
Viroids of the Mediterranean Basin


Abstract
1. Introduction
Viroid Acronyms



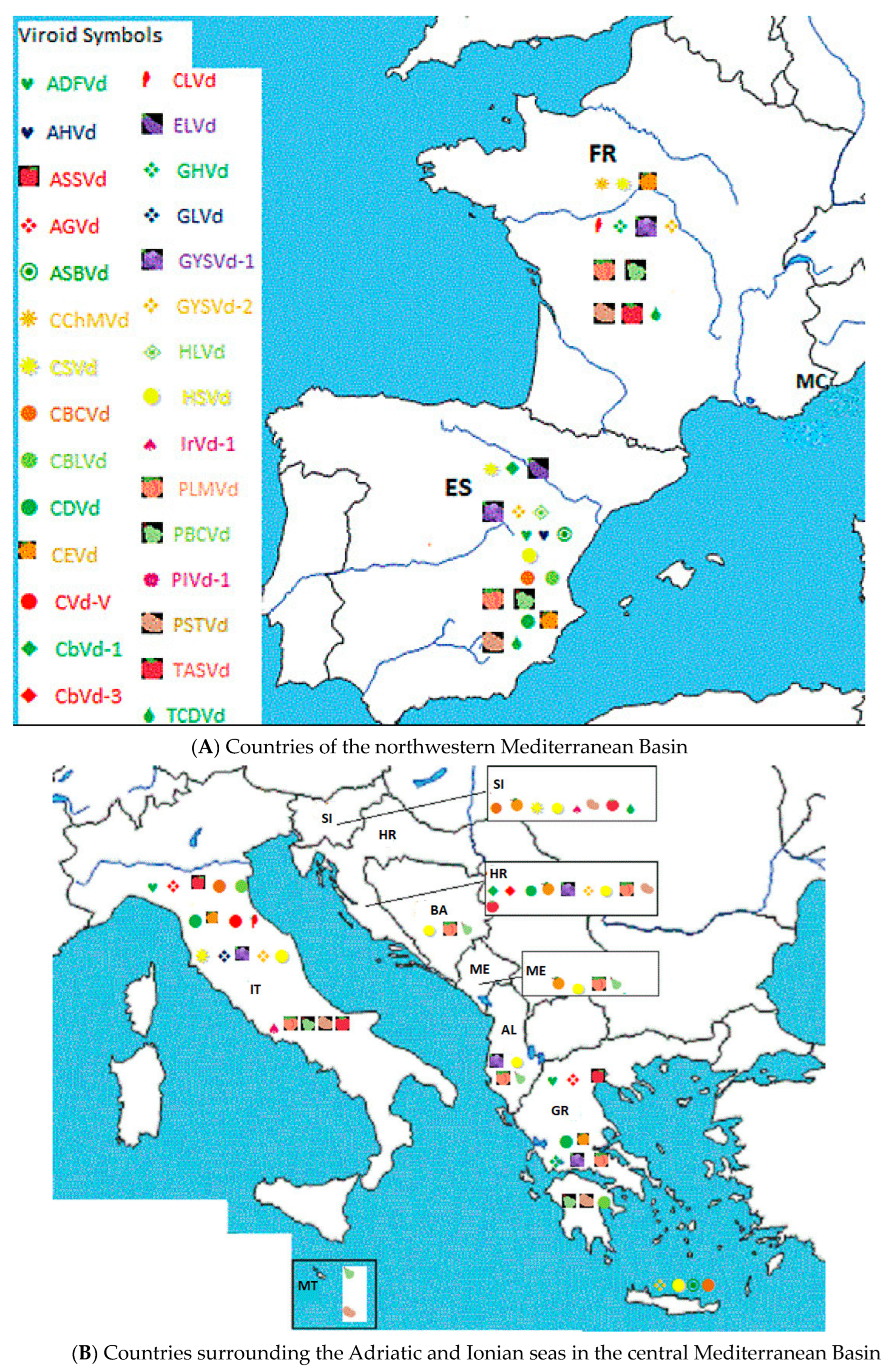
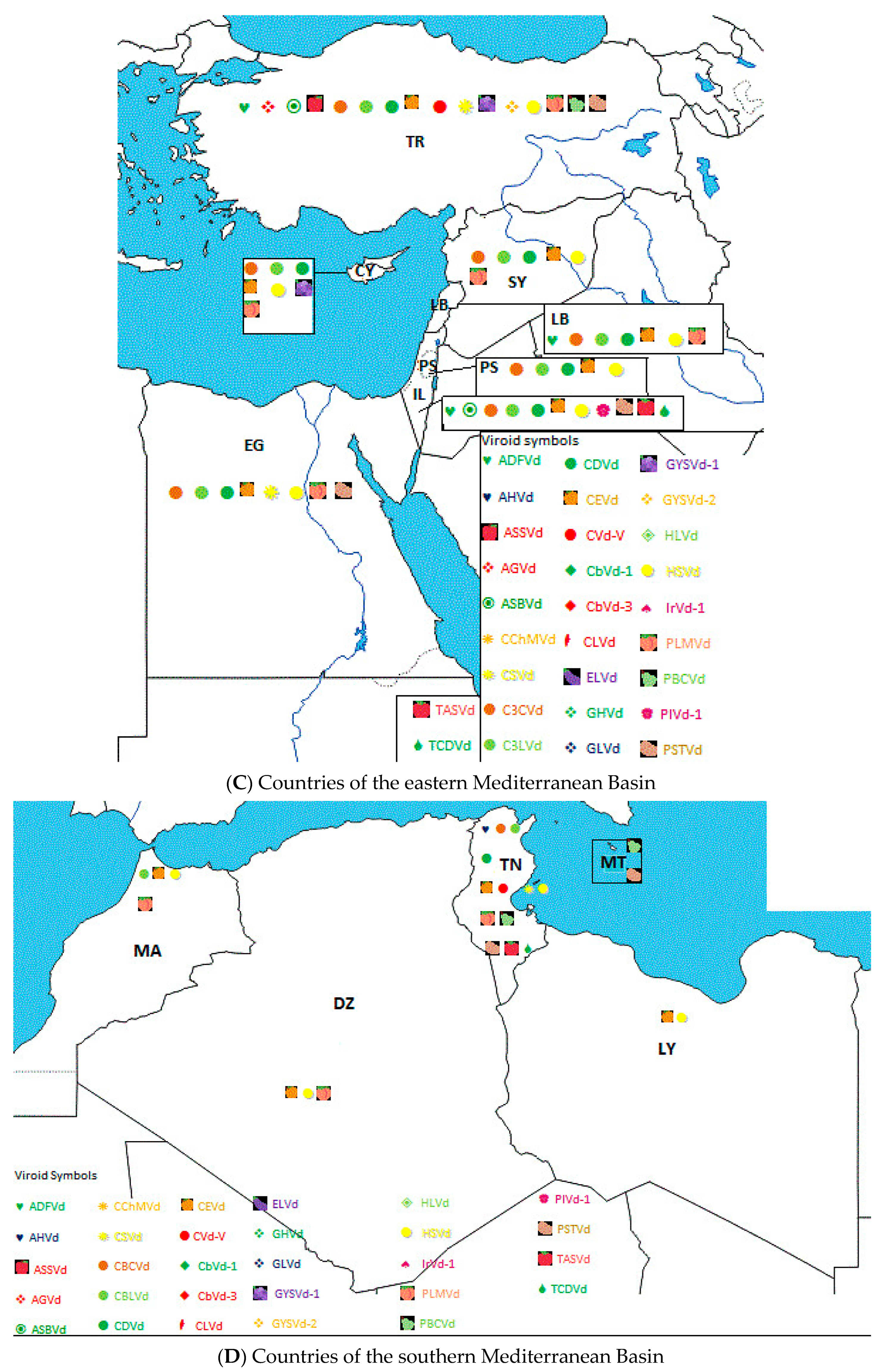
2. Geographical Distribution and Host Range of Viroids in the Mediterranean Basin
3. Viroids of Woody Plants: Fruit and Nut Trees, and Grapevine
4. Viroids of Herbaceous and Ornamental Crops
5. Viroids Not Reported in the Mediterranean Basin
6. Incidence of Viroids and Viroid Diseases in Countries of the Mediterranean Basin
6.1. Albania
6.2. Algeria
6.3. Bosnia and Herzegovina
6.4. Croatia
6.5. Cyprus
6.6. Egypt
6.7. France
6.8. Greece
6.9. Israel
6.10. Italy
6.11. Lebanon
6.12. Libya
6.13. Malta
6.14. Monaco
6.15. Montenegro
6.16. Morocco
6.17. The Palestinian Authority
6.18. Slovenia
6.19. Spain
6.20. Syria
6.21. Tunisia
6.22. Turkey
7. Origin, Evolution, and Spread of Viroids in the Mediterranean Basin
7.1. ADFVd
7.2. AHVd
7.3. AGVd
7.4. ASBVd
7.5. ASSVd
7.6. CBCVd
7.7. CBLVd
7.8. CEVd
7.9. CDVd
7.10. CVd-V
7.11. CSVd
7.12. CbVd-1, CbVd-3
7.13. GHVd
7.14. HSVd
7.15. PLMVd
7.16. PBCVd
7.17. PSTVd
7.18. TASVd
7.19. TCDVd
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Dedication
References
- The International Committee of Taxonomy on Viruses (ICTV). Current ICTV Taxonomy Release. Available online: https://ictv.global/taxonomy (accessed on 24 March 2024).
