

Herpesvirus Infection of Endothelial Cells as a Systemic Pathological Axis in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome

Abstract
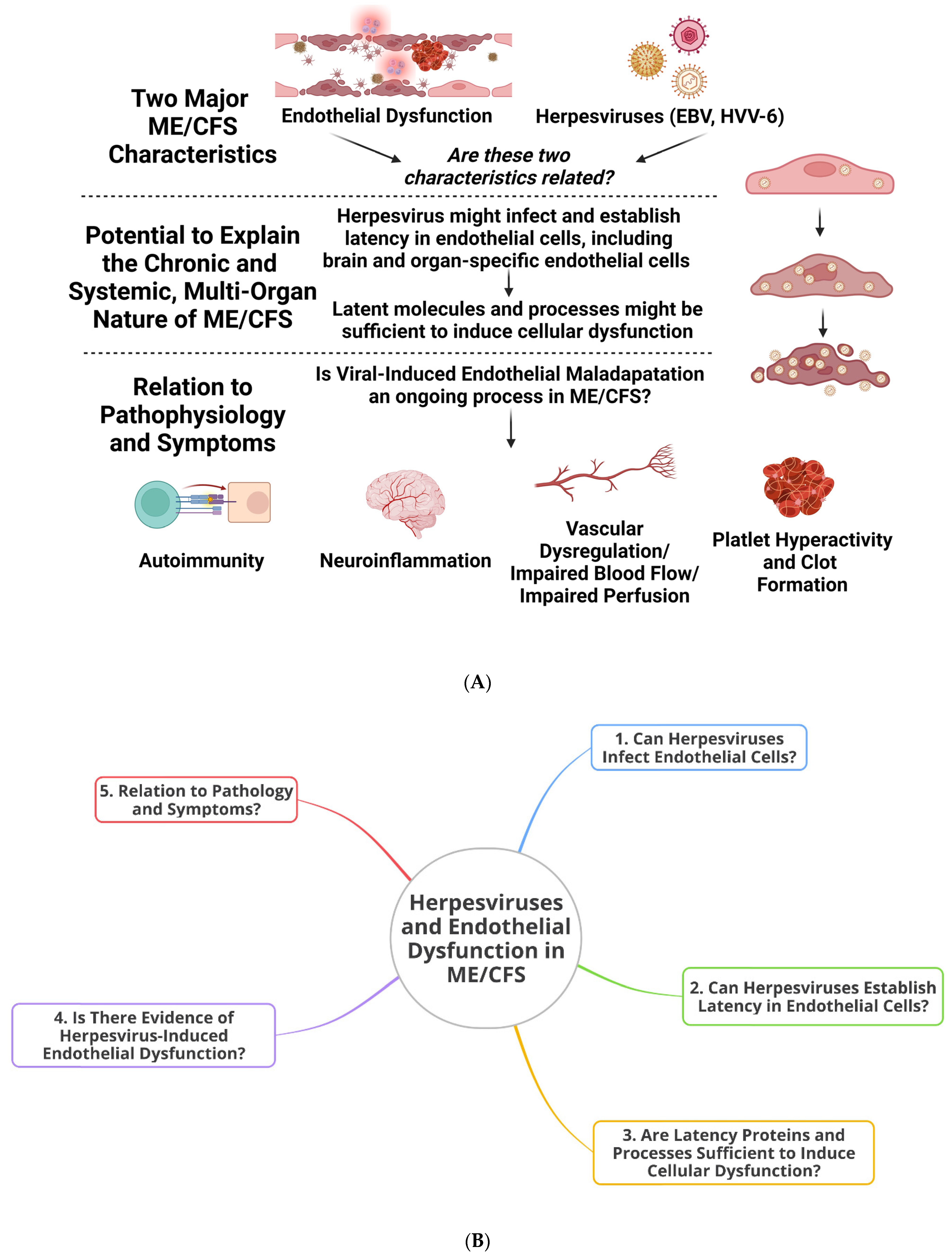
:1. Introduction
2. Infection of Endothelial Cells by Herpesviruses, Latent Modulation, Systemic Complications, and the Potential for Long-Term Maladaptation
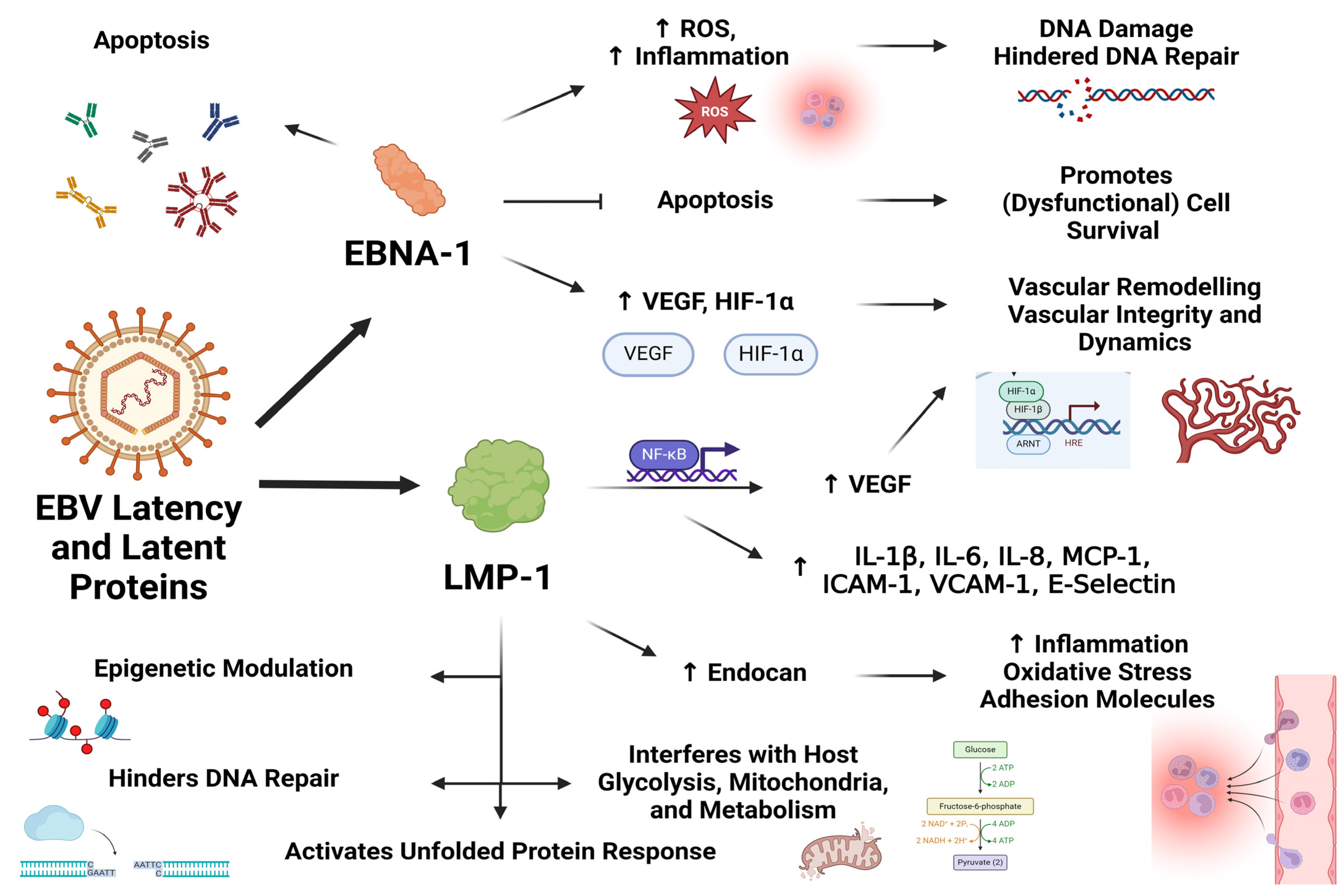
3. Latent Infection by Herpesviruses Is Sufficient to Bring about Cellular Dysfunction and Might Hold Relevance for Endothelial Dysfunction and Symptom Manifestation in ME/CFS
4. Evidence for Herpesvirus-Induced Endothelial Dysfunction
5. Herpesvirus-Induced Endothelial Dysfunction and Its Relevance to ME/CFS
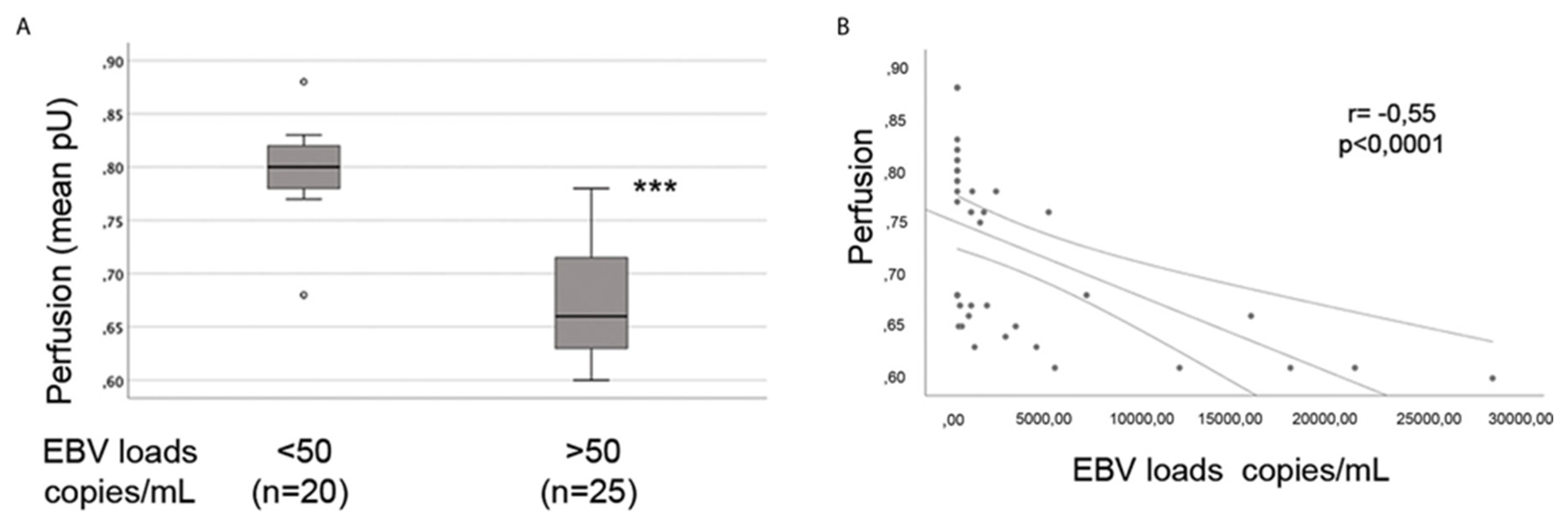
6. Endothelial Cells, Vascular Dysregulation, and Perfusion: Do Herpesviruses Have a Role to Play in the Dysregulation of Blood Flow Observed in ME/CFS?
