de-Oliveira, R.; Marques, J.T.; Sreenu, V.B.; Atyame Nten, C.; Aguiar, E.; Varjak, M.; Kohl, A.; Failloux, A.B. Culex quinquefasciatus mosquitoes do not support replication of Zika virus. J. Gen. Virol. 2018, 99, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Quiroga, D.; Ramírez-Medina, M.; Gutiérrez-Ortega, A.; Elizondo-Quiroga, A.; Muñoz-Medina, J.E.; Sánchez-Tejeda, G.; González-Acosta, C.; Correa-Morales, F. Vector competence of Aedes aegypti and Culex quinquefasciatus from the metropolitan area of Guadalajara, Jalisco, Mexico for Zika virus. Sci. Rep. 2019, 9, 16955. [Google Scholar] [CrossRef] [PubMed]
- van den Hurk, A.F.; Hall-Mendelin, S.; Jansen, C.C.; Higgs, S. Zika virus and Culex quinquefasciatus mosquitoes: A tenuous link. Lancet Infect. Dis. 2017, 17, 1014–1016. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, H.J.; Dimopoulos, G. Detailed Analyses of Zika Virus Tropism in Culex quinquefasciatus Reveal Systemic Refractoriness. mBio 2020, 11, e01765-20. [Google Scholar] [CrossRef] [PubMed]
- Viveiros-Rosa, S.G.; Regis, E.G.; Santos, W.C. Vector competence of Culex mosquitoes (Diptera: Culicidae) in Zika virus transmission: An integrative review. Rev. Panam. De Salud Publica=Pan Am. J. Public Health 2020, 44, e7. [Google Scholar] [CrossRef]
- Hart, C.E.; Roundy, C.M.; Azar, S.R.; Huang, J.H.; Yun, R.; Reynolds, E.; Leal, G.; Nava, M.R.; Vela, J.; Stark, P.M.; et al. Zika Virus Vector Competency of Mosquitoes, Gulf Coast, United States. Emerg. Infect. Dis. 2017, 23, 559–560. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Mogi, M.; Armbruster, P.A.; Tuno, N.; Aranda, C.; Yong, H.S. The Climate Range Expansion of Aedes albopictus (Diptera: Culicidae) in Asia Inferred From the Distribution of Albopictus Subgroup Species of Aedes (Stegomyia). J. Med. Entomol. 2017, 54, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.; Shearer, F.M.; Brady, O.J.; Messina, J.P.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; et al. The global compendium of Aedes aegypti and Ae. albopictus occurrence. Sci. Data 2015, 2, 150035. [Google Scholar] [CrossRef] [PubMed]
- Tuten, H.C.; Bridges, W.C.; Paul, K.S.; Adler, P.H. Blood-feeding ecology of mosquitoes in zoos. Med. Vet. Entomol. 2012, 26, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.C.; Silva, N.N.; Marques, G.R.; Brito, M. Host-feeding patterns of potential human disease vectors in the Paraiba Valley region, State of Sao Paulo, Brazil. J. Vector Ecol. 2003, 28, 74–78. [Google Scholar] [PubMed]
- Garcia-Rejon, J.E.; Navarro, J.C.; Cigarroa-Toledo, N.; Baak-Baak, C.M. An Updated Review of the Invasive Aedes albopictus in the Americas; Geographical Distribution, Host Feeding Patterns, Arbovirus Infection, and the Potential for Vertical Transmission of Dengue Virus. Insects 2021, 12, 967. [Google Scholar] [CrossRef] [PubMed]
- Little, E.A.H.; Harriott, O.T.; Akaratovic, K.I.; Kiser, J.P.; Abadam, C.F.; Shepard, J.J.; Molaei, G. Host interactions of Aedes albopictus, an invasive vector of arboviruses, in Virginia, USA. PLoS Negl. Trop. Dis. 2021, 15, e0009173. [Google Scholar] [CrossRef] [PubMed]