- Di Serio, F.; Li, S.-F.; Pallas, V.; Owens, R.A.; Randles, J.W.; Sano, T.; Verhoeven, J.T.J.; Vidalakis, G.; Flores, R. Viroid Taxonomy. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; pp. 135–146. [Google Scholar]
- Öksal, H.D.; Aydin, S.; Baran, M.; Emir, C.; Karanfil, A.; Bozdogan, O.; Sipahioglu, H. Persimmon (Diospyros kaki L.) and Johnsongrass [Sorghum halepense (L.) Pers.] are new natural hosts of peach latent mosaic viroid. Appl. Ecol. Environ. Res. 2021, 19, 4629–4639. [Google Scholar] [CrossRef]
- Hadidi, A.; Flores, R.; Randles, J.W.; Palukaitis, P. Viroids and Satellites; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; 716p. [Google Scholar]
- Hadidi, A. Next-generation sequencing and CRISPR/Cas 13 editing in viroid research and molecular diagnosis. Viruses 2019, 11, 120. [Google Scholar] [CrossRef] [PubMed]
- Di Serio, F.; Alioto, D.; Aloj, B.; Ragozzino, A. Infossatura crateriforme della mela: Una nuova malattia di probabile eziologia viroidale osservata in Italia meridionale. Inf. Fitopatol. 1997, 12, 14–17. [Google Scholar]
- Di Serio, F.; Alioto, D.; Ragozzino, A.; Giunchedi, L.; Flores, R. Identification of apple dimple fruit viroid in different commercial varieties of apple grown in Italy. Acta Hortic. 1998, 472, 595–602. [Google Scholar] [CrossRef]
- Di Serio, F.; Afechtal, M.; Attard, D.; Choueiri, E.; Gumus, M.; Kaymak, S.; Lolić, B.; Matić, S.; Navarro, B.; Yesilcollou, S.; et al. Detection by tissue printing hybridization of pome fruit viroids in the Mediterranean basin. Jul.-Kühn Archiv. 2010, 427, 357–360. [Google Scholar]
- Chiumenti, M.; Torchetti, E.M.; Di Serio, F.; Minafra, A. Identification and characterization of a viroid resembling apple dimple fruit viroid in fig (Ficus carica L.) by next generation sequencing of small RNAs. Virus Res. 2014, 188, 5459. [Google Scholar] [CrossRef] [PubMed]
- Choueiri, E.; El Zammar, S.; Jreijiri, F.; Hobeika, C.; Myrta, A.; Di Serio, F. First report of apple dimple fruit viroid in Lebanon. J. Plant Pathol. 2007, 89, 304. [Google Scholar]
- Ruiz-García, A.B.; Olmos, A.; Marais, A.; Faure, C.; Candresse, T. Natural Infection of Pomegranate (Punica granatum) by apple dimple fruit viroid. Cells 2023, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Sassalou, C.L.; Katis, N.I.; Maliogka, V.I. First report of apple dimple fruit viroid in fig (Ficus carica L.) in Greece. In Proceedings of the Conference on Viroids, Heraklion, Greece, 14–16 September 2022; Meeting; Abstracts volume. p. 48. [Google Scholar]
- Berbati, M.; Tzanetakis, I.; Vidalakis, G.; Voloudakis, A. Viruses and viroids in the fig core collection of the Agricultural University of Athens. In Proceedings of the Conference on Viroids, Heraklion, Greece, 14–16 September 2022; Meeting; Abstracts volume. p. 37. [Google Scholar]
- Buzkan, N.; Balsak, S.C. First report of apple dimple fruit viroid in fig (Ficus carica L.) trees in Turkey. J. Plant Pathol. 2022, 104, 897. [Google Scholar] [CrossRef]
- Zhang, Z.; Qi, S.; Tang, N.; Zhang, X.; Chen, S.; Zhu, P.; Ma, L.; Cheng, J.; Xu, Y.; Lu, M.; et al. Discovery of replicating circular RNAs by RNA-seq and computational algorithms. PLoS Pathog. 2014, 10, e1004553. [Google Scholar] [CrossRef] [PubMed]
- Szostek, S.A.; Wright, A.; Harper, S. First report of apple hammerhead viroid in the US, Japan, Italy, Spain, and New Zealand. Plant Dis. 2018, 102, 2670. [Google Scholar] [CrossRef]
- Canales, C.; Morán, F.; Olmos, A.; Ruiz-García, A.B. First detection and molecular characterization of apple stem grooving virus, apple chlorotic leaf spot virus, and apple hammerhead viroid in Loquat in Spain. Plants 2021, 10, 2293. [Google Scholar] [CrossRef] [PubMed]
- Kyriakopoulou, P.E.; Hadidi, A. Natural infection of wild and cultivated pears with apple skin viroid in Greece. Acta Hort. 1998, 472, 617–625. [Google Scholar] [CrossRef]
- Kyriakopoulou, P.E.; Giunchedi, L.; Hadidi, A. Peach latent mosaic and pome fruit viroids in naturally infected cultivated (Pyrus communis) and wild pear (P. amygdaliformis): Implications on possible origin of these viroids in the Mediterranean region. J. Plant Pathol. 2001, 83, 51–62. [Google Scholar]
- Kyriakopoulou, P.E.; Osaki, H.; Zhu, S.F.; Hadidi, A. Apple scar skin viroid in pear. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, VIC, Australia, 2003; pp. 142–145. [Google Scholar]
- Kaponi, M.S. Pome and Stone Fruit Viroids in Greece—Molecular Characterization and Phylogenetic Relationships. Ph.D. Dissertation, Agricultural University of Athens, Athens, Greece, 2009. English summary. 230p. [Google Scholar]
- Boubourakas, I.N. Peach Latent Mosaic Viroid (PLMVd) and Pome Fruit Viroids in Greece. Ph.D. Thesis, Agricultural University of Athens, Athens, Greece, 2010. English summary. 217p. [Google Scholar]
- Kaponi, M.S.; Sano, T.; Kyriakopoulou, P.E. Natural infection of sweet cherry trees with apple scar skin viroid. J. Plant Pathol. 2013, 95, 429–433. [Google Scholar]
- Sipahiŏglu, H.M.; Usta, M.; Ocak, M. Development of a rapid enzymatic cDNA amplification test for the detection of apple scar skin viroid (ASSVd) in apple trees from eastern Anatolia. Turkey. Arch. Phytopathol. Plant Protect. 2009, 42, 352–360. [Google Scholar] [CrossRef]
- Kryovrysanaki, N.; Katsarou, K.; Olmos, A.; Ruiz-García, A.B.; Kalantidis, K.; Pappi, P. First report of Australian grapevine viroid in grapevine in Greece. J. Plant Pathol. 2021, 103, 1023–1024. [Google Scholar] [CrossRef]
- Gambino, G.; Navarro, B.; Torchetti, E.M.; La Notte, P.; Schneider, A.; Mannini, F.; Di Serio, F. Survey on viroids infecting grapevine in Italy: Identification and characterization of Australian grapevine viroid and grapevine yellow speckle viroid 2. Eur. J. Plant Pathol. 2014, 140, 199–205. [Google Scholar] [CrossRef]
- Elleuch, A.; Fakhfakh, H.; Pelchat, M.; Landry, P.; Marrakchi, M.; Perreault, J.-P. Sequencing of Australian grapevine viroid and yellow speckle viroid I isolated from a Tunisian grapevine without passage in an indicator plant. Eur. J. Plant Pathol. 2002, 108, 815–820. [Google Scholar] [CrossRef]
- Çandar, A.; Öner, S.; Usta, M.; Paylan, I.C.; Sipahioglu, M.; Gümüş, M. First report of Australian grapevine viroid (AGVd) in the Aegean region vinyards of Turkey. In Proceedings of the 18th Congress of ICVG, Ankara, Turkey, 7–11 September 2015; pp. 196–197. [Google Scholar]