7. Herpesviruses, Endothelial Cells, Platelets, and Coagulation
8. Herpesviruses and Neurological Issues in ME/CFS: Implications at the Cerebro-Endothelium?
9. Ways Forward
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lim, E.-J.; Son, C.-G. Review of case definitions for myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS). J. Transl. Med. 2020, 18, 289. [Google Scholar] [CrossRef] [PubMed]
- Rasa, S.; Nora-Krukle, Z.; Henning, N.; Eliassen, E.; Shikova, E.; Harrer, T.; Scheibenbogen, C.; Murovska, M.; Prusty, B.K. Chronic viral infections in myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS). J. Transl. Med. 2018, 16, 268. [Google Scholar] [CrossRef] [PubMed]
- Rasa-Dzelzkaleja, S.; Krumina, A.; Capenko, S.; Nora-Krukle, Z.; Gravelsina, S.; Vilmane, A.; Ievina, L.; Shoenfeld, Y.; Murovska, M. The persistent viral infections in the development and severity of myalgic encephalomyelitis/chronic fatigue syndrome. J. Transl. Med. 2023, 21, 33. [Google Scholar] [CrossRef] [PubMed]
- Ariza, M.E. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: The Human Herpesviruses Are Back! Biomolecules 2021, 11, 185. [Google Scholar] [CrossRef] [PubMed]
- Kasimir, F.; Toomey, D.; Liu, Z.; Kaiping, A.C.; Ariza, M.E.; Prusty, B.K. Tissue specific signature of HHV-6 infection in ME/CFS. Front. Mol. Biosci. 2022, 9, 1044964. [Google Scholar] [CrossRef] [PubMed]
- Shikova, E.; Reshkova, V.; Kumanova, A.; Raleva, S.; Alexandrova, D.; Capo, N.; Murovska, M. Cytomegalovirus, Epstein-Barr virus, and human herpesvirus-6 infections in patients with myalgic encephalomyelitis/chronic fatigue syndrome. J. Med. Virol. 2020, 92, 3682–3688. [Google Scholar] [CrossRef] [PubMed]
- Cox, B.S.; Alharshawi, K.; Mena-Palomo, I.; Lafuse, W.P.; Ariza, M.E. EBV/HHV-6A dUTPases contribute to myalgic encephalomyelitis/chronic fatigue syndrome pathophysiology by enhancing TFH cell differentiation and extrafollicular activities. JCI Insight 2022, 7, e158193. [Google Scholar] [CrossRef]
- Ruiz-Pablos, M.; Paiva, B.; Montero-Mateo, R.; Garcia, N.; Zabaleta, A. Epstein-Barr Virus and the Origin of Myalgic Encephalomyelitis or Chronic Fatigue Syndrome. Front. Immunol. 2021, 12, 656797. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.M.; Cox, B.; Lafuse, W.P.; Ariza, M.E. Epstein-Barr Virus dUTPase Induces Neuroinflammatory Mediators: Implications for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Clin. Ther. 2019, 41, 848–863. [Google Scholar] [CrossRef]
- Mozhgani, S.-H.; Rajabi, F.; Qurbani, M.; Erfani, Y.; Yaslianifard, S.; Moosavi, A.; Pourrostami, K.; Bagheri, A.B.; Soleimani, A.; Behzadian, F.; et al. Human Herpesvirus 6 Infection and Risk of Chronic Fatigue Syndrome: A Systematic Review and Meta-Analysis. Intervirology 2021, 65, 49–57. [Google Scholar] [CrossRef]
- Navaneetharaja, N.; Griffiths, V.; Wileman, T.; Carding, S.R. A Role for the Intestinal Microbiota and Virome in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS)? J. Clin. Med. 2016, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, W. ME/CFS, case definition, and serological response to Epstein-Barr virus. A systematic literature review. Fatigue Biomed. Health Behav. 2018, 6, 220–234. [Google Scholar] [CrossRef]
- Hatton, O.L.; Harris-Arnold, A.; Schaffert, S.; Krams, S.M.; Martinez, O.M. The interplay between Epstein-Barr virus and B lymphocytes: Implications for infection, immunity, and disease. Immunol. Res. 2014, 58, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Sinzger, C.; Grefte, A.; Plachter, B.; Gouw, A.S.; The, T.H.; Jahn, G. Fibroblasts, epithelial cells, endothelial cells and smooth muscle cells are major targets of human cytomegalovirus infection in lung and gastrointestinal tissues. J. Gen. Virol. 1995, 76, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Adler, B.; Sinzger, C. Endothelial cells in human cytomegalovirus infection: One host cell out of many or a crucial target for virus spread? Thromb. Haemost. 2009, 102, 1057–1063. [Google Scholar] [PubMed]
- Scherbakov, N.; Szklarski, M.; Hartwig, J.; Sotzny, F.; Lorenz, S.; Meyer, A.; Grabowski, P.; Doehner, W.; Scheibenbogen, C. Peripheral endothelial dysfunction in myalgic encephalomyelitis/chronic fatigue syndrome. ESC Heart Fail. 2020, 7, 1064–1071. [Google Scholar] [CrossRef] [PubMed]
- Sørland, K.; Sandvik, M.K.; Rekeland, I.G.; Ribu, L.; Småstuen, M.C.; Mella, O.; Fluge, Ø. Reduced Endothelial Function in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome–Results from Open-Label Cyclophosphamide Intervention Study. Front. Med. 2021, 8, 642710. [Google Scholar] [CrossRef] [PubMed]
- Sandvik, M.K.; Sørland, K.; Leirgul, E.; Rekeland, I.G.; Stavland, C.S.; Mella, O.; Fluge, Ø. Endothelial dysfunction in ME/CFS patients. PLoS ONE 2023, 18, e0280942. [Google Scholar] [CrossRef]
- Haffke, M.; Freitag, H.; Rudolf, G.; Seifert, M.; Doehner, W.; Scherbakov, N.; Hanitsch, L.; Wittke, K.; Bauer, S.; Konietschke, F.; et al. Endothelial dysfunction and altered endothelial biomarkers in patients with post-COVID-19 syndrome and chronic fatigue syndrome (ME/CFS). J. Transl. Med. 2022, 20, 138. [Google Scholar] [CrossRef]
- Blauensteiner, J.; Bertinat, R.; León, L.E.; Riederer, M.; Sepúlveda, N.; Westermeier, F. Altered endothelial dysfunction-related miRs in plasma from ME/CFS patients. Sci. Rep. 2021, 11, 10604. [Google Scholar] [CrossRef]
- Bertinat, R.; Villalobos-Labra, R.; Hofmann, L.; Blauensteiner, J.; Sepúlveda, N.; Westermeier, F. Decreased NO production in endothelial cells exposed to plasma from ME/CFS patients. Vasc. Pharmacol. 2022, 143, 106953. [Google Scholar] [CrossRef] [PubMed]
- Newton, D.J.; Kennedy, G.; Chan, K.K.; Lang, C.C.; Belch, J.J.; Khan, F. Large and small artery endothelial dysfunction in chronic fatigue syndrome. Int. J. Cardiol. 2012, 154, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Flaskamp, L.; Roubal, C.; Uddin, S.; Sotzny, F.; Kedor, C.; Bauer, S.; Scheibenbogen, C.; Seifert, M. Serum of Post-COVID-19 Syndrome Patients with or without ME/CFS Differentially Affects Endothelial Cell Function In Vitro. Cells 2022, 11, 2376. [Google Scholar] [CrossRef] [PubMed]
- Cambras, T.; Zerón-Rugerio, M.F.; Díez-Noguera, A.; Zaragozá, M.C.; Domingo, J.C.; Sanmartin-Sentañes, R.; Alegre-Martin, J.; Castro-Marrero, J. Skin Temperature Circadian Rhythms and Dysautonomia in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: The Role of Endothelin-1 in the Vascular Tone Dysregulation. Int. J. Mol. Sci. 2023, 24, 4835. [Google Scholar] [CrossRef] [PubMed]
- Campen, C.M.C.v.; Rowe, P.C.; Visser, F.C. Orthostatic Symptoms and Reductions in Cerebral Blood Flow in Long-Haul COVID-19 Patients: Similarities with Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Medicina 2022, 58, 28. [Google Scholar] [CrossRef] [PubMed]
- Van Campen, C.M.C.; Rowe, P.C.; Visser, F.C. Cerebral blood flow remains reduced after tilt testing in myalgic encephalomyelitis/chronic fatigue syndrome patients. Clin. Neurophysiol. Pract. 2021, 6, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Van Campen, C.M.C.; Rowe, P.C.; Visser, F.C. Cerebral Blood Flow Is Reduced in Severe Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Patients During Mild Orthostatic Stress Testing: An Exploratory Study at 20 Degrees of Head-Up Tilt Testing. Healthcare 2020, 8, 169. [Google Scholar] [CrossRef] [PubMed]
- Van Campen, C.M.C.; Verheugt, F.W.A.; Rowe, P.C.; Visser, F.C. Cerebral blood flow is reduced in ME/CFS during head-up tilt testing even in the absence of hypotension or tachycardia: A quantitative, controlled study using Doppler echography. Clin. Neurophysiol. Pract. 2020, 5, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Staud, R.; Boissoneault, J.; Craggs, J.G.; Lai, S.; Robinson, M.E. Task related cerebral blood flow changes of patients with chronic fatigue syndrome: An arterial spin labeling study. Fatigue Biomed. Health Behav. 2018, 6, 63–79. [Google Scholar] [CrossRef]
- Boissoneault, J.; Letzen, J.; Robinson, M.; Staud, R. Cerebral blood flow and heart rate variability predict fatigue severity in patients with chronic fatigue syndrome. Brain Imaging Behav. 2019, 13, 789–797. [Google Scholar] [CrossRef]
- Biswal, B.; Kunwar, P.; Natelson, B.H. Cerebral blood flow is reduced in chronic fatigue syndrome as assessed by arterial spin labeling. J. Neurol. Sci. 2011, 301, 9–11. [Google Scholar] [CrossRef] [PubMed]
- COSTA, D.C.; Tannock, C.; Brostoff, J. Brainstem perfusion is impaired in chronic fatigue syndrome. QJM Int. J. Med. 1995, 88, 767–773. [Google Scholar]
- Yoshiuchi, K.; Farkas, J.; Natelson, B.H. Patients with chronic fatigue syndrome have reduced absolute cortical blood flow. Clin. Physiol. Funct. Imaging 2006, 26, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Shungu, D.C.; Weiduschat, N.; Murrough, J.W.; Mao, X.; Pillemer, S.; Dyke, J.P.; Medow, M.S.; Natelson, B.H.; Stewart, J.M.; Mathew, S.J. Increased ventricular lactate in chronic fatigue syndrome. III. Relationships to cortical glutathione and clinical symptoms implicate oxidative stress in disorder pathophysiology. NMR Biomed. 2012, 25, 1073–1087. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Julin, P.; Li, T.-Q. Limbic Perfusion Is Reduced in Patients with Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Tomography 2021, 7, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Ambrosino, P.; Calcaterra, I.; Molino, A.; Moretta, P.; Lupoli, R.; Spedicato, G.A.; Papa, A.; Motta, A.; Maniscalco, M.; Di Minno, M.N.D. Persistent Endothelial Dysfunction in Post-Acute COVID-19 Syndrome: A Case-Control Study. Biomedicines 2021, 9, 957. [Google Scholar] [CrossRef] [PubMed]