- Ponlawat, A.; Harrington, L.C. Blood feeding patterns of Aedes aegypti and Aedes albopictus in Thailand. J. Med. Entomol. 2005, 42, 844–849. [Google Scholar] [CrossRef]
- Sene, N.M.; Diouf, B.; Gaye, A.; Ndiaye, E.H.; Ngom, E.M.; Gueye, A.; Seck, F.; Diagne, C.T.; Dia, I.; Diallo, D.; et al. Blood Feeding Patterns of Aedes aegypti Populations in Senegal. Am. J. Trop. Med. Hyg. 2022, 106, 1402–1405. [Google Scholar] [CrossRef]
- Richards, S.L.; Ponnusamy, L.; Unnasch, T.R.; Hassan, H.K.; Apperson, C.S. Host-feeding patterns of Aedes albopictus (Diptera: Culicidae) in relation to availability of human and domestic animals in suburban landscapes of central North Carolina. J. Med. Entomol. 2006, 43, 543–551. [Google Scholar] [CrossRef]
- Haby, M.M.; Pinart, M.; Elias, V.; Reveiz, L. Prevalence of asymptomatic Zika virus infection: A systematic review. Bull. World Health Organ. 2018, 96, 402–413D. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika virus outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef]
- Subissi, L.; Daudens-Vaysse, E.; Cassadou, S.; Ledrans, M.; Bompard, P.; Gustave, J.; Aubry, M.; Cao-Lormeau, V.M.; Mallet, H.P. Revising rates of asymptomatic Zika virus infection based on sentinel surveillance data from French Overseas Territories. Int. J. Infect. Dis. 2017, 65, 116–118. [Google Scholar] [CrossRef]
- World Health Organization Zika Epidemiology Update July 2019. Available online: https://www.who.int/publications/m/item/zika-epidemiology-update (accessed on 2 March 2024).
- Kleber de Oliveira, W.; Cortez-Escalante, J.; De Oliveira, W.T.G.H.; do Carmo, G.M.I.; Henriques, C.M.P.; Coelho, G.E.; Araújo de França, G.V. Increase in Reported Prevalence of Microcephaly in Infants Born to Women Living in Areas with Confirmed Zika Virus Transmission During the First Trimester of Pregnancy—Brazil, 2015. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Lazear, H.M.; Diamond, M.S. Zika Virus: New Clinical Syndromes and Its Emergence in the Western Hemisphere. J. Virol. 2016, 90, 4864–4875. [Google Scholar] [CrossRef] [PubMed]
- Brasil, P.; Pereira, J.P., Jr.; Moreira, M.E.; Ribeiro Nogueira, R.M.; Damasceno, L.; Wakimoto, M.; Rabello, R.S.; Valderramos, S.G.; Halai, U.A.; Salles, T.S.; et al. Zika Virus Infection in Pregnant Women in Rio de Janeiro. N. Engl. J. Med. 2016, 375, 2321–2334. [Google Scholar] [CrossRef]
- Brasil, P.; Sequeira, P.C.; Freitas, A.D.; Zogbi, H.E.; Calvet, G.A.; de Souza, R.V.; Siqueira, A.M.; de Mendonca, M.C.; Nogueira, R.M.; de Filippis, A.M.; et al. Guillain-Barre syndrome associated with Zika virus infection. Lancet 2016, 387, 1482. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.M.; Blake, A.; Mons, S.; Lastere, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barre Syndrome outbreak associated with Zika virus infection in French Polynesia: A case-control study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef]