- Lotos, L.; Kavroulakis, N.; Navarro, B.; Di Serio, F.; Olmos, A.; Ruiz, G.A.; Katis, N.I.; Maliogka, V.I. First report of avocado sunblotch viroid (ASBVd) naturally infecting avocado (Persea americana) in Greece. Plant Dis. 2018, 102, 1470. [Google Scholar] [CrossRef]
- Spiegel, S.; Alper, M.; Allen, R.N. Evaluation of biochemical methods for the diagnosis of the avocado sunblotch viroid in Israel. Phytoparasitica 1984, 12, 37–43. [Google Scholar] [CrossRef]
- Pallás, V.; García-Luque, I.; Domingo, E.; Flores, R. Sequence variability in avocado sunblotch viroid (ASBVd). Nucleic Acids Res. 1988, 16, 9864. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Önelge, N.; Ertuǧrul, B. Detection of avocado sunblotch viroid (ASBVd) in Turkish avocado introduction material by polyacrylamide gel electrophoresis. J. Turk. Phytopathol. 1997, 26, 97–101. [Google Scholar]
- Horst, R.K. Chrysanthemum chlorotic mottle. In The Viroids; Diener, T.O., Ed.; Plenum Press: New York, NY, USA, 1987; pp. 291–295. [Google Scholar]
- El-Dougdoug, K.A.; Rezk, A.A.; Rehab, D.A.; Sofy, A.R. Partial nucleotide sequence and secondary structure of chrysanthemum stunt viroid Egyptian isolate from infected chrysanthemum plants. Int. J. Virol. 2012, 8, 191–202. [Google Scholar] [CrossRef]
- Flores, R.; Gago-Zachert, S.; Serra, P.; De la Pena, M.; Navarro, B. Chrysanthemum chlorotic mottle viroid. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; pp. 331–338. [Google Scholar]
- Faggioli, F.; Duran-Vila, N.; Tsagris, M.; Pallás, V. Geographical Distribution of Viroids in Europe. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; pp. 473–484. [Google Scholar]
- Hanani, A.; Khater, M.; Drais, M.I.; Djelouah, K. First report and molecular characterization of citrus dwarfing viroid (CDVd) and citrus bark cracking viroid (CBCVd) on Citrus volkameriana in Egypt. J. Plant Pathol. 2019, 101, 187. [Google Scholar] [CrossRef]
- Al-Harthi, A.; Al-Sadi, A.M.; Al-Saady, A.A. Potential of citrus budlings originating in the Middle East as sources of citrus viroids. Crop Prot. 2013, 48, 13–15. [Google Scholar] [CrossRef]
- Mathioudakis, M.M.; Tektonidis, N.; Karagianni, A.; Mikalef, L.; Gomez, P.; Hasiow-Jaroszewska, B. Incidence and epidemiology of citrus viroids in Greece: Role of host and cultivar in epidemiological characteristics. Viruses 2023, 15, 605. [Google Scholar] [CrossRef]
- Puchta, H.; Ramm, K.; Luckinger, R.; Hadas, R.; Bar-Joseph, M.; Sänger, H.L. Primary and secondary structure of Citrus viroid IV (CVd IV), a new chimeric viroid present in dwarfed grapefruit in Israel. Nucleic Acids Res. 1991, 19, 6640. [Google Scholar] [CrossRef] [PubMed]
- Abualrob, A. Molecular Detection of Citrus Viroids in West Bank. Master’s Thesis, Faculty of Graduate Studies, An-Najah National University, Palestine Authority, Nablus, Palestine, 2022. [Google Scholar]
- Önelge, N.; Kersting, U.; Guang, Y.; Bar Joseph, M.; Bozan, O. Nucleotide sequences of citrus viroids CVd-IIIa and CVd-IV obtained from dwarfed Meyer lemon trees grafted on sour orange. J. Plant Dis. Prot. 2000, 107, 387–391. [Google Scholar]
- Jakše, J.; Radisekb, S.; Pokorna, T.; Matousek, J.; Javornik, B. Deep-sequencing revealed citrus bark cracking viroid (CBCVd) as a highly aggressive pathogen on hop. Plant Pathol. 2015, 64, 831–842. [Google Scholar] [CrossRef]
- Ashulin, L.; Lachman, O.; Hadas, R.; Bar-Joseph, M. Nucleotide sequence of a new viroid species, citrus bent leaf viroid (CBLVd) isolated from grapefruit in Israel. Nucleic Acids Res. 1991, 19, 4767. [Google Scholar] [CrossRef] [PubMed]
- Ben Shaul, A.; Guan, Y.; Mogilner, N.; Hadas, R.; Mawassi, M.; Gafny, R.; Bar-Joseph, M. Genomic diversity among populations of two citrus viroids from different graft transmissible dwarfing complexes in Israel. Phytopathology 1995, 85, 359–364. [Google Scholar] [CrossRef]
- Hadidi, A.; Mazyad, H.M.; Madkour, M.A.; Bar-Joseph, M. Viroids in the Middle East. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, VIC, Australia, 2003; pp. 275–278. [Google Scholar]
- Škoríc, D.; Szychowski, J.A.; Krajǎcíc, M.; Semancik, J.S. Detection of citrus viroids in Croatia. In Proceedings of the 15th Conference of the International Organization of Citrus Virologists, Nicosia, Cyprus, 11–19 November 2001; Rouchouze, D., Hadjinicoli, A., Evrypidou, X., Eds.; Agricultural Research Institute: Nicosia, Cyprus, 2001; p. 148. [Google Scholar]
- Owens, R.A.; Yang, G.; Gundersen-Rindal, D.; Hammond, R.W.; Candresse, T.; Bar-Joseph, M. Both point mutation and RNA recombination contribute to the sequence diversity of citrus viroid III. Virus Genes. 2000, 20, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Elleuch, A.; Djilani Khouaja, F.; Hamdi, I.; Bsais, N.; Perreault, J.-P.; Marrakchi, M.; Fakhfakh, H. Sequence analysis of three citrus viroids infecting a single Tunisian citrus tree (Citrus reticulata, Clementine). Gen. Mol. Biol. 2006, 29, 705–710. [Google Scholar] [CrossRef]
- Duran-Vila, N. Citrus exocortis viroid. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; pp. 169–179. [Google Scholar]
- Yakoubi, S.; Elleuch, A.; Besaies, N.; Marrakchi, M.; Fakhfakh, H. First report of hop stunt viroid and citrus exocortis viroid on fig with symptoms of fig mosaic disease. J. Phytopathol. 2007, 155, 125–128. [Google Scholar] [CrossRef]
- El-Dougdoug, K.A.; Rehab, D.A.; Rezk, A.A.; Sofy, A.R. Incidence of fruit trees viroid diseases by tissue print hybridization in Egypt. Int. J. Virol. 2012, 8, 114–120. [Google Scholar] [CrossRef]
- Saade, P.; D’Onghia, A.M.; Khoury, W.; Turturo, C.; Savino, V. Surveys and certification sanitary status of citrus in Lebanon. In Proceedings of the 14th IOCV, Las Campinas, SP, Brazil, 13–18 September 2000; pp. 326–331. [Google Scholar]
- El-Dougdoug, K.; Caglayan, K.; Elleuch, A.; Al-Tuwariqi, H.; Gyamera, E.; Hadidi, A. Geographical Distribution of Viroids in Africa and the Middle East. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; pp. 485–496. [Google Scholar]
- Kyriakou, A.P. Incidence in Cyprus of citrus exocortis viroid and its mechanical transmission. Plant Pathol. 1992, 41, 20–24. [Google Scholar] [CrossRef]
- Papayiannis, L.C.; Kapari-Isaia, T. Hop stunt viroid and citrus exocortis viroid in Cyprus. Genetic analysis, characterization, host range and detection. In Proceedings of the 17th Pan-Hellenic Phytopathological Conference, Volos, Greece, 13–17 October 2014; p. 106. [Google Scholar]