- Stahl, K.; Gronski, P.A.; Kiyan, Y.; Seeliger, B.; Bertram, A.; Pape, T.; Welte, T.; Hoeper, M.M.; Haller, H.; David, S. Injury to the Endothelial Glycocalyx in Critically Ill Patients with COVID-19. Am. J. Respir. Crit. Care Med. 2020, 202, 1178–1181. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Khan, M.A.; Putrino, D.; Woodcock, A.; Kell, D.B.; Pretorius, E. Long COVID: Pathophysiological factors and abnormalities of coagulation. Trends Endocrinol. Metab. 2023, 34, 321–344. [Google Scholar] [CrossRef] [PubMed]
- Kedor, C.; Freitag, H.; Meyer-Arndt, L.; Wittke, K.; Zoller, T.; Steinbeis, F.; Haffke, M.; Rudolf, G.; Heidecker, B.; Volk, H.D.; et al. Chronic COVID-19 Syndrome and Chronic Fatigue Syndrome (ME/CFS) following the first pandemic wave in Germany—A first analysis of a prospective observational study. medRxiv, 2021; medRxiv:06.21249256. [Google Scholar]
- Poenaru, S.; Abdallah, S.J.; Corrales-Medina, V.; Cowan, J. COVID-19 and post-infectious myalgic encephalomyelitis/chronic fatigue syndrome: A narrative review. Ther. Adv. Infect. Dis. 2021, 8, 20499361211009385. [Google Scholar] [CrossRef]
- Wong, T.L.; Weitzer, D.J. Long COVID and Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS)—A Systemic Review and Comparison of Clinical Presentation and Symptomatology. Medicina 2021, 57, 418. [Google Scholar] [CrossRef]
- Tate, W.; Walker, M.; Sweetman, E.; Helliwell, A.; Peppercorn, K.; Edgar, C.; Blair, A.; Chatterjee, A. Molecular Mechanisms of Neuroinflammation in ME/CFS and Long COVID to Sustain Disease and Promote Relapses. Front. Neurol. 2022, 13, 877772. [Google Scholar] [CrossRef] [PubMed]
- Nunes, J.M.; Kruger, A.; Proal, A.; Kell, D.B.; Pretorius, E. The Occurrence of Hyperactivated Platelets and Fibrinaloid Microclots in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Pharmaceuticals 2022, 15, 931. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Vlok, M.; Venter, C.; Bezuidenhout, J.A.; Laubscher, G.J.; Steenkamp, J.; Kell, D.B. Persistent clotting protein pathology in Long COVID/Post-Acute Sequelae of COVID-19 (PASC) is accompanied by increased levels of antiplasmin. Cardiovasc. Diabetol. 2021, 20, 172. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.B.; Pretorius, E. The potential role of ischaemia–reperfusion injury in chronic, relapsing diseases such as rheumatoid arthritis, Long COVID, and ME/CFS: Evidence, mechanisms, and therapeutic implications. Biochem. J. 2022, 479, 1653–1708. [Google Scholar] [CrossRef] [PubMed]
- Laubshder, G.; Khan, M.; Venter, C.; Pretorius, J.; Kell, D.; Pretorius, E. Treatment of Long COVID symptoms with triple anticoagulant therapy. Res. Sq. 2023; Preprint. [Google Scholar] [CrossRef]
- Bonilla, H.; Hampton, D.; Marques de Menezes, E.G.; Deng, X.; Montoya, J.G.; Anderson, J.; Norris, P.J. Comparative Analysis of Extracellular Vesicles in Patients with Severe and Mild Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Front. Immunol. 2022, 13, 841910. [Google Scholar] [CrossRef] [PubMed]
- Jahanbani, F.; Maynard, R.D.; Sing, J.C.; Jahanbani, S.; Perrino, J.J.; Spacek, D.V.; Davis, R.W.; Snyder, M.P. Phenotypic characteristics of peripheral immune cells of Myalgic encephalomyelitis/chronic fatigue syndrome via transmission electron microscopy: A pilot study. PLoS ONE 2022, 17, e0272703. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Vu, L.T.; Zhu, H.; Iu, D.S.H.; Fogarty, E.A.; Kwak, Y.; Chen, W.; Franconi, C.J.; Munn, P.R.; Tate, A.E.; et al. Single-cell transcriptomics of the immune system in ME/CFS at baseline and following symptom provocation. bioRxiv 2022. bioRxiv:13.512091. [Google Scholar]
- Berg, D.; Berg, L.H.; Couvaras, J.; Harrison, H. Chronic fatigue syndrome and/or fibromyalgia as a variation of antiphospholipid antibody syndrome: An explanatory model and approach to laboratory diagnosis. Blood Coagul. Fibrinolysis 1999, 10, 435–438. [Google Scholar] [CrossRef]
- Van Hinsbergh, V.W.M. Endothelium—Role in regulation of coagulation and inflammation. Semin. Immunopathol. 2012, 34, 93–106. [Google Scholar] [CrossRef]
- Neubauer, K.; Zieger, B. Endothelial cells and coagulation. Cell Tissue Res. 2022, 387, 391–398. [Google Scholar] [CrossRef]
- Wirth, K.; Scheibenbogen, C. A Unifying Hypothesis of the Pathophysiology of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): Recognitions from the finding of autoantibodies against ß2-adrenergic receptors. Autoimmun. Rev. 2020, 19, 102527. [Google Scholar] [CrossRef]
- Wirth, K.J.; Scheibenbogen, C. Pathophysiology of skeletal muscle disturbances in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). J. Transl. Med. 2021, 19, 162. [Google Scholar] [CrossRef] [PubMed]
- Wirth, K.J.; Scheibenbogen, C.; Paul, F. An attempt to explain the neurological symptoms of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. J. Transl. Med. 2021, 19, 471. [Google Scholar] [CrossRef] [PubMed]
- Sfera, A.; Osorio, C.; Zapata Martín del Campo, C.; Pereida, S.; Maurer, S.; Maldonado, J.C.; Kozlakidis, Z. Endothelial Senescence and Chronic Fatigue Syndrome, a COVID-19 Based Hypothesis. Front. Cell. Neurosci. 2021, 15, 673217. [Google Scholar] [CrossRef] [PubMed]
- Chapenko, S.; Krumina, A.; Logina, I.; Rasa, S.; Chistjakovs, M.; Sultanova, A.; Viksna, L.; Murovska, M. Association of Active Human Herpesvirus-6, -7 and Parvovirus B19 Infection with Clinical Outcomes in Patients with Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Adv. Virol. 2012, 2012, 205085. [Google Scholar] [CrossRef] [PubMed]
- Blomberg, J.; Rizwan, M.; Böhlin-Wiener, A.; Elfaitouri, A.; Julin, P.; Zachrisson, O.; Rosén, A.; Gottfries, C.-G. Antibodies to Human Herpesviruses in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Patients. Front. Immunol. 2019, 10, 1946. [Google Scholar] [CrossRef] [PubMed]
- Ban, S.; Goto, Y.; Kamada, K.; Takahama, M.; Watanabe, H.; Iwahori, T.; Takeuchi, H. Systemic granulomatous arteritis associated with Epstein-Barr virus infection. Virchows Arch. 1999, 434, 249–254. [Google Scholar] [CrossRef]
- Guarner, J.; Unger, E.R. Association of Epstein-Barr virus in epithelioid angiomatosis of AIDS patients. Am. J. Surg. Pathol. 1990, 14, 956–960. [Google Scholar] [CrossRef]
- Farina, A.; Cirone, M.; York, M.; Lenna, S.; Padilla, C.; Mclaughlin, S.; Faggioni, A.; Lafyatis, R.; Trojanowska, M.; Farina, G.A. Epstein–Barr Virus Infection Induces Aberrant TLR Activation Pathway and Fibroblast–Myofibroblast Conversion in Scleroderma. J. Investig. Dermatol. 2014, 134, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Kanai, K.; Kuwabara, S.; Mori, M.; Arai, K.; Yamamoto, T.; Hattori, T. Leukocytoclastic-vasculitic neuropathy associated with chronic Epstein–Barr virus infection. Muscle Nerve 2003, 27, 113–116. [Google Scholar] [CrossRef]
- Truszewska, A.; Wirkowska, A.; Gala, K.; Truszewski, P.; Krzemień-Ojak, L.; Mucha, K.; Pączek, L.; Foroncewicz, B. EBV load is associated with cfDNA fragmentation and renal damage in SLE patients. Lupus 2021, 30, 1214–1225. [Google Scholar] [CrossRef]
- Jones, K.; Rivera, C.; Sgadari, C.; Franklin, J.; E Max, E.; Bhatia, K.; Tosato, G. Infection of human endothelial cells with Epstein-Barr virus. J. Exp. Med. 1995, 182, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Casiraghi, C.; Dorovini-Zis, K.; Horwitz, M.S. Epstein-Barr virus infection of human brain microvessel endothelial cells: A novel role in multiple sclerosis. J. Neuroimmunol. 2011, 230, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Farina, A.; Rosato, E.; York, M.; Gewurz, B.E.; Trojanowska, M.; Farina, G.A. Innate Immune Modulation Induced by EBV Lytic Infection Promotes Endothelial Cell Inflammation and Vascular Injury in Scleroderma. Front. Immunol. 2021, 12, 651013. [Google Scholar] [CrossRef] [PubMed]
- Caruso, A.; Rotola, A.; Comar, M.; Favilli, F.; Galvan, M.; Tosetti, M.; Campello, C.; Caselli, E.; Alessandri, G.; Grassi, M.; et al. HHV-6 infects human aortic and heart microvascular endothelial cells, increasing their ability to secrete proinflammatory chemokines. J. Med. Virol. 2002, 67, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, R.; D’accolti, M.; Bortolotti, D.; Caccuri, F.; Caruso, A.; Di Luca, D.; Caselli, E. Human Herpesvirus 6A and 6B inhibit in vitro angiogenesis by induction of Human Leukocyte Antigen G. Sci. Rep. 2018, 8, 17683. [Google Scholar] [CrossRef] [PubMed]