- Wilder-Smith, A.; Brickley, E.B.; Ximenes, R.A.A.; Miranda-Filho, D.B.; Turchi Martelli, C.M.; Solomon, T.; Jacobs, B.C.; Pardo, C.A.; Osorio, L.; Parra, B.; et al. The legacy of ZikaPLAN: A transnational research consortium addressing Zika. Glob. Health Action. 2021, 14, 2008139. [Google Scholar] [CrossRef]
- Culjat, M.; Darling, S.E.; Nerurkar, V.R.; Ching, N.; Kumar, M.; Min, S.K.; Wong, R.; Grant, L.; Melish, M.E. Clinical and Imaging Findings in an Infant With Zika Embryopathy. Clin. Infect. Dis. 2016, 63, 805–811. [Google Scholar]
- Paixao, E.S.; Cardim, L.L.; Costa, M.C.N.; Brickley, E.B.; de Carvalho-Sauer, R.C.O.; Carmo, E.H.; Andrade, R.F.S.; Rodrigues, M.S.; Veiga, R.V.; Costa, L.C.; et al. Mortality from Congenital Zika Syndrome—Nationwide Cohort Study in Brazil. N. Engl. J. Med. 2022, 386, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Neelam, V.; Woodworth, K.R.; Chang, D.J.; Roth, N.M.; Reynolds, M.R.; Akosa, A.; Carr, C.P.; Anderson, K.N.; Mulkey, S.B.; DeBiasi, R.L.; et al. Outcomes up to age 36 months after congenital Zika virus infection-U.S. states. Pediatr. Res. 2024, 95, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Sobral da Silva, P.F.; Eickmann, S.H.; Arraes de Alencar Ximenes, R.; Ramos Montarroyos, U.; de Carvalho Lima, M.; Turchi Martelli, C.M.; Velho Barreto de Araújo, T.; Brickley, E.B.; Cunha Rodrigues, L.; Lima da Silva Pastich Gonçalves, F.C.; et al. Pediatric neurodevelopment by prenatal Zika virus exposure: A cross-sectional study of the Microcephaly Epidemic Research Group Cohort. BMC Pediatr. 2020, 20, 472. [Google Scholar] [CrossRef] [PubMed]
- Soriano-Arandes, A.; Frick, M.A.; García López-Hortelano, M.; Sulleiro, E.; Rodó, C.; Sánchez-Seco, M.P.; Cabrera-Lafuente, M.; Suy, A.; De la Calle, M.; Santos, M.; et al. Clinical Outcomes of a Zika Virus Mother-Child Pair Cohort in Spain. Pathogens 2020, 9, 352. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, A.C.; Toth, D.; Ridenour, T.; Lima Nóbrega, L.; Borba Firmino, R.; Marques da Silva, C.; Carvalho, P.; Marques, D.; Okoniewski, K.; Ventura, L.O.; et al. Developmental Outcomes Among Young Children With Congenital Zika Syndrome in Brazil. JAMA Netw. Open 2020, 3, e204096. [Google Scholar] [CrossRef] [PubMed]
- Peçanha, P.M.; Gomes Junior, S.C.; Pone, S.M.; Pone, M.; Vasconcelos, Z.; Zin, A.; Vilibor, R.H.H.; Costa, R.P.; Meio, M.; Nielsen-Saines, K.; et al. Neurodevelopment of children exposed intra-uterus by Zika virus: A case series. PLoS ONE 2020, 15, e0229434. [Google Scholar] [CrossRef] [PubMed]
- Valentine, M.J.; Murdock, C.C.; Kelly, P.J. Sylvatic cycles of arboviruses in non-human primates. Parasites Vectors 2019, 12, 463. [Google Scholar] [CrossRef]
- Polen, K.D.; Gilboa, S.M.; Hills, S.; Oduyebo, T.; Kohl, K.S.; Brooks, J.T.; Adamski, A.; Simeone, R.M.; Walker, A.T.; Kissin, D.M.; et al. Update: Interim Guidance for Preconception Counseling and Prevention of Sexual Transmission of Zika Virus for Men with Possible Zika Virus Exposure—United States, August 2018. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 868–871. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Filho, E.F.; Carneiro, I.O.; Fischer, C.; Kuhne, A.; Postigo-Hidalgo, I.; Ribas, J.R.L.; Schumann, P.; Nowak, K.; Gogarten, J.F.; de Lamballerie, X.; et al. Evidence against Zika virus infection of pets and peri-domestic animals in Latin America and Africa. J. Gen. Virol. 2022, 103, 001709. [Google Scholar] [CrossRef]