- Pallás, V.; Gómez, G.; Duran-Vila, N. Viroids in Europe. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, VIC, Australia, 2003; pp. 268–274. [Google Scholar]
- Marn, M.V.; Pleško, I.M. Detection of citrus exocortis viroid in Solanum jasminoides in Slovenia. J. Plant Pathol. 2011, 93, S4.79. [Google Scholar]
- Marn, M.V.; Pleško, I.M.; Zindović, J. First report of citrus exocortis viroid in Verbena sp. in Montenegro. Plant Dis. 2012, 96, 593. [Google Scholar] [CrossRef] [PubMed]
- Luigi, M.; Luison, D.; Tomassoli, L.; Faggioli, F. Natural spread and molecular analysis of pospoviroids infecting ornamentals in Italy. J. Plant Pathol. 2011, 93, 491–495. [Google Scholar]
- Fagoaga, C.; Duran-Vila, N. Naturally occurring variants of citrus exocortis viroid in vegetable crops. Plant Pathol. 1996, 45, 45–53. [Google Scholar] [CrossRef]
- Serra, P.; Eiras, M.; Bani-Hashemian, S.M.; Murcia, N.; Kitajima, E.W.; Daros, J.A.; Flores, R.; Duran-Vila, N. Citrus viroid V: Occurrence, host range, diagnosis, and identification of new variants. Phytopathology 2008, 98, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, I.; Elleuch, A.; Bessaies, N.; Grubb, C.D.; Fakhfakh, H. First report of citrus viroid V in North Africa. J. Gen. Plant Pathol. 2015, 81, 87. [Google Scholar] [CrossRef]
- Önelge, N.; Yurtmen, M. First report of citrus viroid V in Turkey. J. Plant Pathol. 2012, 94, 88. [Google Scholar]
- Jezernik, K. Molecular Characterization of Coleus Blumei Viroids from Coleus (Solenostemon scutellarioides (L.) Codd). Diploma Thesis, Faculty of Science Repository, Department of Biology, University of Zagreb, Zagreb, Croatia, 2014. [Google Scholar]
- Škorić, D.; Černi, S.; Jezernik, K.; Butkovic, A. Molecular characterization of Coleus blumei viroids 1 and 3 in Plectranthus scutellarioides in Croatia. Eur. J. Plant Pathol. 2019, 155, 731–742. [Google Scholar] [CrossRef]
- Steyer, S.; Olivier, T.; Skelton, A.; Nixon, T.; Hobden, E. Columnea latent viroid (CLVd): First report in tomato in France. Plant Pathol. 2010, 59, 794. [Google Scholar] [CrossRef]
- Parrella, G.; Pacella, R.; Crescenzi, A. First record of columnea latent viroid (CLVd) in tomato in Italy. In Proceedings of the III International Symposium on Tomato Diseases, Ischia, Italy, 25–30 July 2010; p. 126. [Google Scholar]
- Fagoaga, C.; Duran-Vila, N. Eggplant latent. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, VIC, Australia, 2003; p. 333. [Google Scholar]
- Daros, J.-A. Eggplant latent viroid. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; pp. 339–344. [Google Scholar]
- Wu, Q.; Wang, Y.; Cao, M.; Pantaleo, V.; Burgyan, J.; Li, W.X.; Ding, S.W. Homology-independent discovery of replicating pathogenic circular RNAs by deep sequencing. Proc. Natl. Acad. Sci. USA 2012, 109, 3938–3943. [Google Scholar] [CrossRef]
- Candresse, T.; Faure, C.; Theil, S.; Spilmont, A.S.; Marais, A. First report of grapevine hammerhead viroid-like RNA infecting grapevine (Vitis vinifera) in France. Plant Dis. 2017, 101, 2155. [Google Scholar] [CrossRef]
- Pappi, P.G.; Efthimiou, K.; Lotos, L.; Katis, N.; Malliogka, V.I. First report of grapevine hammerhead viroid-like RNA (GHVd) in grapevine in Greece. J. Plant Pathol. 2020, 102, 257. [Google Scholar] [CrossRef]
- Rotunno, S.; Vaira, A.M.; Marian, D.; Schneider, A.; Raimondi, S.; Di Serio, F.; Navarro, B.; Miozzi, L. First report of grapevine latent viroid infecting grapevine (Vitis vinifera) in Italy. Plant Dis. 2018, 102, 1672–1673. [Google Scholar] [CrossRef]
- Voncina, D.; Almeida, R.P.P. Screening of some Croatian autochthonous grapevine varieties reveals a multitude of viruses, including novel ones. Arch. Virol. 2018, 163, 2239–2243. [Google Scholar] [CrossRef] [PubMed]
- Minafra, A.; Martelli, G.P.; Savino, V. Viroids of grapevine in Italy. Vitis 1990, 29, 173–182. [Google Scholar]
- Gazel, M.; Önelge, N. First report of grapevine viroids in the east Mediterranean region of Turkey. Plant Pathol. 2003, 52, 405. [Google Scholar] [CrossRef]
- Önelge, N.; Gazel, M. Detection of the viroids of grapevine in East Mediterranean Region. J. Turk. Phytopathol. 2001, 30, 70. [Google Scholar]
- Sassalou, C.L.; Katsarou, K.; Lotos, L.; Orfanidou, G.; Maliogka, V.I.; Kalantidis, K.; Katis, N.I.; Pappi, P.G. First report of grapevine yellow speckle viroid-2 in grapevine in Greece. Plant Dis. 2020, 104, 1879. [Google Scholar] [CrossRef]
- Pallás, V.; Navarro, A.; Flores, R. Isolation of a viroid-like RNA from hop different from hop stunt viroid. J. Gen. Virol. 1987, 68, 2095–2102. [Google Scholar] [CrossRef]
- Reanwarakorn, K.; Semancik, J.S. Correlation of hop stunt viroid variants to cachexia and xyloporosis diseases of citrus. Phytopathology 1999, 89, 568–574. [Google Scholar] [CrossRef]
- Önelge, N.; Semancik, J.S. Citrus gummy bark. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, VIC, Australia, 2003; pp. 330–332. [Google Scholar]
- El-Dougdoug, K.A.; Osman, M.E.; Hayam, S.A.; Rehab, D.A.; Elbaz, R.M. Biological and molecular detection of HSVd-infecting peach and pear trees in Egypt. Austr. J. Basic Appl. Sci. 2010, 4, 19–26. [Google Scholar]
- Nasr-Eldin, M.A.; El-Dougdoug, K.A.; Othman, B.A.; Ahmed, S.A.; Abdel-Aziz, S.H. Three viroids frequency naturally infecting grapevine in Egypt. Int. J. Virol. 2012, 8, 1–13. [Google Scholar] [CrossRef]
- Sofy, A.R.; Mousa, A.A.; Soliman, A.M.; El-Dougdoug, K.A. The limiting of climatic factors and predicting of suitable habitat for citrus gummy bark disease occurrence using GIS. Int. J. Virol. 2012, 8, 165–177. [Google Scholar] [CrossRef]
- Sofy, A.R.; El-Dougdoug, K.A. First record of a hop stunt viroid variant associated with gumming and stem pitting on Citrus volkameriana trunk rootstock in Egypt. New Dis. Rep. 2014, 30, 11. [Google Scholar] [CrossRef]
- Matíc, S.; Al-Rwahnih, M.; Myrta, A. Occurrence of stone fruit viroids in Bosnia and Herzegovina. Phytopathol. Mediterr. 2005, 44, 285–290. [Google Scholar]
- Loconsole, G.; Onelge, N.; Yokomi, R.; Abou Kubaa, R.; Savino, V.; Saponari, M. Rapid differentiation of citrus hop stunt viroid variants by real-time RT-PCR and high-resolution melting analysis. Mol. Cell. Probes 2013, 27, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Amari, K.; Cañizares, M.C.; Myrta, A.; Sabanadzovic, S.; Srhiri, M.; Gavriel, I.; Caglayan, K.; Varveri, C.; Gatt, M.; Di Terlizzi, B.; et al. First report of hop stunt viroid (HSVd) from Some Mediterranean Countries. Phytopathol. Mediterr. 2000, 39, 271–276. [Google Scholar]