- Caruso, A.; Favilli, F.; Rotola, A.; Comar, M.; Horejsh, D.; Alessandri, G.; Grassi, M.; Di Luca, D.; Fiorentini, S. Human herpesvirus-6 modulates RANTES production in primary human endothelial cell cultures. J. Med. Virol. 2003, 70, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Rotola, A.; Di Luca, D.; Cassai, E.; Ricotta, D.; Giulio, A.; Turano, A.; Caruso, A.; Muneretto, C. Human herpesvirus 6 infects and replicates in aortic endothelium. J. Clin. Microbiol. 2000, 38, 3135–3136. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.A.; Shanley, J.D. Chronic infection of human umbilical vein endothelial cells by human herpesvirus-6. J. Gen. Virol. 1998, 79, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Eliassen, E.; Krueger, G. Human herpesvirus 6-induced inflammatory cardiomyopathy in immunocompetent children. Ann. Pediatr. Cardiol. 2017, 10, 259–268. [Google Scholar]
- Visser, M.R.; Tracy, P.B.; Vercellotti, G.M.; Goodman, J.L.; White, J.G.; Jacob, H.S. Enhanced thrombin generation and platelet binding on herpes simplex virus-infected endothelium. Proc. Natl. Acad. Sci. USA 1988, 85, 8227–8230. [Google Scholar] [CrossRef]
- Fonsato, V.; Buttiglieri, S.; Deregibus, M.C.; Bussolati, B.; Caselli, E.; Di Luca, D.; Camussi, G. PAX2 expression by HHV-8–infected endothelial cells induced a proangiogenic and proinvasive phenotype. Blood 2008, 111, 2806–2815. [Google Scholar] [CrossRef] [PubMed]
- Dolcino, M.; Puccetti, A.; Barbieri, A.; Bason, C.; Tinazzi, E.; Ottria, A.; Patuzzo, G.; Martinelli, N.; Lunardi, C. Infections and autoimmunity: Role of human cytomegalovirus in autoimmune endothelial cell damage. Lupus 2015, 24, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Lunardi, C.; Bason, C.; Corrocher, R.; Puccetti, A. Induction of endothelial cell damage by hCMV molecular mimicry. Trends Immunol. 2005, 26, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.; Xu, L.; Chen, D.; Tang, W.; Huang, Y. Human cytomegalovirus-encoded miR-UL112 contributes to HCMV-mediated vascular diseases by inducing vascular endothelial cell dysfunction. Virus Genes. 2018, 54, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Yaiw, K.-C.; Mohammad, A.-A.; Costa, H.; Taher, C.; Badrnya, S.; Assinger, A.; Wilhelmi, V.; Ananthaseshan, S.; Estekizadeh, A.; Davoudi, B.; et al. Human Cytomegalovirus Up-Regulates Endothelin Receptor Type B: Implication for Vasculopathies? Open Forum Infect. Dis. 2015, 2, ofv155. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.I. Herpesvirus Latency. J. Clin. Investig. 2020, 130, 3361–3369. [Google Scholar] [CrossRef] [PubMed]
- Weidner-Glunde, M.; Kruminis-Kaszkiel, E.; Savanagouder, M. Herpesviral Latency—Common Themes. Pathogens 2020, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Caruso, A.; Caselli, E.; Fiorentini, S.; Rotola, A.; Prandini, A.; Garrafa, E.; Saba, E.; Alessandri, G.; Cassai, E.; Di Luca, D. U94 of human herpesvirus 6 inhibits in vitro angiogenesis and lymphangiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 20446–20451. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, D.; Luka, J.; Kallakury, B.; Rusnock, E.; Azumi, N. Immortalization and transformation of human endothelial cells infected with human herpesvirus-6 Objective Background. In Laboratory Investigation; Nature Publishing Group: New York, NY, USA, 2023. [Google Scholar]
- Brewer, J.H.; Berg, D. Hypercoaguable State Associated with Active Human Herpesvirus-6 (HHV-6) Viremia in Patients with Chronic Fatigue Syndrome. J. Chronic Fatigue Syndr. 2001, 8, 111–116. [Google Scholar] [CrossRef]
- Liu, Z.; Hollmann, C.; Kalanidhi, S.; Grothey, A.; Keating, S.; Mena-Palomo, I.; Lamer, S.; Schlosser, A.; Kaiping, K.; Scheller, C.; et al. Increased circulating fibronectin, depletion of natural IgM and heightened EBV, HSV-1 reactivation in ME/CFS and long COVID. medRxiv, 2023; medRxiv:2023.06.23.23291827. [Google Scholar]
- Kaprelyants, A.S.; Gottschal, J.C.; Kell, D.B. Dormancy in non-sporulating bacteria. FEMS Microbiol. Rev. 1993, 10, 271–285. [Google Scholar] [CrossRef]
- Kell, D.B.; Kaprelyants, A.S.; Weichart, D.H.; Harwood, C.R.; Barer, M.R. Viability and activity in readily culturable bacteria: A review and discussion of the practical issues. Antonie van Leeuwenhoek 1998, 73, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Nikolich-Zugich, J.; Goodrum, F.; Knox, K.; Smithey, M.J. Known unknowns: How might the persistent herpesvirome shape immunity and aging? Curr. Opin. Immunol. 2017, 48, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Simanek, A.M.; Dowd, J.B.; Pawelec, G.; Melzer, D.; Dutta, A.; Aiello, A.E. Seropositivity to cytomegalovirus, inflammation, all-cause and cardiovascular disease-related mortality in the United States. PLoS ONE 2011, 6, e16103. [Google Scholar] [CrossRef] [PubMed]
- Griffin, B.D.; Verweij, M.C.; Wiertz, E.J.H.J. Herpesviruses and immunity: The art of evasion. Vet. Microbiol. 2010, 143, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, O.; Zdunek, S.; Felker, A.; Salehpour, M.; Alkass, K.; Bernard, S.; Sjostrom, S.L.; Szewczykowska, M.; Jackowska, T.; dos Remedios, C.; et al. Dynamics of Cell Generation and Turnover in the Human Heart. Cell 2015, 161, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Hobson, B.; Denekamp, J. Endothelial proliferation in tumours and normal tissues: Continuous labelling studies. Br. J. Cancer 1984, 49, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Xiong, A.; Clarke-Katzenberg, R.H.; Valenzuela, G.; Izumi, K.M.; Millan, M.T. Epstein-Barr virus latent membrane protein 1 activates nuclear factor-kappa B in human endothelial cells and inhibits apoptosis. Transplantation 2004, 78, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, M.; Mori, Y.; Ueda, K.; Kondo, K.; Yamanishi, K. Productive human herpesvirus 6 infection causes aberrant accumulation of p53 and prevents apoptosis. J. Gen. Virol. 2004, 85, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G.; Akbar, A.; Beverley, P.; Caruso, C.; Derhovanessian, E.; Fülöp, T.; Griffiths, P.; Grubeck-Loebenstein, B.; Hamprecht, K.; Jahn, G.; et al. Immunosenescence and Cytomegalovirus: Where do we stand after a decade? Immun. Ageing 2010, 7, 13. [Google Scholar] [CrossRef]
- Hogestyn, J.M.; Mock, D.J.; Mayer-Proschel, M. Contributions of neurotropic human herpesviruses herpes simplex virus 1 and human herpesvirus 6 to neurodegenerative disease pathology. Neural Regen. Res. 2018, 13, 211–221. [Google Scholar]
- Yin, H.; Qu, J.; Peng, Q.; Gan, R. Molecular mechanisms of EBV-driven cell cycle progression and oncogenesis. Med. Microbiol. Immunol. 2019, 208, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Elgui de Oliveira, D.; Müller-Coan, B.G.; Pagano, J.S. Viral Carcinogenesis Beyond Malignant Transformation: EBV in the Progression of Human Cancers. Trends Microbiol. 2016, 24, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Yajima, M.; Ikuta, K. Epstein-Barr virus strain variation and cancer. Cancer Sci. 2019, 110, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Niller, H.H.; Wolf, H.; Ay, E.; Minarovits, J. Epigenetic dysregulation of epstein-barr virus latency and development of autoimmune disease. Adv. Exp. Med. Biol. 2011, 711, 82–102. [Google Scholar] [PubMed]
- Lee, M.-A.; Diamond Margaret, E.; Yates John, L. Genetic Evidence that EBNA-1 Is Needed for Efficient, Stable Latent Infection by Epstein-Barr Virus. J. Virol. 1999, 73, 2974–2982. [Google Scholar] [CrossRef] [PubMed]
- Leight, E.R.; Sugden, B. EBNA-1: A protein pivotal to latent infection by Epstein–Barr virus. Rev. Med. Virol. 2000, 10, 83–100. [Google Scholar] [CrossRef]
- Szymula, A.; Palermo, R.D.; Bayoumy, A.; Groves, I.J.; Ba Abdullah, M.; Holder, B.; White, R.E. Epstein-Barr virus nuclear antigen EBNA-LP is essential for transforming naïve B cells, and facilitates recruitment of transcription factors to the viral genome. PLoS Pathog. 2018, 14, e1006890. [Google Scholar] [CrossRef]
- Young, L.S.; Murray, P.G. Epstein–Barr virus and oncogenesis: From latent genes to tumours. Oncogene 2003, 22, 5108–5121. [Google Scholar] [CrossRef]
- Kang, M.-S.; Kieff, E. Epstein–Barr virus latent genes. Exp. Mol. Med. 2015, 47, e131. [Google Scholar] [CrossRef]
- Gruhne, B.; Sompallae, R.; Marescotti, D.; Kamranvar, S.A.; Gastaldello, S.; Masucci, M.G. The Epstein-Barr virus nuclear antigen-1 promotes genomic instability via induction of reactive oxygen species. Proc. Natl. Acad. Sci. USA 2009, 106, 2313–2318. [Google Scholar] [CrossRef]
- Kamranvar, S.A.; Masucci, M.G. The Epstein–Barr virus nuclear antigen-1 promotes telomere dysfunction via induction of oxidative stress. Leukemia 2011, 25, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Gruhne, B.; Sompallae, R.; Masucci, M.G. Three Epstein–Barr virus latency proteins independently promote genomic instability by inducing DNA damage, inhibiting DNA repair and inactivating cell cycle checkpoints. Oncogene 2009, 28, 3997–4008. [Google Scholar] [CrossRef] [PubMed]
- Frappier, L. Contributions of Epstein-Barr nuclear antigen 1 (EBNA1) to cell immortalization and survival. Viruses 2012, 4, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Poole, B.D.; Scofield, R.H.; Harley, J.B.; James, J.A. Epstein-Barr virus and molecular mimicry in systemic lupus erythematosus. Autoimmunity 2006, 39, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Lanz, T.V.; Brewer, R.C.; Ho, P.P.; Moon, J.-S.; Jude, K.M.; Fernandez, D.; Fernandes, R.A.; Gomez, A.M.; Nadj, G.-S.; Bartley, C.M.; et al. Clonally expanded B cells in multiple sclerosis bind EBV EBNA1 and GlialCAM. Nature 2022, 603, 321–327. [Google Scholar] [CrossRef] [PubMed]