- Han, B.A.; Majumdar, S.; Calmon, F.P.; Glicksberg, B.S.; Horesh, R.; Kumar, A.; Perer, A.; von Marschall, E.B.; Wei, D.; Mojsilovic, A.; et al. Confronting data sparsity to identify potential sources of Zika virus spillover infection among primates. Epidemics 2019, 27, 59–65. [Google Scholar] [CrossRef]
- Seferovic, M.; Sanchez-San Martin, C.; Tardif, S.D.; Rutherford, J.; Castro, E.C.C.; Li, T.; Hodara, V.L.; Parodi, L.M.; Giavedoni, L.; Layne-Colon, D.; et al. Experimental Zika Virus Infection in the Pregnant Common Marmoset Induces Spontaneous Fetal Loss and Neurodevelopmental Abnormalities. Sci. Rep. 2018, 8, 6851. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Sanchez-San Martin, C.; Bouquet, J.; Li, T.; Yagi, S.; Tamhankar, M.; Hodara, V.L.; Parodi, L.M.; Somasekar, S.; Yu, G.; et al. Experimental Zika Virus Inoculation in a New World Monkey Model Reproduces Key Features of the Human Infection. Sci. Rep. 2017, 7, 17126. [Google Scholar] [CrossRef]
- Berry, N.; Ferguson, D.; Ham, C.; Hall, J.; Jenkins, A.; Giles, E.; Devshi, D.; Kempster, S.; Rose, N.; Dowall, S.; et al. High susceptibility, viral dynamics and persistence of South American Zika virus in New World monkey species. Sci. Rep. 2019, 9, 14495. [Google Scholar] [CrossRef]
- Li, X.F.; Dong, H.L.; Huang, X.Y.; Qiu, Y.F.; Wang, H.J.; Deng, Y.Q.; Zhang, N.N.; Ye, Q.; Zhao, H.; Liu, Z.Y.; et al. Characterization of a 2016 Clinical Isolate of Zika Virus in Non-human Primates. EBioMedicine 2016, 12, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Osuna, C.E.; Lim, S.Y.; Deleage, C.; Griffin, B.D.; Stein, D.; Schroeder, L.T.; Omange, R.W.; Best, K.; Luo, M.; Hraber, P.T.; et al. Zika viral dynamics and shedding in rhesus and cynomolgus macaques. Nat. Med. 2016, 22, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Dudley, D.M.; Aliota, M.T.; Mohr, E.L.; Weiler, A.M.; Lehrer-Brey, G.; Weisgrau, K.L.; Mohns, M.S.; Breitbach, M.E.; Rasheed, M.N.; Newman, C.M.; et al. A rhesus macaque model of Asian-lineage Zika virus infection. Nat. Commun. 2016, 7, 12204. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.D.; Perez-Sautu, U.; Wiley, M.R.; Miller, L.J.; Kimmel, A.E.; Principe, L.M.; Wollen-Roberts, S.E.; Shamblin, J.D.; Valdez, S.M.; Cazares, L.H.; et al. Modeling mosquito-borne and sexual transmission of Zika virus in an enzootic host, the African green monkey. PLoS Negl. Trop. Dis. 2020, 14, e0008107. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.J.; Nasar, F.; Schellhase, C.W.; Norris, S.L.; Kimmel, A.E.; Valdez, S.M.; Wollen-Roberts, S.E.; Shamblin, J.D.; Sprague, T.R.; Lugo-Roman, L.A.; et al. Zika Virus Infection in Syrian Golden Hamsters and Strain 13 Guinea Pigs. Am. J. Trop. Med. Hyg. 2018, 98, 864–867. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Krause, K.K.; Azouz, F.; Nakano, E.; Nerurkar, V.R. A guinea pig model of Zika virus infection. Virol. J. 2017, 14, 75. [Google Scholar] [CrossRef]