- Kaponi, M.S.; Kyriakopoulou, P.E. First Report of hop stunt viroid infecting Japanese plum, cherry plum, and peach in Greece. Plant Dis. 2013, 97, 1662. [Google Scholar] [CrossRef] [PubMed]
- Kaponi, M.; Kyriakopoulou, P.E.; Luigi, M.; Barba, M.; Faggioli, F.; Israellides, K. Hop stunt viroid (HSVd), a new pathogen of stone and pome fruit trees in Greece. Phytopathol. Mediterr. 2019, 48, 308–309. [Google Scholar]
- Choueiri, E.; Abou Ghanem-Sabanadzovic, N.; Khazzaka, K.; Sabanadzovic, S.; Di Terlizzi, B.; Myrta, A.; El-Zammar, S.; Jreijiri, F.; Savino, V.F. First record of hop stunt viroid on apricot in Lebanon. J. Plant Pathol. 2002, 84, 69. [Google Scholar]
- El Beaino, T.; Choueiri, E.; Digiaro, M. First report of hop stunt viroid in Lebanese fig trees. J. Plant Pathol. 2013, 95, 218. [Google Scholar]
- El Beaino, T.; Abou Kubaa, R.; Choueiri, E.; Digiaro, M.; Navarro, B. Occurrence of hop stunt viroid in Mulberry (Morus alba) in Lebanon and Italy. J. Phytopathol. 2012, 160, 48–51. [Google Scholar] [CrossRef]
- Ismaeil, F.; Abou Ghanem-Sabanadzovic, N.; Myrta, A.; Di Terlizzi, B.; Savino, V. First record of peach latent mosaic viroid and hop stunt viroid in Syria. J. Plant Pathol. 2001, 82, 227. [Google Scholar]
- El Beaino, T.; Abou Kubaa, R.; Ismaeil, E.; Mando, J.; Digiaro, M. Viruses and hop stunt viroid of fig trees in Syria. J. Plant Pathol. 2012, 94, 687–691. [Google Scholar]
- Fekih Hassen, I.; Roussel, S.; Kummert, J.; Fakhfakh, H.; Marrakchi, M.; Jijakli, M.H. Development of a rapid RT-PCR test for the detection of peach latent mosaic viroid, pear blister canker viroid, hop stunt viroid and apple scar skin viroid in fruit trees from Tunisia. J. Phytopathol. 2006, 154, 217–223. [Google Scholar] [CrossRef]
- Gorsane, F.; Elleuch, A.; Hamdi, I.; Salhi-Hannachi, A.; Fakhfakh, H. Molecular detection and characterization of hop stunt viroid sequence variants from naturally infected pomegranate (Punica granatum L.) in Tunisia. Phytopathol. Mediterr. 2010, 49, 152–162. [Google Scholar]
- Önelge, N. Occurrence of hop stunt viroid (HSVd) on pomegranate (Punica granatum) trees in Turkey. J. Turk. Phytopathol. 2000, 29, 49–52. [Google Scholar]
- Gazel, M.; Ulubaş, C.; Çağlayan, K. Detection of hop stunt viroid in sweet and sour cherry trees in Turkey by RT-PCR. Acta Hortic. 2008, 795, 955–958. [Google Scholar] [CrossRef]
- Bostan, S.C.; Buzkan, N. Detection and genetic diversity of hop stunt viroid infecting pistachio (Pistacia vera L.) trees in Turkey. Eur. J. Plant Pathol. 2021, 159, 707. [Google Scholar]
- Sorrentino, R.; Torchetti, E.M.; Minutolo, M.; Di Serio, F.; Alioto, D. First report of iresine viroid 1 in ornamental plants in Italy and of Celosia cristata as a novel natural host. Plant Dis. 2015, 99, 1655. [Google Scholar] [CrossRef]
- Marn, V.M.; Pleško, M.I. First report of iresine viroid 1 in Portulaca sp. in Slovenia. J. Plant Pathol. 2012, 94, S4.85–S4.105. [Google Scholar]
- Skoric, D.; Al Rwahnih, M.; Myrta, A. First record of peach latent mosaic viroid in Croatia. Acta Hortic. 2008, 781, 535–539. [Google Scholar]
- Zindović, J.; Musić, M.S.; Škorić, D. Comparison of PLMVd Isolates from Montenegro and Croatia. In Proceedings of the Viroid Meeting, Heraklion, Greece, 14–16 September 2022; Abstracts volume; Poster–22. p. 57. [Google Scholar]
- Flores, R.; Navarro, B.; Delgado, S.; Hernandez, C.; Xu, W.-X.; Barba, M.; Hadidi, A.; Di Serio, F. Peach latent mosaic viroid in infected peach. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; pp. 307–316. [Google Scholar]
- Kyriakopoulou, P.E.; Giunchedi, L.; Barba, M.; Boubourakas, I.N.; Kaponi, M.S.; Hadidi, A. Peach latent mosaic viroid in temperate fruit crops other than peach. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; pp. 317–329. [Google Scholar]
- Kyriakopoulou, P.E.; Hadidi, A.; Dougdoug, K.; Giunchedi, L. Natural infection of wild and cultivated pears in Greece with peach latent mosaic viroid and pome fruit viroids. In Proceedings of the 7th International Congress of Plant Pathology (ICPP 98), Edinburgh, UK, 1–31 July 1998. paper number 3.7.49. [Google Scholar]
- Shamloul, A.; Minafra, A.; Hadidi, A.; Giunchedi, L.; Waterworth, H.; Allam, E. PLMVd: Nucleotide sequence of an Italian isolate, sensitive detection using RT-PCR and geographic distribution. Acta Hortic. 1995, 386, 522–530. [Google Scholar] [CrossRef]
- Hadidi, A.; Giunchedi, L.; Shamloul, A.M.; Poggi-Pollini, G.; Amer, M.A. Occurrence of peach latent mosaic viroid in stone fruits and its transmission with contaminated blades. Plant Dis. 1997, 81, 154–158. [Google Scholar] [CrossRef]
- Giunchedi, L.; Gentit, P.; Nemchinov, L.; Poggi-Pollini, G.; Hadidi, A. Plum spotted fruit: A disease associated with peach latent mosaic viroid. Acta Hortic. 1998, 472, 571–580. [Google Scholar] [CrossRef]
- Faggioli, F.; Loreti, S.; Barba, M. Occurrence of peach latent mosaic viroid (PLMVd) on plum in Italy. Plant Dis. 1997, 81, 423. [Google Scholar] [CrossRef] [PubMed]
- Faggioli, F.; Barba, M. Peach latent mosaic viroid: Major findings of our studies over a period of fifteen years in Italy. Acta Hortic. 2008, 781, 529–534. [Google Scholar] [CrossRef]
- Crescenzi, A.; Piazzolla, P.; Hadidi, A. First report of peach latent mosaic viroid in sweet cherry in Italy. J. Plant Pathol. 2002, 84, 168. [Google Scholar]
- Diekman, M.; Putter, C.A.J. FAO/IPGRI Technical Guidelines for Safe Movement of Germplasm. N. 16. Stone Fruits; Food and Agriculture Organization of the United Nations/International Plant Genetic Resources Institute: Rome, Italy, 1996. [Google Scholar]
- Fekih Hassen, I.; Massart, S.; Motard, J.; Roussel, S.; Parisi, O.; Kummert, J.; Fakhfakh, H.; Marrakchi, M.; Perreault, J.-P.; Jijakli, M.H. Molecular features of new peach latent mosaic viroid variants suggest that recombination may have contributed to the evolution of this infectious RNA. Virology 2007, 360, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Sipahiŏglu, H.M.; Demir, S.; Myrta, A.; Al Rwahnih, M.; Polat, B.; Schena, L.; Usta, M.; Akkopru, A.; Selcuk, M.; Ippolito, A.; et al. Viroid, phytoplasma and fungal diseases of stone fruit in eastern Anatolia, Turkey. N. Z. J. Crop Hort. 2006, 34, 1–6. [Google Scholar] [CrossRef]