- De Bellis, A.; Bellastella, G.; Pernice, V.; Cirillo, P.; Longo, M.; Maio, A.; Scappaticcio, L.; Maiorino, M.I.; Bellastella, A.; Esposito, K.; et al. Hypothalamic-Pituitary Autoimmunity and Related Impairment of Hormone Secretions in Chronic Fatigue Syndrome. J. Clin. Endocrinol. Metab. 2021, 106, e5147–e5155. [Google Scholar] [CrossRef] [PubMed]
- Sotzny, F.; Blanco, J.; Capelli, E.; Castro-Marrero, J.; Steiner, S.; Murovska, M.; Scheibenbogen, C. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome—Evidence for an autoimmune disease. Autoimmun. Rev. 2018, 17, 601–609. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.D.; Owen, T.J.; Wood, V.H.J.; Date, K.L.; Valentine, R.; Chukwuma, M.B.; Arrand, J.R.; Dawson, C.W.; Young, L.S. Epstein-Barr virus-encoded EBNA1 modulates the AP-1 transcription factor pathway in nasopharyngeal carcinoma cells and enhances angiogenesis in vitro. J. Gen. Virol. 2008, 89 Pt 11, 2833–2842. [Google Scholar] [CrossRef] [PubMed]
- Murono, S.; Inoue, H.; Tanabe, T.; Joab, I.; Yoshizaki, T.; Furukawa, M.; Pagano, J.S. Induction of cyclooxygenase-2 by Epstein–Barr virus latent membrane protein 1 is involved in vascular endothelial growth factor production in nasopharyngeal carcinoma cells. Proc. Natl. Acad. Sci. USA 2001, 98, 6905–6910. [Google Scholar] [CrossRef]
- Kung, C.-P.; Raab-Traub, N. Epstein-Barr Virus Latent Membrane Protein 1 Induces Expression of the Epidermal Growth Factor Receptor through Effects on Bcl-3 and STAT3. J. Virol. 2008, 82, 5486–5493. [Google Scholar] [CrossRef]
- Gires, O.; Kohlhuber, F.; Kilger, E.; Baumann, M.; Kieser, A.; Kaiser, C.; Zeidler, R.; Scheffer, B.; Ueffing, M.; Hammerschmidt, W. Latent membrane protein 1 of Epstein-Barr virus interacts with JAK3 and activates STAT proteins. Embo J. 1999, 18, 3064–3073. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, A.G.; Young, L.S. LMP1 structure and signal transduction. Semin. Cancer Biol. 2001, 11, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.-H.; Chou, S.-F.; Chen, C.-L.; Hung, H.; Lai, C.-Y.; Yang, P.-M.; Jeng, Y.-M.; Liaw, S.-F.; Kuo, H.-H.; Hsu, H.-C.; et al. Upregulation of endocan by Epstein-Barr virus latent membrane protein 1 and its clinical significance in nasopharyngeal carcinoma. PLoS ONE 2013, 8, e82254. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, L.; Yu, X.-H.; Hu, M.; Zhang, Y.-K.; Liu, X.; He, P.; Ouyang, X. Endocan: A Key Player of Cardiovascular Disease. Front. Cardiovasc. Med. 2021, 8, 798699. [Google Scholar] [CrossRef] [PubMed]
- Bentz, G.L.; Whitehurst, C.B.; Pagano, J.S. Epstein-Barr virus latent membrane protein 1 (LMP1) C-terminal-activating region 3 contributes to LMP1-mediated cellular migration via its interaction with Ubc9. J. Virol. 2011, 85, 10144–10153. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Hu, Z.-Y.; Dong, X.; Tan, Z.; Li, W.; Tang, M.; Chen, L.; Yang, L.; Tao, Y.; Jiang, Y.; et al. Targeting Epstein-Barr virus oncoprotein LMP1-mediated glycolysis sensitizes nasopharyngeal carcinoma to radiation therapy. Oncogene 2014, 33, 4568–4578. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.K.; Dawson, C.W.; Young, L.S.; Ko, C.; Hau, P.; Lo, K. Activation of the FGFR1 signalling pathway by the Epstein-Barr virus-encoded LMP1 promotes aerobic glycolysis and transformation of human nasopharyngeal epithelial cells. J. Pathol. 2015, 237, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yan, B.; Lai, W.; Shi, Y.; Xiao, D.; Jia, J.; Liu, S.; Li, H.; Lu, J.; Li, Z.; et al. Repression of Hox genes by LMP1 in nasopharyngeal carcinoma and modulation of glycolytic pathway genes by HoxC8. Oncogene 2015, 34, 6079–6091. [Google Scholar] [CrossRef] [PubMed]
- Hino, R.; Uozaki, H.; Murakami, N.; Ushiku, T.; Shinozaki, A.; Ishikawa, S.; Morikawa, T.; Nakaya, T.; Sakatani, T.; Takada, K.; et al. Activation of DNA methyltransferase 1 by EBV latent membrane protein 2A leads to promoter hypermethylation of PTEN gene in gastric carcinoma. Cancer Res. 2009, 69, 2766–2774. [Google Scholar] [CrossRef]
- Martin, K.A.; Lupey, L.N.; Tempera, I. Epstein-Barr Virus Oncoprotein LMP1 Mediates Epigenetic Changes in Host Gene Expression through PARP1. J. Virol. 2016, 90, 8520–8530. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, P.; Feng, J.; Li, R.; Chen, J.; Zheng, W.V.; Zhang, H.; Yao, P. LMP1 mediates tumorigenesis through persistent epigenetic modifications and PGC1β upregulation. Oncol. Rep. 2023, 49, 53. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-T.; Chen, Y.-R.; Chen, S.-C.; Hu, C.-Y.; Lin, C.-S.; Chang, Y.-T.; Wang, W.-B.; Chen, J.-Y. Epstein-Barr virus latent membrane protein 1 induces micronucleus formation, represses DNA repair and enhances sensitivity to DNA-damaging agents in human epithelial cells. Oncogene 2004, 23, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Sugden, B. The latent membrane protein 1 oncogene modifies B-cell physiology by regulating autophagy. Oncogene 2008, 27, 2833–2842. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Sugden, B. The LMP1 oncogene of EBV activates PERK and the unfolded protein response to drive its own synthesis. Blood 2008, 111, 2280–2289. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Shi, F.; Li, Y.; Li, W.; Yu, X.; Zhao, L.; Zhou, M.; Hu, J.; Luo, X.; Tang, M.; et al. Drp1-dependent remodeling of mitochondrial morphology triggered by EBV-LMP1 increases cisplatin resistance. Signal Transduct. Target. Ther. 2020, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Mancao, C.; Hammerschmidt, W. Epstein-Barr virus latent membrane protein 2A is a B-cell receptor mimic and essential for B-cell survival. Blood 2007, 110, 3715–3721. [Google Scholar] [CrossRef] [PubMed]
- Rancan, C.; Schirrmann, L.; Hüls, C.; Zeidler, R.; Moosmann, A. Latent Membrane Protein LMP2A Impairs Recognition of EBV-Infected Cells by CD8+ T Cells. PLOS Pathog. 2015, 11, e1004906. [Google Scholar] [CrossRef] [PubMed]
- Iwakiri, D. Epstein-Barr Virus-Encoded RNAs: Key Molecules in Viral Pathogenesis. Cancers 2014, 6, 1615–1630. [Google Scholar] [CrossRef] [PubMed]
- Rotola, A.; Ravaioli, T.; Gonelli, A.; Dewhurst, S.; Cassai, E.; Di Luca, D. U94 of human herpesvirus 6 is expressed in latently infected peripheral blood mononuclear cells and blocks viral gene expression in transformed lymphocytes in culture. Proc. Natl. Acad. Sci. USA 1998, 95, 13911–13916. [Google Scholar] [CrossRef]
- Campbell, A.; Hogestyn, J.M.; Folts, C.J.; Lopez, B.; Pröschel, C.; Mock, D.; Mayer-Pröschel, M. Expression of the Human Herpesvirus 6A Latency-Associated Transcript U94A Disrupts Human Oligodendrocyte Progenitor Migration. Sci. Rep. 2017, 7, 3978. [Google Scholar] [CrossRef]
- Caccuri, F.; Sommariva, M.; Marsico, S.; Giordano, F.; Zani, A.; Giacomini, A.; Fraefel, C.; Balsari, A.; Caruso, A. Inhibition of DNA Repair Mechanisms and Induction of Apoptosis in Triple Negative Breast Cancer Cells Expressing the Human Herpesvirus 6 U94. Cancers 2019, 11, 1006. [Google Scholar] [CrossRef] [PubMed]
- Indari, O.; Tiwari, D.; Tanwar, M.; Kumar, R.; Jha, H.C. Early biomolecular changes in brain microvascular endothelial cells under Epstein–Barr virus influence: A Raman microspectroscopic investigation. Integr. Biol. 2022, 14, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Bentz, G.L.; Jarquin-Pardo, M.; Chan, G.; Smith, M.S.; Sinzger, C.; Yurochko, A.D. Human cytomegalovirus (HCMV) infection of endothelial cells promotes naive monocyte extravasation and transfer of productive virus to enhance hematogenous dissemination of HCMV. J. Virol. 2006, 80, 11539–11555. [Google Scholar] [CrossRef] [PubMed]
- Wekerle, H. Epstein-Barr virus sparks brain autoimmunity in multiple sclerosis. Nature 2022, 603, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Kanno, H.; Watabe, D.; Shimizu, N.; Sawai, T. Adhesion of Epstein–Barr virus-positive natural killer cell lines to cultured endothelial cells stimulated with inflammatory cytokines. Clin. Exp. Immunol. 2008, 151, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Waldman, W.J.; Williams, M.V.; Lemeshow, S.; Binkley, P.; Guttridge, D.; Kiecolt-Glaser, J.K.; Knight, D.A.; Ladner, K.J.; Glaser, R. Epstein-Barr virus-encoded dUTPase enhances proinflammatory cytokine production by macrophages in contact with endothelial cells: Evidence for depression-induced atherosclerotic risk. Brain Behav. Immun. 2008, 22, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Barrett, M.M.; Williams, M.V.; Lemeshow, S.; Binkley, P.; Guttridge, D.; Kiecolt-Glaser, J.K.; Knight, D.A.; Ladner, K.J.; Glaser, R. Lymphocytic Arteritis in Epstein–Barr Virus Vulvar Ulceration (Lipschütz Disease): A Report of 7 Cases. Am. J. Dermatopathol. 2015, 37, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Indari, O.; Chandramohanadas, R.; Jha, H.C. Epstein–Barr virus infection modulates blood–brain barrier cells and its co-infection with Plasmodium falciparum induces RBC adhesion. Pathog. Dis. 2020, 79, ftaa080. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-K.; Chen, X.-R.; Wang, L.-N.; Wang, J.-H.; Li, J.-K.; Zhou, Z.-Y.; Li, X.; Cai, L.-B.; Zhong, S.-S.; Zhang, J.-J.; et al. Exosomal HMGA2 protein from EBV-positive NPC cells destroys vascular endothelial barriers and induces endothelial-to-mesenchymal transition to promote metastasis. Cancer Gene Ther. 2022, 29, 1439–1451. [Google Scholar] [CrossRef]