- Li, S.; Armstrong, N.; Zhao, H.; Hou, W.; Liu, J.; Chen, C.; Wan, J.; Wang, W.; Zhong, C.; Liu, C.; et al. Zika Virus Fatally Infects Wild Type Neonatal Mice and Replicates in Central Nervous System. Viruses 2018, 10, 49. [Google Scholar] [CrossRef]
- Paul, A.M.; Acharya, D.; Neupane, B.; Thompson, E.A.; Gonzalez-Fernandez, G.; Copeland, K.M.; Garrett, M.L.; Liu, H.; Lopez, M.E.; de Cruz, M.; et al. Congenital Zika Virus Infection in Immunocompetent Mice Causes Postnatal Growth Impediment and Neurobehavioral Deficits. Front. Microbiol. 2018, 9, 2028. [Google Scholar] [CrossRef]
- Miller, M.R.; Fagre, A.C.; Clarkson, T.C.; Markle, E.D.; Foy, B.D. Three Immunocompetent Small Animal Models That Do Not Support Zika Virus Infection. Pathogens 2021, 10, 971. [Google Scholar] [CrossRef]
- Ragan, I.K.; Blizzard, E.L.; Gordy, P.; Bowen, R.A. Investigating the Potential Role of North American Animals as Hosts for Zika Virus. Vector Borne Zoonotic Dis. 2017, 17, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Ambagala, A.; Truong, T.; Cottam-Birt, C.; Berhane, Y.; Gerdts, V.; Karniychuk, U.; Safronetz, D.; Babiuk, S. Susceptibility of Chicken Embryos, Sheep, Cattle, Pigs, and Chickens to Zika Virus Infection. Front. Vet. Sci. 2020, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.K.; Chanas, A.C.; Shockley, P.; Squires, E.J.; Gardner, P.; Wallace, C.; Simpson, D.I.; Bowen, E.T.; Platt, G.S.; Way, H.; et al. Arbovirus isolations from, and serological studies on, wild and domestic vertebrates from Kano Plain, Kenya. Trans. R. Soc. Trop. Med. Hyg. 1977, 71, 512–517. [Google Scholar] [CrossRef]
- Olson, J.G.; Ksiazek, T.G.; Gubler, D.J.; Lubis, S.I.; Simanjuntak, G.; Lee, V.H.; Nalim, S.; Juslis, K.; See, R. A survey for arboviral antibodies in sera of humans and animals in Lombok, Republic of Indonesia. Ann. Trop. Med. Parasitol. 1983, 77, 131–137. [Google Scholar] [CrossRef]
- Li, F.; Zhou, J.Z.; Zhou, L.; Fu, S.H.; Tian, Z.Z.; Wang, Q.; Shao, N.; Li, D.; He, Y.; Lei, W.W.; et al. Serological Survey of Zika Virus in Humans and Animals in Dejiang Prefecture, Guizhou Province, China. Biomed. Environ. Sci. 2019, 32, 875–880. [Google Scholar]
- Taylor, R.M. Studies on certain viruses isolated in the tropics of Africa and South America; their growth and behavior in the embryonated hen egg. J. Immunol. 1952, 68, 473–494. [Google Scholar] [CrossRef] [PubMed]
- Goodfellow, F.T.; Tesla, B.; Simchick, G.; Zhao, Q.; Hodge, T.; Brindley, M.A.; Stice, S.L. Zika Virus Induced Mortality and Microcephaly in Chicken Embryos. Stem Cells Dev. 2016, 25, 1691–1697. [Google Scholar] [CrossRef]