- Gümüs, M.; Paylan, I.C.; Matíc, S.; Myrta, A.; Sipahiŏglu, H.M.; Erkan, S. Occurrence and distribution of stone fruit viruses and viroids in commercial plantings of Prunus species in Western Anatolia. Turk. J. Plant Pathol. 2007, 89, 265–268. [Google Scholar]
- Navarro, B.; Bacu, A.; Torchetti, M.; Kongjika, E.; Susuri, L.; Di Serio, F.; Myrta, A. First record of pear blister canker viroid on pear in Albania. J. Plant Pathol. 2011, 93, S4.70. [Google Scholar]
- Lolic, B.; Afechtal, M.; Matíc, S.; Myrta, A.; Di Serio, F. Detection by tissue-printing of pome fruit viroids and characterization of pear blister canker viroid in Bosnia and Herzegovina. J. Plant Pathol. 2007, 89, 369–375. [Google Scholar]
- Ambrós, S.; Desvignes, J.C.; Llácer, G.; Flores, R. Peach latent mosaic viroid and pear blister canker viroids: Detection by molecular hybridization and relationships with specifics maladies affecting peach and pear trees. Acta Hortic. 1995, 386, 515–521. [Google Scholar] [CrossRef]
- Hernández, C.; Elena, S.F.; Moga, A.; Flores, R. Pear blister canker viroid is a member of ASSVd subgroup andalso has sequence homology with viroids from other subgroups. J. Gen. Virol. 1992, 73, 2503–2507. [Google Scholar] [CrossRef] [PubMed]
- Attard, D.; Afechtal, M.; Agius, M.; Matic, S.; Gatt, M.; Myrta, A.; Di Serio, F. First report of pear blister canker viroid in Malta. J. Plant Pathol. 2007, 89, S71. [Google Scholar]
- Boubourakas, I.N.; Arambatzis, C.; Dovas, C.; Kyriakopoulou, P.E. Amelioration of a reverse transcription polymerase chain reaction (RT-PCR) for the detection of ASSVd, PBCVd and PLMVd viroids, and their presence in cultivated and wild pome and stone fruits in Greece. Acta Hortic. 2008, 781, 519–527. [Google Scholar] [CrossRef]
- Kaponi, M.S.; Luigi, M.; Barba, M.; Kyriakopoulou, P.E. Molecular characterization of Hellenic variants of apple scar skin viroid and pear blister canker viroid in pome fruit trees. Jul.-Kühn-Archiv. 2010, 427, 266–272. [Google Scholar]
- Kaponi, M.S.; Luigi, M.; Kyriakopoulou, P.E. Mixed infections of pome and stone fruit viroids in cultivated and wild trees in Greece. New Dis. Rep. 2012, 26, 8. [Google Scholar] [CrossRef]
- Yesilcollou, S.; Minoia, S.; Torchetti, E.M.; Kaymak, S.; Gumus, M.; Myrta, A.; Navarro, B.; Di Serio, F. Molecular characterization of Turkish isolates of pear blister canker viroid and assessment of the sequence variability of this viroid. J. Plant Pathol. 2010, 92, 813–819. [Google Scholar]
- Bester, R.; Malan, S.S.; Maree, H.J. A plum marbling conundrum: Identification of a new viroid associated with marbling and corky flesh in Japanese plums. Phytopathology 2020, 110, 1476–1482. [Google Scholar] [CrossRef] [PubMed]
- El-Dougdoug, K.A. Studies on Some Viroids in Egypt. Ph.D. Thesis, Ain Shams University, Cairo, Egypt, 1988; 185p. [Google Scholar]
- Bostan, H.; Gazel, M.; Elibuyuk, I.O.; Caglayan, K. Occurrence of pospiviroid in potato, tomato, and some ornamental plants in Turkey. Afr. J. Biotechnol. 2010, 9, 2613–2617. [Google Scholar]
- Güner, Ü.; Sipahioğlu, H.M.; Usta, M. Incidence and genetic stability of potato spindle tuber pospiviroid in potato in Turkey. Turk. J. Agric. For. 2012, 36, 9. [Google Scholar] [CrossRef]
- Navarro, B.; Silletti, M.R.; Trisciuzzi, V.N.; Di Serio, F. Identification, and characterization of potato spindle tuber viroid infecting tomato in Italy. J. Plant Pathol. 2009, 91, 723–726. [Google Scholar]
- Milanović, J.; Kajić, V.; Milhaljević, S. Occurrence and molecular variability of potato spindle tuber viroid and tomato apical stunt viroid in ornamental plants in Croatia. Eur. J. Plant Pathol. 2014, 139, 785–788. [Google Scholar] [CrossRef]
- Malandraki, I.; Papachristopoulou, M.; Vassilakos, N. First report of potato spindle tuber viroid (PSTVd) in ornamental plants in Greece. New Dis. Rep. 2010, 21, 9. [Google Scholar] [CrossRef]
- Marn, M.V.; Pleško, I.M.; Stare, B.G. Variability of Potato spindle tuber viroid isolates from ornamental hosts in Slovenia. J. Plant Pathol. 2013, 95, 411–415. [Google Scholar]
- Verhoeven, J.; Jansen, C.; Roenhorst, J. First Report of tomato apical stunt viroid in tomato in Tunisia. Plant Dis. 2006, 90, 528. [Google Scholar] [CrossRef] [PubMed]
- Marn, M.V.; Pleško, I.M. First report of tomato apical stunt viroid in Solanum jasminoides in Slovenia. New Dis. Rep. 2012, 26, 7. [Google Scholar] [CrossRef]
- Candresse, T.; Marais, A.; Tassus, X.; Suhard, P.; Renaudin, I.; Leguay, A.; Poliakoff, F.; Blancard, D. First report of tomato chlorotic dwarf viroid in tomato in France. Plant Dis. 2010, 94, 633. [Google Scholar] [CrossRef] [PubMed]
- Marn, M.V.; Pleško, I.M. First report of tomato chlorotic dwarf viroid in Petunia spp. in Slovenia. Plant Dis. 2010, 94, 1171. [Google Scholar] [CrossRef] [PubMed]
- Gramazio, P.; Lerma, M.D.; Villanueva, G.; Vilanova, S.; García-Fortea, E.; Mangino, G.; Figàs, M.R.; Arrones, A.; Alonso, D.; San Bautista, A.; et al. Detection, molecular characterization and aspects involving the transmission of tomato chlorotic dwarf viroid in eggplant. Ann. Appl. Biol. 2019, 175, 172–183. [Google Scholar] [CrossRef]
- Di Serio, F.; Izadpanah, K.; Hajizadeh, M.; Navarro, B. Viroids infecting the grapevine. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G., Golino, D., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 373–392. [Google Scholar]
- Sano, T.; Hataya, T.; Terai, Y.; Shikata, E. Hop stunt viroid strains from dapple fruit disease of plum and peach in Japan. J. Gen. Virol. 1989, 70, 1311–1319. [Google Scholar] [CrossRef]
- Amari, K.; Ruiz, D.; Gómez, G.; Sánchez-Pina, M.A.; Pallás, V.; Egea, J. An important new apricot disease in Spain is associated with hop stunt viroid infection. Eur. J. Plant Pathol. 2007, 118, 173–181. [Google Scholar] [CrossRef]
- Roistacher, C.N. Diagnosis and management of virus and virus like diseases of citrus. In Diseases of Fruits and Vegetables; Naqvi, S.A.M.H., Ed.; Springer: Dordrecht, The Netherlands, 2004; Volume I, pp. 109–189. [Google Scholar]
- EPPO. PQR Database; European and Mediterranean Plant Protection Organization: Paris, France, 2014; Available online: http://www.eppo.int/DATABASES/pqr/pqr.htm (accessed on 6 March 2024).