- Granato, M.; Santarelli, R.; Farina, A.; Gonnella, R.; Lotti, L.V.; Faggioni, A.; Cirone, M. Epstein-barr virus blocks the autophagic flux and appropriates the autophagic machinery to enhance viral replication. J. Virol. 2014, 88, 12715–12726. [Google Scholar] [CrossRef]
- Baglio, S.R.; van Eijndhoven, M.A.J.; Koppers-Lalic, D.; Berenguer, J.; Lougheed, S.M.; Gibbs, S.; Léveillé, N.; Rinkel, R.N.P.M.; Hopmans, E.S.; Swaminathan, S.; et al. Sensing of latent EBV infection through exosomal transfer of 5′pppRNA. Proc. Natl. Acad. Sci. USA 2016, 113, E587–E596. [Google Scholar] [CrossRef] [PubMed]
- McNamara, R.P.; Chugh, P.E.; Bailey, A.; Costantini, L.M.; Ma, Z.; Bigi, R.; Cheves, A.; Eason, A.B.; Landis, J.T.; Host, K.M.; et al. Extracellular vesicles from Kaposi Sarcoma-associated herpesvirus lymphoma induce long-term endothelial cell reprogramming. PLOS Pathog. 2019, 15, e1007536. [Google Scholar] [CrossRef] [PubMed]
- Takatsuka, H.; Wakae, T.; Mori, A.; Okada, M.; Fujimori, Y.; Takemoto, Y.; Okamoto, T.; Kanamaru, A.; Kakishita, E. Endothelial damage caused by cytomegalovirus and human herpesvirus-6. Bone Marrow Transplant. 2003, 31, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Hara, J.; Miyoshi, H.; Osugi, Y.; Fujisaki, H.; Takai, K.; Ohta, H.; Tanaka-Taya, K.; Yamanishi, K.; Okada, S. Thrombotic microangiopathy associated with reactivation of human herpesvirus-6 following high-dose chemotherapy with autologous bone marrow transplantation in young children. Bone Marrow Transplant. 1999, 24, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Shioda, S.; Kasai, F.; Ozawa, M.; Hirayama, N.; Satoh, M.; Kameoka, Y.; Watanabe, K.; Shimizu, N.; Tang, H.; Mori, Y.; et al. The human vascular endothelial cell line HUV-EC-C harbors the integrated HHV-6B genome which remains stable in long term culture. Cytotechnology 2018, 70, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Vallbracht, K.B.; Schwimmbeck, P.L.; Kuhl, U.; Rauch, U.; Seeberg, B.; Schultheiss, H.-P. Differential Aspects of Endothelial Function of the Coronary Microcirculation Considering Myocardial Virus Persistence, Endothelial Activation, and Myocardial Leukocyte Infiltrates. Circulation 2005, 111, 1784–1791. [Google Scholar] [CrossRef] [PubMed]
- Maximilian Buja, L. HHV-6 in Cardiovascular Pathology. In Perspectives in Medical Virology; Krueger, G., Ablashi, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 233–241. [Google Scholar]
- Kühl, U.; Lassner, D.; Wallaschek, N.; Gross, U.M.; Krueger, G.R.; Seeberg, B.; Kaufer, B.B.; Escher, F.; Poller, W.; Schultheiss, H. Chromosomally integrated human herpesvirus 6 in heart failure: Prevalence and treatment. Eur. J. Heart Fail. 2015, 17, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Comar, M.; D’Agaro, P.; Campello, C.; Poli, A.; Breinholt, J.P.; A Towbin, J.; Vatta, M. Human herpes virus 6 in archival cardiac tissues from children with idiopathic dilated cardiomyopathy or congenital heart disease. J. Clin. Pathol. 2009, 62, 80–83. [Google Scholar] [CrossRef]
- Wada, A.; Muramatsu, K.; Sunaga, Y.; Mizuno, T.; Takei, M.; Ogasawara, S.; Uchida, M.; Tsukida, K.; Tashiro, M. Brainstem infarction associated with HHV-6 infection in an infant. Brain Dev. 2018, 40, 242–246. [Google Scholar] [CrossRef]
- Brennan, Y.; Gottlieb, D.J.; Baewer, D.; Blyth, E. A fatal case of acute HHV-6 myocarditis following allogeneic haemopoietic stem cell transplantation. J. Clin. Virol. 2015, 72, 82–84. [Google Scholar] [CrossRef]
- Leveque, N.; Boulagnon, C.; Brasselet, C.; Lesaffre, F.; Boutolleau, D.; Metz, D.; Fornes, P.; Andreoletti, L. A fatal case of Human Herpesvirus 6 chronic myocarditis in an immunocompetent adult. J. Clin. Virol. 2011, 52, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Escher, F.; Escher, F.; Kuehl, U.; Lassner, D.; Poller, W.; Tschoepe, C.; Schultheiss, H.-P. Cardiac involvement of human herpesvirus 6 in patients with inflammatory cardiomyopathy. J. Am. Coll. Cardiol. 2015, 65 (Suppl. S10), A945. [Google Scholar] [CrossRef]
- Mahrholdt, H.; Wagner, A.; Deluigi, C.C.; Kispert, E.; Hager, S.; Meinhardt, G.; Vogelsberg, H.; Fritz, P.; Dippon, J.; Bock, C.T.; et al. Presentation, Patterns of Myocardial Damage, and Clinical Course of Viral Myocarditis. Circulation 2006, 114, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, H.; Miyakawa, K.; Nishida, H.; Yano, S.; Sonoda, T.; Suenobu, S.-I.; Izumi, T.; Daa, T.; Ihara, K. Unique cell tropism of HHV-6B in an infantile autopsy case of primary HHV-6B encephalitis. Neuropathology 2018, 38, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Miyake, Y.; Imoto, K.; Hattori, S.; Miyake, S.; Ishizaki, T.; Yamada, A.; Kurata, T.; Nagai, T.; Suga, S.; et al. Distribution of human herpesvirus 6 and varicella-zoster virus in organs of a fatal case with exanthem subitum and varicella. Acta Paediatr. Jpn. 1996, 38, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Harberts, E.; Yao, K.; Wohler, J.E.; Maric, D.; Ohayon, J.; Henkin, R.; Jacobson, S. Human herpesvirus-6 entry into the central nervous system through the olfactory pathway. Proc. Natl. Acad. Sci. USA 2011, 108, 13734–13739. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Asano, Y. Central nervous system complications in human herpesvirus-6 infection. Brain Dev. 2000, 22, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Eliassen, E.; Hemond, C.C.; Santoro, J.D. HHV-6-Associated Neurological Disease in Children: Epidemiologic, Clinical, Diagnostic, and Treatment Considerations. Pediatr. Neurol. 2020, 105, 10–20. [Google Scholar] [CrossRef]
- Bartolini, L.; Theodore, W.H.; Jacobson, S.; Gaillard, W.D. Infection with HHV-6 and its role in epilepsy. Epilepsy Res. 2019, 153, 34–39. [Google Scholar] [CrossRef]
- Santpere, G.; Telford, M.; Andrés-Benito, P.; Navarro, A.; Ferrer, I. The Presence of Human Herpesvirus 6 in the Brain in Health and Disease. Biomolecules 2020, 10, 1520. [Google Scholar] [CrossRef]
- Romeo, M.A.; Montani, M.S.G.; Gaeta, A.; D’Orazi, G.; Faggioni, A.; Cirone, M. HHV-6A infection dysregulates autophagy/UPR interplay increasing beta amyloid production and tau phosphorylation in astrocytoma cells as well as in primary neurons, possible molecular mechanisms linking viral infection to Alzheimer’s disease. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165647. [Google Scholar] [CrossRef] [PubMed]
- Romeo, M.A.; Santarelli, R.; Montani, M.S.G.; Gonnella, R.; Benedetti, R.; Faggioni, A.; Cirone, M. Viral Infection and Autophagy Dysregulation: The Case of HHV-6, EBV and KSHV. Cells 2020, 9, 2624. [Google Scholar] [CrossRef] [PubMed]
- Reynaud, J.M.; Jégou, J.-F.; Welsch, J.C.; Horvat, B. Human herpesvirus 6A infection in CD46 transgenic mice: Viral persistence in the brain and increased production of proinflammatory chemokines via Toll-like receptor 9. J. Virol. 2014, 88, 5421–5436. [Google Scholar] [CrossRef] [PubMed]
- Flamand, L.; Gosselin, J.; D’Addario, M.; Hiscott, J.; Ablashi, D.V.; Gallo, R.C.; Menezes, J. Human herpesvirus 6 induces interleukin-1 beta and tumor necrosis factor alpha, but not interleukin-6, in peripheral blood mononuclear cell cultures. J. Virol. 1991, 65, 5105–5110. [Google Scholar] [CrossRef] [PubMed]
- Flamand, L.; Menezes, J. Cyclic AMP-responsive element-dependent activation of Epstein-Barr virus zebra promoter by human herpesvirus 6. J. Virol. 1996, 70, 1784–1791. [Google Scholar] [CrossRef] [PubMed]
- Van Campen, C.; Rowe, P.C.; Visser, F.C. Deconditioning does not explain orthostatic intolerance in ME/CFS (myalgic encephalomyelitis/chronic fatigue syndrome). J. Transl. Med. 2021, 19, 193. [Google Scholar] [CrossRef] [PubMed]
- Van Campen, C.M.C.; Visser, F.C. Comparison of the Degree of Deconditioning in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS) Patients with and without Orthostatic Intolerance. Med. Res. Arch. 2022, 10, 2058. [Google Scholar] [CrossRef]
- Nelson, M.J.; Bahl, J.S.; Buckley, J.D.; Thomson, R.L.; Davison, K. Evidence of altered cardiac autonomic regulation in myalgic encephalomyelitis/chronic fatigue syndrome: A systematic review and meta-analysis. Medicine 2019, 98, e17600. [Google Scholar] [CrossRef]
- Vallet, B. Endothelial cell dysfunction and abnormal tissue perfusion. Crit. Care Med. 2002, 30 (Suppl. S5), S229–S234. [Google Scholar] [CrossRef]
- Levy, B.I.; Schiffrin, E.; Mourad, J.-J.; Agostini, D.; Vicaut, E.; Safar, M.E.; Struijker-Boudier, H.A. Impaired tissue perfusion: A pathology common to hypertension, obesity, and diabetes mellitus. Circulation 2008, 118, 968–976. [Google Scholar] [CrossRef]
- Bauersachs, J.; Widder, J.D. Endothelial dysfunction in heart failure. Pharmacol. Rep. 2008, 60, 119–126. [Google Scholar] [PubMed]