- Willard, K.A.; Demakovsky, L.; Tesla, B.; Goodfellow, F.T.; Stice, S.L.; Murdock, C.C.; Brindley, M.A. Zika Virus Exhibits Lineage-Specific Phenotypes in Cell Culture, in Aedes aegypti Mosquitoes, and in an Embryo Model. Viruses 2017, 9, 383. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, M.; Deng, J.; Yu, J.; Yang, X.; Zhao, W.; Chen, G.; Wang, P. Zika Virus Induced More Severe Inflammatory Response Than Dengue Virus in Chicken Embryonic Livers. Front. Microbiol. 2019, 10, 1127. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; Yip, C.C.; Tsang, J.O.; Tee, K.M.; Cai, J.P.; Chik, K.K.; Zhu, Z.; Chan, C.C.; Choi, G.K.; Sridhar, S.; et al. Differential cell line susceptibility to the emerging Zika virus: Implications for disease pathogenesis, non-vector-borne human transmission and animal reservoirs. Emerg. Microbes Infect. 2016, 5, e93. [Google Scholar] [CrossRef] [PubMed]
- Barr, K.L.; Anderson, B.D.; Prakoso, D.; Long, M.T. Working with Zika and Usutu Viruses In Vitro. PLoS Negl. Trop. Dis. 2016, 10, e0004931. [Google Scholar] [CrossRef] [PubMed]
- Lazear, H.M.; Govero, J.; Smith, A.M.; Platt, D.J.; Fernandez, E.; Miner, J.J.; Diamond, M.S. A Mouse Model of Zika Virus Pathogenesis. Cell Host Microbe 2016, 19, 720–730. [Google Scholar] [CrossRef] [PubMed]
- Dridi, M.; Rauw, F.; Muylkens, B.; Lecollinet, S.; van den Berg, T.; Lambrecht, B. Setting up a SPF chicken model for the pathotyping of West Nile virus (WNV) strains. Transbound. Emerg. Dis. 2013, 60 (Suppl. 2), 51–62. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Barber, M.R.; Aldridge, J.R., Jr.; Fleming-Canepa, X.; Wang, Y.D.; Webster, R.G.; Magor, K.E. Identification of avian RIG-I responsive genes during influenza infection. Mol. Immunol. 2013, 54, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, R.; Kuchipudi, S.V.; Chantrey, J.; Ganapathy, K. Pathogenicity and tissue tropism of infectious bronchitis virus is associated with elevated apoptosis and innate immune responses. Virology 2016, 488, 232–241. [Google Scholar] [CrossRef]
- Kuchipudi, S.V.; Tellabati, M.; Sebastian, S.; Londt, B.Z.; Jansen, C.; Vervelde, L.; Brookes, S.M.; Brown, I.H.; Dunham, S.P.; Chang, K.-C. Highly pathogenic avian influenza virus infection in chickens but not ducks is associated with elevated host immune and pro-inflammatory responses. Vet. Res. 2014, 45, 118. [Google Scholar] [CrossRef]
- Suzuki, K.; Okada, H.; Itoh, T.; Tada, T.; Mase, M.; Nakamura, K.; Kubo, M.; Tsukamoto, K. Association of increased pathogenicity of Asian H5N1 highly pathogenic avian influenza viruses in chickens with highly efficient viral replication accompanied by early destruction of innate immune responses. J. Virol. 2009, 83, 7475–7486. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Goebel, S.; Snyder, B.; Sellati, T.; Saeed, M.; Ptak, R.; Murray, M.; Bostwick, R.; Rayner, J.; Koide, F.; Kalkeri, R. A sensitive virus yield assay for evaluation of antivirals against Zika Virus. J. Virol. Methods 2016, 238, 13–20. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.L.; Meberg, B.; Schiltz, J.; Nilles, M.L.; Bradley, D.S. Zika Virus-Specific IgY Results Are Therapeutic Following a Lethal Zika Virus Challenge without Inducing Antibody-Dependent Enhancement. Viruses 2019, 11, 301. [Google Scholar] [CrossRef] [PubMed]