- Flores, R.; Randles, J.W.; Owens, R.A. Classification. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, VIC, Australia, 2003; pp. 71–75. [Google Scholar]
- Chiumenti, M.; Navarro, B.; Candresse, T.; Flores, R.; Di Serio, F. Reassessing species demarcation criteria in viroid taxonomy by pairwise identity matrices. Virus Evol. 2021, 7, 1–14. [Google Scholar] [CrossRef]
- EPPO. EPPO Global Database; European and Mediterranean Plant Protection Organization: Paris, France, 2023; Available online: https://gd.eppo.int/ (accessed on 19 February 2024).
- Torres, H.; Gómez, G.; Pallás, V.; Stamo, B.; Shalaby, A.; Aouane, B.; Gavriel, I.; Kominek, P.; Caglayan, K.; Sipahioglu, M.; et al. Detection by tissue printing of stone fruit viroids, from Europe, the Mediterranean and North and South America. Acta Hortic. 2004, 657, 379–383. [Google Scholar] [CrossRef]
- Rouag, N.; Guechi, A.; Matic, S.; Myrta, A. Viruses and viroids of stone fruits in Algeria. J. Plant Pathol. 2008, 90, 391–393. [Google Scholar]
- El-Ferran, A.; Belkahla, H.; Bouras, N.; Atoui, A.; Bakli, M.; Thermoz, J.-P.; Chatillon, C.; Boutoumi, H. Application of real time PCR for the prevalence determination of citrus exocortisviroid (CEVd) in Algeria. Adv. Res.Life Sci. 2023, 7, 68–72. [Google Scholar] [CrossRef]
- Škorić, D.; Černi, S.; Ćurković-Perica, M.; Ježić, M.; Krajačić, M.; Šeruga Musić, M. Legacy of plant virology in Croatia- from virus identification to molecular epidemiology, evolution, genomics and beyond. Viruses 2021, 13, 2339. [Google Scholar] [CrossRef] [PubMed]
- Plant Protection and Quarantine (PPQ). Cooperative Agricultural Pest Survey (CAPS)-Pest Datasheet for Pospiviroid Potato Spindle Tuber Viroid (Pospiviroidae): Potato Spindle Tuber; United States Department of Agriculture, Animal and Plant Health Inspection Service, Plant Protection and Quarantine (PPQ): Raleigh, NC, USA, 2022.
- EPPO. Potato Spindle Tuber Viroid (PSTVd); European and Mediterranean Plant Protection Organization: Paris, France, 2022; Available online: https://gd.eppo.int/taxon/PSTVD0 (accessed on 19 February 2024).
- CABI & EPPO. Grapevine Yellow Speckle Viroid 2. [Distribution Map]. In Distribution Maps of Plant Diseases; CABI: Wallingford, UK; EPPO: Paris, France, 2022. [Google Scholar] [CrossRef]
- Kyriakou, A.; Ioannou, M.; Hadjinicolis, A.; Hoffman, R.; Antoniou, E.; Papayiannis, L.; Kapari, T.; Ioannou, N. Citrus exocortis and cachexia viroids affect growth, yield and fruit quality of Lapithou lemon on sour orange rootstock in Cyprus. In International Organization of Citrus Virologists Conference Proceedings (1957–2010); Sixteenth IOCV Conference; IOCV: Riverside, CA, USA, 2005; Volume 16. [Google Scholar] [CrossRef]
- Papayiannis, L.C.; Kapari-Isaia, T. Viroids: The microscopic and dynamic pathogens of plants. In EnimerotikoDeltio 6; Agricultural Research Institute Publications: Nicosia, Cyprus, 2015; 28p. (In Greek) [Google Scholar]
- Kapari, T.; Voloudakis, A.; Kyriakou, A.; Ioannides, I.; Papayiannis, L.; Samouel, S.; Koutsioumari, E.; Georgiou, A.; Minas, G. Sanitation of citrus varieties and/or clones by in vitro micrografting in Cyprus and Greece. Acta Hortic. 2011, 892, 279–285. [Google Scholar] [CrossRef]
- CABI & EPPO. Citrus bark cracking viroid. [Distribution map]. In Distribution Maps of Plant Diseases; CABI: Wallingford, UK; EPPO: Paris, France, 2019. [Google Scholar] [CrossRef]
- CABI & EPPO. Citrus bent leaf viroid. [Distribution map]. In Distribution Maps of Plant Diseases; CABI: Wallingford, UK; EPPO: Paris, France, 2019. [Google Scholar] [CrossRef]
- CABI & EPPO. Citrus dwarfing viroid. [Distribution map]. In Distribution Maps of Plant Diseases; CABI: Wallingford, UK; EPPO: Paris, France, 2021. [Google Scholar] [CrossRef]
- Desvignes, J.C. Les maladies à virus du poirier et leur detection. CTILF Doc. 1970, 26, 1–12. [Google Scholar]
- Desvignes, J.C.; Boye, R.; Cornaggia, D.; Grasseau, N. Maladies à Virus des Arbres Fruitiers (Virus Diseases of Fruit Trees); CTIFL (Centre Technique Interprofessionnel des Fruits et Légumes): Paris, France, 1999; pp. 54–58. [Google Scholar]
- Marais, A.; Faure, C.; Deogratias, J.-M.; Candresse, T. First report of chrysanthemum stunt viroid in various cultivars of Argyranthemum frutescens in France. Plant Dis. 2011, 95, 1196. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Plant Health (PLH). Scientific opinion on the risk to plant health posed by chrysanthemum stunt viroid for the EU territory, with identification and evaluation of risk reduction options. EFSA J. 2012, 10, 3027. [Google Scholar]
- EFSA Panel on Plant Health (PLH). Scientific opinion on the assessment of the risk of solanaceous pospiviroids for the EU territory and the identification and evaluation of risk management options. EFSA J. 2011, 9, 2330. [Google Scholar] [CrossRef]
- EPPO Reporting Service (NPPO of France 2013-10). First Report of Tomato Apical Stunt Viroid in France; EPPO: Paris, France, 2013; Number of article 2013/236 no. 11–2013. [Google Scholar]
- Kyriakopoulou, P.E.; Tzortzakaki, S.; Tsagris, M. Grapevine asteroid mosaic in Greece: Positive indexing results and viroids associated. In Proceedings of the Extended Abstracts 11th Meeting ICVG, Montreux, Switzerland, 6–9 September 1993; Volume 41. [Google Scholar]
- Maliogka, V.I.; Olmos, A.; Pappi, P.G.; Lotos, L.; Efthimiou, K.; Grammatikaki, G.; Candresse, T.; Katis, N.I.; Avgelis, A.D. A novel grapevine badnavirus is associated with the Roditis leaf discoloration disease. Virus Res. 2015, 203, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Protopapadakis, E.; Tzortzakaki, S.; Kasapakis, J.; Tsagris, E.M. A New Graft-transmissible disease of Bergamot in Greece. In International Organization of Citrus Virologists Conference Proceedings (1957–2010); IOCV: Riverside, CA, USA, 2002; Volume 15. [Google Scholar] [CrossRef]
- Barbarossa, L.; Loconsole, G.; Vovlas, C. Virus and virus-like diseases of citrus in Epirus. J. Plant Pathol. 2007, 89, 273–276. [Google Scholar]
- Boubourakas, I.; Voloudakis, A.; Agorastou, T.; Magripis, G.; Kyriakopoulou, P.; Vidalakis, G. Pilot survey of citrus mother trees in Greece for the presence of viruses and viroids. In International Organization of Citrus Virologists Conference Proceedings (1957–2010); IOCV: Riverside, CA, USA, 2010; Volume 17. [Google Scholar]
- Wang, J.; Boubourakas, I.N.; Voloudakis, A.E.; Agorastou, T.; Magripis, G.; Rucker, T.L.; Kyriakopoulou, P.E.; Vidalakis, G. Identification and characterization of known and novel viroid variants in the Greek national citrus germplasm collection: Threats to the industry. Eur. J. Plant Pathol. 2013, 137, 17–27. [Google Scholar] [CrossRef]
- Duran-Vila, N.; Semancik, J.S. Citrus viroids. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, VIC, Australia, 2003; pp. 178–194. [Google Scholar]
- Tektonidis, N.; Mikalef, L.; Karagianni, A.; Mathioudakis, M.M. Epidemiology and molecular characterization of citrusviroids in Greece. In Proceedings of the Viroids 2022 Meeting, Heraklion, Greece, 14–16 September 2022; Abstracts volume. p. 51, Poster–16. [Google Scholar]
- Katsarou, K.; Chiumenti, M.; Kalantidis, K.; Mathioudakis, M.M. First report of citrus viroids infecting Persian (Tahiti) lime in Greece. Plant Dis. 2020, 104, 998. [Google Scholar] [CrossRef]
- Kaponi, M.S.; Vellios, E.K.; Filippou, K.S.; Sano, T. Viral pathogens occurring in fig-mosaic diseased trees in Greece. PSJ Plant Virus Dis. Workshop Rep. (Spec. Ed.) 2015, 12, 30–42. [Google Scholar]
- Ambrós, S.; Hernández, C.; Desvignes, J.C.; Flores, R. Genomic structure of three phenotypically different isolates of peach latent mosaic viroid: Implications of the existence of constraints limiting the heterogeneity of viroid quasispecies. J. Virol. 1998, 72, 7397–7406. [Google Scholar] [CrossRef] [PubMed]
- Amari, K.; Gómez, G.; Myrta, A.; Di Terlizzi, B.; Pallás, V. The molecular characterization of 16 new sequence variants of hop stunt viroid reveals the existence of invariable regions and a conserved hammerhead like structure on the viroid molecule. J. Gen. Virol. 2001, 82, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Boubourakas, I.N.; Faggioli, F.; Fucuta, S.; Barba, M.; Kyriakopoulou, P.E. The presence of peach latent mosaic (PLMVd) in Greece. Phytopathol. Mediterr. 2009, 48, 321. [Google Scholar]
- Kaponi, M. (Benaki Phytopathological Institute (BPI), Kifissia, Greece). Unpublished data.