- Hasdai, D.; Gibbons, R.J.; HolmesJr, D.R.; Higano, S.T.; Lerman, A. Coronary Endothelial Dysfunction in Humans Is Associated With Myocardial Perfusion Defects. Circulation 1997, 96, 3390–3395. [Google Scholar] [CrossRef] [PubMed]
- Machin, D.R.; Bloom, S.I.; Campbell, R.A.; Phuong, T.T.T.; Gates, P.E.; Lesniewski, L.A.; Rondina, M.T.; Donato, A.J. Advanced age results in a diminished endothelial glycocalyx. Am. J. Physiol.-Heart Circ. Physiol. 2018, 315, H531–H539. [Google Scholar] [CrossRef] [PubMed]
- Ozbek, O.; Koç, O.; Paksoy, Y.; Aydin, K.; Nayman, A. Epstein-Barr virus encephalitis: Findings of MRI, MRS, diffusion and perfusion. Turk. J. Pediatr. 2011, 53, 680–683. [Google Scholar] [PubMed]
- Yoshinari, S.; Hamano, S.-I.; Minamitani, M.; Tanaka, M.; Eto, Y. Human Herpesvirus 6 Encephalopathy Predominantly Affecting the Frontal Lobes. Pediatr. Neurol. 2007, 36, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.-T.; Chiu, N.-C.; Shen, E.-Y.; Ho, C.-S.; Wu, M.-C. Cerebral perfusion in children with Alice in Wonderland syndrome. Pediatr. Neurol. 1998, 19, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, S.; Abdullah, S.; Shaheen, F.; Mueller, L.; Gagen, B.; Duchesne, J.; Steele, C.; Pociask, D.; Kolls, J.; Jackson-Weaver, O. Glycocalyx degradation and the endotheliopathy of viral infection. PLoS ONE 2022, 17, e0276232. [Google Scholar] [CrossRef] [PubMed]
- Wadowski, P.P.; Jilma, B.; Kopp, C.W.; Ertl, S.; Gremmel, T.; Koppensteiner, R. Glycocalyx as Possible Limiting Factor in COVID-19. Front. Immunol. 2021, 12, 607306. [Google Scholar] [CrossRef] [PubMed]
- Terasawa, M.; Hiramoto, K.; Uchida, R.; Suzuki, K. Anti-Inflammatory Activity of Orally Administered Monostroma nitidum Rhamnan Sulfate against Lipopolysaccharide-Induced Damage to Mouse Organs and Vascular Endothelium. Mar. Drugs 2022, 20, 121. [Google Scholar] [CrossRef]
- De Pasquale, V.; Quiccione, M.S.; Tafuri, S.; Avallone, L.; Pavone, L.M. Heparan Sulfate Proteoglycans in Viral Infection and Treatment: A Special Focus on SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 6574. [Google Scholar] [CrossRef]
- Lee, D.H.; Dane, M.J.C.; van den Berg, B.M.; Boels, M.G.S.; Van Teeffelen, J.W.; De Mutsert, R.; den Heijer, M.; Rosendaal, F.R.; Van der Vlag, J.; van Zonneveld, A.J.; et al. Deeper Penetration of Erythrocytes into the Endothelial Glycocalyx Is Associated with Impaired Microvascular Perfusion. PLoS ONE 2014, 9, e96477. [Google Scholar] [CrossRef] [PubMed]
- Beurskens, D.M.; Bol, M.; Delhaas, T.; van de Poll, M.C.; Reutelingsperger, C.P.; Nicolaes, G.A.; Sels, J.-W.E. Decreased endothelial glycocalyx thickness is an early predictor of mortality in sepsis. Anaesth. Intensive Care 2020, 48, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Cabrales, P.; Vázquez, B.Y.S.; Tsai, A.G.; Intaglietta, M. Microvascular and capillary perfusion following glycocalyx degradation. J. Appl. Physiol. 2007, 102, 2251–2259. [Google Scholar] [CrossRef] [PubMed]
- Chappell, D.; Westphal, M.; Jacob, M. The impact of the glycocalyx on microcirculatory oxygen distribution in critical illness. Curr. Opin. Anesthesiol. 2009, 22, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Ikonomidis, I.; Pavlidis, G.; Lambadiari, V.; Rafouli-Stergiou, P.; Makavos, G.; Thymis, J.; Kostelli, G.; Varoudi, M.; Katogiannis, K.; Theodoropoulos, K.; et al. Endothelial glycocalyx and microvascular perfusion are associated with carotid intima-media thickness and impaired myocardial deformation in psoriatic disease. J. Hum. Hypertens. 2022, 36, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Caselli, E.; Soffritti, I.; D’accolti, M.; Bortolotti, D.; Rizzo, R.; Sighinolfi, G.; Giuggioli, D.; Ferri, C. HHV-6A Infection and Systemic Sclerosis: Clues of a Possible Association. Microorganisms 2020, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Kresch, E.; Achua, J.; Saltzman, R.; Khodamoradi, K.; Arora, H.; Ibrahim, E.; Kryvenko, O.N.; Almeida, V.W.; Firdaus, F.; Hare, J.M.; et al. COVID-19 Endothelial Dysfunction Can Cause Erectile Dysfunction: Histopathological, Immunohistochemical, and Ultrastructural Study of the Human Penis. World J. Mens. Health 2021, 39, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Chimenti, C.; Verardo, R.; Grande, C.; Francone, M.; Frustaci, A. Infarct-like myocarditis with coronary vasculitis and aneurysm formation caused by Epstein–Barr virus infection. ESC Heart Fail. 2020, 7, 938–941. [Google Scholar] [CrossRef] [PubMed]
- Jamal, O.; Sahel, N.; Saouab, R.; El Qatni, M.; Zaizaa, M.; El Kassimi, I.; Rkiouak, A.; Hammi, S.; Sekkach, Y. Fatal Systemic Vasculitis Associated with Chronic Active Epstein-Barr Virus Infection. Mo. Med. 2021, 118, 226–232. [Google Scholar]
- Gatto, A.; Angelici, S.; Soligo, M.; Di Giuda, D.; Manni, L.; Curatola, A.; Ferretti, S.; Chiaretti, A. Pediatric cerebral stroke induced by Epstein-Barr virus infection: Role of Interelukin overexpression. Acta Biomed. 2021, 92 (Suppl. S1), e2021135. [Google Scholar]
- Bader, M.S.; Haroon, B.; Trahey, J.; Al-Musawi, A.; Hawboldt, J.B. Disseminated Intravascular Coagulation and Venous Thromboembolism Due to Acute Epstein-Barr Virus Infection. Infect. Dis. Clin. Pract. 2011, 19, 284–285. [Google Scholar] [CrossRef]
- O’Connor, D.S.; Elmariah, S.; Aledort, L.; Pinney, S. Disseminated Intravascular Coagulation Complicating Epstein-Barr Virus Infection in a Cardiac Transplant Recipient: A Case Report. Transplant. Proc. 2010, 42, 1973–1975. [Google Scholar] [CrossRef] [PubMed]
- Plummer, M.P.; Young, A.M.H.; O’leary, R.; Damian, M.S.; Lavinio, A. Probable Catastrophic Antiphospholipid Syndrome with Intracerebral Hemorrhage Secondary to Epstein–Barr Viral Infection. Neurocritical Care 2018, 28, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Nohgawa, M. EB virus reactivation triggers thrombotic thrombocytopenic purpura in a healthy adult. Leuk. Res. Rep. 2017, 8, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Mashav, N.; Saar, N.; Chundadze, T.; Steinvil, A.; Justo, D. Epstein-Barr virus-associated venous thromboembolism: A case report and review of the literature. Thromb. Res. 2008, 122, 570–571. [Google Scholar] [CrossRef] [PubMed]
- Hal, S.V.; Senanayake, S.; Hardiman, R. Splenic infarction due to transient antiphospholipid antibodies induced by acute Epstein-Barr virus infection. J. Clin. Virol. 2005, 32, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhao, X.; Zhu, X.; Shi, J.; Liu, L.; Wang, X.; Xie, M.; Ma, C.; Hu, Y.; Sun, J. Functional analysis and expression profile of human platelets infected by EBV in vitro. Infect. Genet. Evol. 2022, 102, 105312. [Google Scholar] [CrossRef] [PubMed]
- Winiarski, J. Antibodies to platelet membrane glycoprotein antigens in three cases of infectious mononucleosis-induced thrombocytopenic purpura. Eur. J. Haematol. 1989, 43, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Kamijo, T.; Nakazawa, Y.; Kurokawa, Y.; Sakashita, K.; Komiyama, A.; Koike, K.; Ueno, I.; Fujisawa, K. Specific autoantibodies to platelet glycoproteins in Epstein-Barr virus-associated immune thrombocytopenia. Int. J. Hematol. 2003, 78, 168–170. [Google Scholar] [CrossRef]
- Yoshimori, M.; Nishio, M.; Ohashi, A.; Tateishi, M.; Mimura, A.; Wada, N.; Saito, M.; Shimizu, N.; Imadome, K.-I.; Arai, A. Interferon-γ Produced by EBV-Positive Neoplastic NK-Cells Induces Differentiation into Macrophages and Procoagulant Activity of Monocytes, Which Leads to HLH. Cancers 2021, 13, 5097. [Google Scholar] [CrossRef]
- Belford, A.; Myles, O.; Magill, A.; Wang, J.; Myhand, R.C.; Waselenko, J.K. Thrombotic microangiopathy (TMA) and stroke due to human herpesvirus-6 (HHV-6) reactivation in an adult receiving high-dose melphalan with autologous peripheral stem cell transplantation. Am. J. Hematol. 2004, 76, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cao, Y.; Ma, L.; Pei, H.; Rausch, W.D.; Li, H. Dysfunction of Cerebrovascular Endothelial Cells: Prelude to Vascular Dementia. Front. Aging Neurosci. 2018, 10, 376. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Herrmann, N.; Dinoff, A.; Mazereeuw, G.; Oh, P.I.; Goldstein, B.I.; Kiss, A.; Shammi, P.; Lanctôt, K.L. Association Between Endothelial Function and Cognitive Performance in Patients with Coronary Artery Disease During Cardiac Rehabilitation. Psychosom. Med. 2019, 81, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Buie, J.J.; Watson, L.S.; Smith, C.J.; Sims-Robinson, C. Obesity-related cognitive impairment: The role of endothelial dysfunction. Neurobiol. Dis. 2019, 132, 104580. [Google Scholar] [CrossRef] [PubMed]
- Riedel, B.; Browne, K.; Silbert, B. Cerebral protection: Inflammation, endothelial dysfunction, and postoperative cognitive dysfunction. Curr. Opin. Anesthesiol. 2014, 27, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Groeneveld, O.N.; Berg, E.v.D.; Johansen, O.E.; Schnaidt, S.; Hermansson, K.; Zinman, B.; A Espeland, M.; Biessels, G.J. Oxidative stress and endothelial dysfunction are associated with reduced cognition in type 2 diabetes. Diab. Vasc. Dis. Res. 2019, 16, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Gozal, D.; Kheirandish-Gozal, L.; Bhattacharjee, R.; Spruyt, K. Neurocognitive and Endothelial Dysfunction in Children with Obstructive Sleep Apnea. Pediatrics 2010, 126, e1161–e1167. [Google Scholar] [CrossRef] [PubMed]