- Langevin, S.A.; Bunning, M.; Davis, B.; Komar, N. Experimental infection of chickens as candidate sentinels for West Nile virus. Emerg. Infect. Dis. 2001, 7, 726–729. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Chen, J.; Zhu, X.; An, S.; Dong, X.; Yu, J.; Zhang, S.; Wu, Y.; Li, G.; Zhang, Y.; et al. NLRP3 Inflammasome Activation Mediates Zika Virus-Associated Inflammation. J. Infect. Dis. 2018, 217, 1942–1951. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, G.; De, W.; Luo, Z.; Pan, P.; Tian, M.; Wang, Y.; Xiao, F.; Li, A.; Wu, K.; et al. Zika virus infection induces host inflammatory responses by facilitating NLRP3 inflammasome assembly and interleukin-1β secretion. Nat. Commun. 2018, 9, 106. [Google Scholar] [CrossRef] [PubMed]
- Orzalli, M.H.; Smith, A.; Jurado, K.A.; Iwasaki, A.; Garlick, J.A.; Kagan, J.C. An Antiviral Branch of the IL-1 Signaling Pathway Restricts Immune-Evasive Virus Replication. Mol. Cell 2018, 71, 825–840.e6. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.R.; Quicke, K.M.; Maddur, M.S.; O’Neal, J.T.; McDonald, C.E.; Fedorova, N.B.; Puri, V.; Shabman, R.S.; Pulendran, B.; Suthar, M.S. Zika Virus Antagonizes Type I Interferon Responses during Infection of Human Dendritic Cells. PLoS Pathog. 2017, 13, e1006164. [Google Scholar] [CrossRef]
- Grant, A.; Ponia, S.S.; Tripathi, S.; Balasubramaniam, V.; Miorin, L.; Sourisseau, M.; Schwarz, M.C.; Sánchez-Seco, M.P.; Evans, M.J.; Best, S.M.; et al. Zika Virus Targets Human STAT2 to Inhibit Type I Interferon Signaling. Cell Host Microbe 2016, 19, 882–890. [Google Scholar] [CrossRef]
- Tripathi, S.; Balasubramaniam, V.R.M.T.; Brown, J.A.; Mena, I.; Grant, A.; Bardina, S.V.; Maringer, K.; Schwarz, M.C.; Maestre, A.M.; Sourisseau, M.; et al. A novel Zika virus mouse model reveals strain specific differences in virus pathogenesis and host inflammatory immune responses. PLOS Pathog. 2017, 13, e1006258. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Thurmond, S.; Zhou, K.; Sánchez-Aparicio, M.T.; Fang, J.; Lu, J.; Gao, L.; Ren, W.; Cui, Y.; Veit, E.C.; et al. Structural basis for STAT2 suppression by flavivirus NS5. Nat. Struct. Mol. Biol. 2020, 27, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Parisien, J.-P.; Lenoir, J.J.; Alvarado, G.; Horvath, C.M. The Human STAT2 Coiled-Coil Domain Contains a Degron for Zika Virus Interferon Evasion. J. Virol. 2022, 96, e01301-21. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Alkie, T.N.; Yitbarek, A.; Hodgins, D.C.; Kulkarni, R.R.; Taha-Abdelaziz, K.; Sharif, S. Development of innate immunity in chicken embryos and newly hatched chicks: A disease control perspective. Avian Pathol. 2019, 48, 288–310. [Google Scholar] [CrossRef] [PubMed]
- Del Vesco, A.P.; Jang, H.J.; Monson, M.S.; Lamont, S.J. Role of the chicken oligoadenylate synthase-like gene during in vitro Newcastle disease virus infection. Poult. Sci. 2021, 100, 101067. [Google Scholar] [CrossRef] [PubMed]
- Tag-El-Din-Hassan, H.T.; Sasaki, N.; Moritoh, K.; Torigoe, D.; Maeda, A.; Agui, T. The chicken 2’-5’ oligoadenylate synthetase A inhibits the replication of West Nile virus. Jpn. J. Vet. Res. 2012, 60, 95–103. [Google Scholar] [PubMed]
- Tag-El-Din-Hassan, H.T.; Sasaki, N.; Torigoe, D.; Morimatsu, M.; Agui, T. Analysis of the Relationship Between Enzymatic and Antiviral Activities of the Chicken Oligoadenylate Synthetase-Like. J. Interferon Cytokine Res. 2017, 37, 71–80. [Google Scholar] [CrossRef]
- Saver, A.E.; Crawford, S.A.; Joyce, J.D.; Bertke, A.S. Route of Infection Influences Zika Virus Shedding in a Guinea Pig Model. Cells 2019, 8, 1437. [Google Scholar] [CrossRef]
- Talavera-Aguilar, L.G.; Murrieta, R.A.; Kiem, S.; Cetina-Trejo, R.C.; Baak-Baak, C.M.; Ebel, G.D.; Blitvich, B.J.; Machain-Williams, C. Infection, dissemination, and transmission efficiencies of Zika virus in Aedes aegypti after serial passage in mosquito or mammalian cell lines or alternating passage in both cell types. Parasites Vectors 2021, 14, 261. [Google Scholar] [CrossRef]
- Dudley, D.M.; Newman, C.M.; Lalli, J.; Stewart, L.M.; Koenig, M.R.; Weiler, A.M.; Semler, M.R.; Barry, G.L.; Zarbock, K.R.; Mohns, M.S.; et al. Infection via mosquito bite alters Zika virus tissue tropism and replication kinetics in rhesus macaques. Nat. Commun. 2017, 8, 2096. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Chick A | Chick B |
| crop a | 7.38 × 105 | 1.72 × 105 |
| liver | 5.20 × 104 | 3.16 × 104 |
| brain | 1.45 × 104 | - |
| kidney | 8.42 × 103 | - |
| spleen | 4.71 × 103 | - |
| lung | - b | - |
| heart | - | - |
| duodenum | - | - |
| eye | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nissly, R.H.; Lim, L.; Keller, M.R.; Bird, I.M.; Bhushan, G.; Misra, S.; Chothe, S.K.; Sill, M.C.; Kumar, N.V.; Sivakumar, A.V.N.; et al. The Susceptibility of Chickens to Zika Virus: A Comprehensive Study on Age-Dependent Infection Dynamics and Host Responses. Viruses 2024, 16, 569. https://doi.org/10.3390/v16040569
Nissly RH, Lim L, Keller MR, Bird IM, Bhushan G, Misra S, Chothe SK, Sill MC, Kumar NV, Sivakumar AVN, et al. The Susceptibility of Chickens to Zika Virus: A Comprehensive Study on Age-Dependent Infection Dynamics and Host Responses. Viruses. 2024; 16(4):569. https://doi.org/10.3390/v16040569
Chicago/Turabian StyleNissly, Ruth H., Levina Lim, Margo R. Keller, Ian M. Bird, Gitanjali Bhushan, Sougat Misra, Shubhada K. Chothe, Miranda C. Sill, Nagaram Vinod Kumar, A. V. N. Sivakumar, and et al. 2024. "The Susceptibility of Chickens to Zika Virus: A Comprehensive Study on Age-Dependent Infection Dynamics and Host Responses" Viruses 16, no. 4: 569. https://doi.org/10.3390/v16040569
APA StyleNissly, R. H., Lim, L., Keller, M. R., Bird, I. M., Bhushan, G., Misra, S., Chothe, S. K., Sill, M. C., Kumar, N. V., Sivakumar, A. V. N., Naik, B. R., Jayarao, B. M., & Kuchipudi, S. V. (2024). The Susceptibility of Chickens to Zika Virus: A Comprehensive Study on Age-Dependent Infection Dynamics and Host Responses. Viruses, 16(4), 569. https://doi.org/10.3390/v16040569