- Heringa, S.; Berg, E.v.D.; Reijmer, Y.; Nijpels, G.; Stehouwer, C.; Schalkwijk, C.; Teerlink, T.; Scheffer, P.; Hurk, K.v.D.; Kappelle, L.; et al. Markers of low-grade inflammation and endothelial dysfunction are related to reduced information processing speed and executive functioning in an older population—The Hoorn Study. Psychoneuroendocrinology 2014, 40, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.-M.; Park, J.-Y.; Seo, S.W.; Song, J. Association of plasma endothelial lipase levels on cognitive impairment. BMC Psychiatry 2019, 19, 187. [Google Scholar] [CrossRef]
- Martins-Filho, R.K.; Zotin, M.C.; Rodrigues, G.; Pontes-Neto, O. Biomarkers Related to Endothelial Dysfunction and Vascular Cognitive Impairment: A Systematic Review. Dement. Geriatr. Cogn. Disord. 2020, 49, 365–374. [Google Scholar] [CrossRef]
- Øie, M.G.; Rødø, A.S.B.; Bølgen, M.S.; Pedersen, M.; Asprusten, T.T.; Wyller, V.B.B. Subjective and objective cognitive function in adolescent with chronic fatigue following Epstein-Barr virus infection. J. Psychosom. Res. 2022, 163, 111063. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.-M.; Cheon, H.-S.; Jo, C.; Koh, Y.H.; Song, J.; Jeon, J.-P. Elevated Epstein-Barr Virus Antibody Level is Associated with Cognitive Decline in the Korean Elderly. J. Alzheimers Dis. 2017, 55, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Tarchini, G. Human Herpesviruses HHV-6A, HHV-6B & HHV-7: Diagnosis and Clinical Management. 3rd ed. Clin. Infect. Dis. 2014, 60, 496–497. [Google Scholar]
- Huang, C.; Liu, W.; Ren, X.; Lv, Y.; Wang, L.; Huang, J.; Zhu, F.; Wu, D.; Zhou, L.; Huang, X.; et al. Association between human herpesvirus 6 (HHV-6) and cognitive function in the elderly population in Shenzhen, China. Aging Clin. Exp. Res. 2022, 34, 2407–2415. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Shigihara, Y.; Funakura, M.; Kanai, E.; Watanabe, Y. Fatigue-associated alterations of cognitive function and electroencephalographic power densities. PLoS ONE 2012, 7, e34774. [Google Scholar] [CrossRef] [PubMed]
- Zerr, D.M.; Fann, J.R.; Breiger, D.; Boeckh, M.; Adler, A.L.; Xie, H.; Delaney, C.; Huang, M.-L.; Corey, L.; Leisenring, W.M. HHV-6 reactivation and its effect on delirium and cognitive functioning in hematopoietic cell transplantation recipients. Blood 2011, 117, 5243–5249. [Google Scholar] [CrossRef] [PubMed]
- Navone, S.E.; Marfia, G.; Invernici, G.; Cristini, S.; Nava, S.; Balbi, S.; Sangiorgi, S.; Ciusani, E.; Bosutti, A.; Alessandri, G.; et al. Isolation and expansion of human and mouse brain microvascular endothelial cells. Nat. Protoc. 2013, 8, 1680–1693. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Hernandez, H.; Cuevas, E.; Lantz, S.M.; Paule, M.G.; Ali, S.F. Isolation and Culture of Brain Microvascular Endothelial Cells for In Vitro Blood-Brain Barrier Studies. Methods Mol. Biol. 2018, 1727, 315–331. [Google Scholar] [PubMed]
- Bueno-Betí, C.; Novella, S.; Lázaro-Franco, M.; Pérez-Cremades, D.; Heras, M.; Sanchís, J.; Hermenegildo, C. An affordable method to obtain cultured endothelial cells from peripheral blood. J. Cell. Mol. Med. 2013, 17, 1475–1483. [Google Scholar] [CrossRef]
- Okumura, N.; Kusakabe, A.; Hirano, H.; Inoue, R.; Okazaki, Y.; Nakano, S.; Kinoshita, S.; Koizumi, N. Density-gradient centrifugation enables the purification of cultured corneal endothelial cells for cell therapy by eliminating senescent cells. Sci. Rep. 2015, 5, 15005. [Google Scholar] [CrossRef]
- Liu, R.; Li, X.; Tulpule, A.; Zhou, Y.; Scehnet, J.S.; Zhang, S.; Lee, J.-S.; Chaudhary, P.M.; Jung, J.; Gill, P.S. KSHV-induced notch components render endothelial and mural cell characteristics and cell survival. Blood 2010, 115, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.-W.; Greene, W.; Ye, F.; Gao, S.-J. Kaposi’s Sarcoma-Associated Herpesvirus Disrupts Adherens Junctions and Increases Endothelial Permeability by Inducing Degradation of VE-Cadherin. J. Virol. 2008, 82, 11902–11912. [Google Scholar] [CrossRef] [PubMed]
- Vercellotti, G.M. Effects of viral activation of the vessel wall on inflammation and thrombosis. Blood Coagul. Fibrinolysis 1998, 9 (Suppl. S2), S3–S6. [Google Scholar] [PubMed]
- Shen, Y.H.; Utama, B.; Wang, J.; Raveendran, M.; Senthil, D.; Waldman, W.; Belcher, J.; Vercellotti, G.; Martin, D.; Mitchelle, B.; et al. Human cytomegalovirus causes endothelial injury through the ataxia telangiectasia mutant and p53 DNA damage signaling pathways. Circ. Res. 2004, 94, 1310–1317. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Links between Herpesvirus Infection and Endothelial Dysfunction | References |
|---|---|
| EBV | |
| ECs infected with EBV exhibit a proinflammatory phenotype, along with NF-κB and TLR9 activation, increased interferon, cytokine, and adhesion molecule expressions, and increased clotting propensity | [66,92,137] |
| ECs increase the expressions of markers associated with vascular injury, such as endothelin-1, thrombospondin 1, and heparan sulphate proteoglycan 2 | [66] |
| Monocytes have the ability to transfer EBV infection to ECs | [138] |
| Microvascular brain ECs infected by EBV exhibit a proinflammatory phenotype and lead to leukocyte recruitment | [65,139] |
| The upregulation of endothelial adhesion marker VCAM-1 upon infection | [140] |
| EBV-infected macrophages induce proinflammatory sequelae in ECs and increase adhesion molecule expression | [141] |
| EBV dUTPase compromises the blood–brain barrier integrity | [9] |
| EBV alters cholesterol, polysaccharide, nucleotides, nucleic acid, and proline moieties in infected brain microvascular ECs | [137] |
| EBV-infected ECs of genital origin express LMP-1 on their membranes | [142] |
| The endothelial microenvironment is influenced by EBV infection | [143] |
| Extracellular vesicles from EBV-infected cells damage endothelial gap junctions and prompt endothelial-to-mesenchymal transitions | [144] |
| The modulation of host autophagy in endothelial cells | [145] |
| Exosomes containing EBV-proteins can cross brain ECs and enter the central nervous system | [146] |
| EBV protein-containing exosomes can lead to long-term endothelial dysfunction | [147] |
| HHV-6 | |
| HHV-6 infection is associated with endothelial dysfunction and a greater extent of endothelial damage than HCMV | [81,148,149] |
| HHV-6 infects ECs but does not induce cytolytic effects, which led to the conclusion that ECs act as a reservoir for HHV-6 in vivo | [67] |
| HVV-6 is able to maintain a low level of replication within ECs | [70,150] |
| An association between HHV-6 and endothelial dysfunction coupled to microcirculatory defects was demonstrated | [151] |
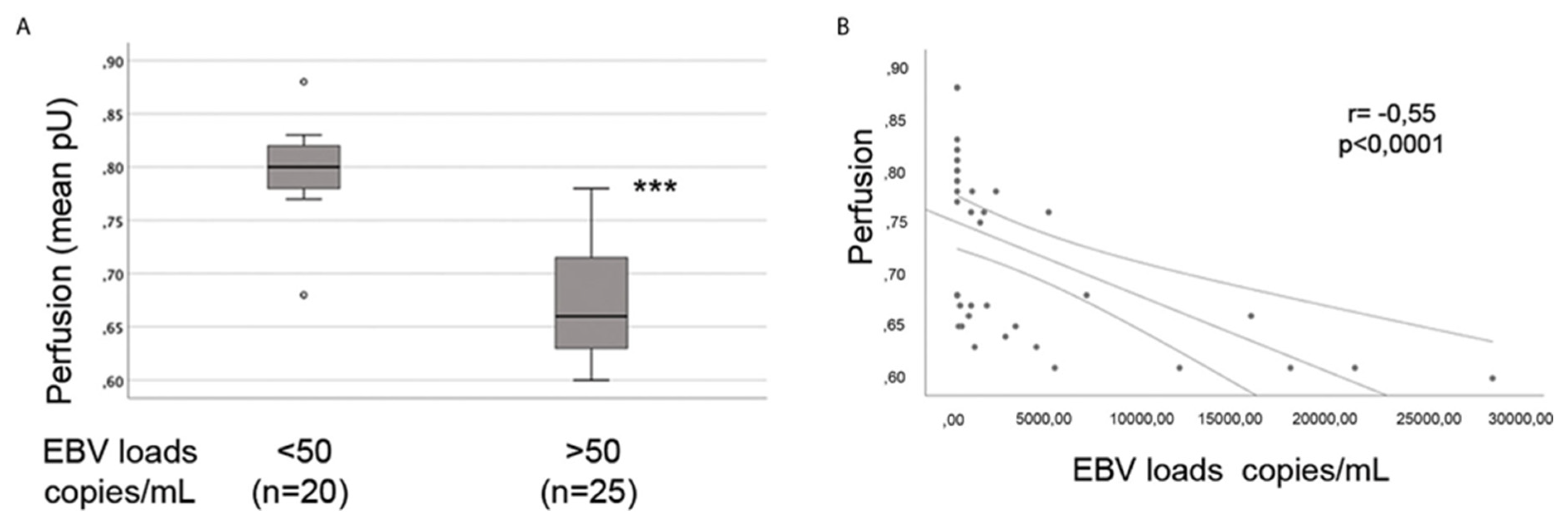
| The induction of endothelial dysfunction by HHV-6 and subsequent influence on perfusion were alluded to | [152] |
| HHV-6 antigens, DNA, and virus particles were found in ECs and associated vascular tissue from patients suffering from various cardiovascular diseases | [72,153,154,155,156,157] |
| Cardiac dysfunction, specifically reduced LVEF, is associated with HHV-6 DNA persistence in endomyocardial biopsies and is ameliorated when HHV-6 latency is resolved | [158] |
| Considered to be a major cause of viral myocarditis | [159] |
| HHV-6 also infects the CNS and ECs lining its vasculature | [160,161,162] |
| HVV-6 is implicated in neurological disease | [163,164,165,166,167,168] |
| Much like EBV, HHV-6 uses TLR9 to upregulate inflammation and promote lymphocyte filtration, as was revealed in a study where mice infected with HHV-6 subtypes resulted in CNS infection and viral persistence in brain tissue for up to 9 months | [169] |
| HHV-6 induces cellular inflammation and upregulates the expressions of IL-8, RANTES, and monocyte chemoattractant protein-1 in ECs, even in a latent state, without viral DNA replication | [67,69,170] |
| It can also promote the reactivation of EBV | [171] |
| Lymphatic ECs also succumb to latent infection by HHV-6, where EC angiogenic and migratory properties are modulated | [81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, J.M.; Kell, D.B.; Pretorius, E. Herpesvirus Infection of Endothelial Cells as a Systemic Pathological Axis in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Viruses 2024, 16, 572. https://doi.org/10.3390/v16040572
Nunes JM, Kell DB, Pretorius E. Herpesvirus Infection of Endothelial Cells as a Systemic Pathological Axis in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Viruses. 2024; 16(4):572. https://doi.org/10.3390/v16040572
Chicago/Turabian StyleNunes, Jean M., Douglas B. Kell, and Etheresia Pretorius. 2024. "Herpesvirus Infection of Endothelial Cells as a Systemic Pathological Axis in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome" Viruses 16, no. 4: 572. https://doi.org/10.3390/v16040572
APA StyleNunes, J. M., Kell, D. B., & Pretorius, E. (2024). Herpesvirus Infection of Endothelial Cells as a Systemic Pathological Axis in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Viruses, 16(4), 572. https://doi.org/10.3390/v16040572