- Bar-Joseph, M. Citrus viroids and citrus dwarfing in Israel. Acta Hortic. 1993, 349, 271–276. [Google Scholar] [CrossRef]
- Önelge, N. Nucleotide sequence of CVd-Ib and CVd-IV viroids collected from citrus gummy bark-diseased sweet orange trees in the east Mediterranean region of Turkey. J. Plant Dis. Protec. 1996, 103, 482–487. [Google Scholar]
- Oppenheimer, C. Growing New Subtropical Fruit Trees in Israel; Hassadeh Library: Tel Aviv, Israel, 1955; pp. 110–113. (In Hebrew) [Google Scholar]
- Blickle, W.; Saenger, H.L. National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/nuccore/X52045.1 (accessed on 7 February 2024).
- Anonymous. Risk Management Proposal: Solanum lycopersicum (Tomato) Seed Forsowing, from All Countries; Ministry for Primary Industries: Auckland, New Zealand, 2012; 17p.
- CPC-CABI. Crop Protection Compendium CAB International; CABI: Wallingford, UK, 2010. [Google Scholar]
- Licciardello, G.; Ferraro, R.; Scuderi, G.; Marcella, R.; Catara, A. A simulation of the use of high throughput sequencing as pre-screening assay to enhance the serveillance of citrus viruses and viroids in the EPPO Region. Agriculture 2021, 11, 400. [Google Scholar] [CrossRef]
- Bar-Joseph, M. Xyloporosis: A history of the emergence and eradication of a citrus viroid disease. J. Cit. Pathol. 2015, 2. [Google Scholar] [CrossRef]
- Antignus, Y.; Lachman, O.; Pearlsman, M.; Gofman, R.; Bar-Joseph, M. A new disease of greenhouse tomatoes in Israel caused by a distinct strain of tomato apical stunt viroid (TASVd). Phytoparasitica 2002, 30, 502–510. [Google Scholar] [CrossRef]
- Antignus, Y.; Lachman, O.; Pearlsman, M. Spread of tomato apical stunt viroid (TASVd) in greenhouse tomato crops is associated with seed transmission and bumble bee activity. Plant Dis. 2007, 91, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Dilworth, D.A. Tomato chlorotic dwarf viroid in the ornamental plant Vinca minor and its transmission throughtomato seed. Eur. J. Plant Pathol. 2009, 123, 111–116. [Google Scholar] [CrossRef]
- Owens, R.A.; Verhoeven, J.T.J. Potato spindle tuber viroid. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press/Elsevier: London, UK; Oxford, UK; San Diego, CA, USA; Cambridge, MA, USA, 2017; pp. 149–158. [Google Scholar]
- Ferraro, R.; Scuderi, G.; Russo, M.; Catara, A.; Licciardello, G. Metagenomic detection of viruses and viroids associated with isolates of citrus tristeza virus. In Proceedings of the XXIV Congress of the Italian Phytopathological Society, Department of Agricultural, Food and Environmental Sciences, Marche Polytechnic University, Ancona, Italy, 5–7 September 2018; Volume 100, pp. 613–653. [Google Scholar]
- Tessitori, M.; Rizza, S.; Reina, A.; Causarano, G.; Di Serio, F. The genetic diversity of citrus dwarfing viroid populations is mainly dependent on the infected host species. J. Gen. Virol. 2013, 94, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, L.; Faggioli, F.; Ragozzino, E.; Pasquini, G.; Barba, M. Diffusione di virus e viroidi in germoplasma di drupaceenell’Italiacentro-meridionale. In Proceedings of the Atti Progetto POM A32, Locorotondo, Italy, 4–7 December 2001; Volume II, pp. 667–673. [Google Scholar]
- Loreti, S.; Faggioli, F.; Barrale, R.; Barba, M. Occurrence of viroids in temperature fruit trees in Italy. Acta Hortic. 1998, 472, 555–560. [Google Scholar] [CrossRef]
- Flores, R.; Duran-Vila, N.; Pallás, V.; Semancik, J.S. Detection of viroid and viroid-like RNAs from grapevine. J. Gen. Virol. 1985, 66, 2095–2102. [Google Scholar] [CrossRef]
- Torchetti, E.; Navarro, B.; Trisciuzzi, V.N.; Nuccitelli, L.; Silletti, M.R.; Di Serio, F. First report of chrysanthemum stunt viroid in Argyranthemum frutescens in Italy. J. Plant Pathol. 2012, 94, 451–454. [Google Scholar]
- Abou Ghanem-Sabanadzovic, N.; Choueiri, E. Presence of peach latent mosaic and hop stunt viroid in Lebanon. In Virus and Virus-Like Diseases of Stone Fruits, with Particular Reference to the Mediterranean Region; Options Méditerranéennes: Série B. Etudes et Recherches; n. 45; Myrta, A., Di Terlizzi, B., Savino, V., Eds.; CIHEAM: Bari, Italy, 2003; pp. 139–141. [Google Scholar]
- Choueiri, E.; Abou Ghanem-Sabanadzovic, N.; Khazzaka, K.; Sabanadzovic, S.; Di Terlizzi, B.; Jreijiri, F.; Martelli, G.P. First record of peach latent mosaic viroid in Lebanon. J. Plant Pathol. 2001, 83, 225–227. [Google Scholar]
- Pleško, I.M.; Marn, M.V.; Miladinovic, Z.; Zindovic, J. First report of peach latent mosaic viroid in peach trees in Montenegro. Plant Dis. 2012, 96, 150. [Google Scholar] [CrossRef] [PubMed]
- Luigi, M.; Zindovic, J.; Stojanovic, I.; Faggioli, F. First report of potato spindle tuber viroid in Montenegro. J. Plant Pathol. 2016, 98, 184. [Google Scholar]
- Bouani, A.; Al-Rwahnih, M.; Abou-Ghanem Sabanadzovic, N.A.; Alami, I.; Zemzami, M.; Myrta, A.; Savino, V. A preliminary account of the sanitary status of stone fruit trees in Morocco. Bull. OEPP 2004, 34, 399–402. [Google Scholar] [CrossRef]
- Afechtal, M.; Jamai, H.; Mokrini, F.; Essarioui, A.; Faddoul, Z.; Sbaghi, M.; Dababat, A. First Report of hop stunt viroid infecting citrus trees in Morocco. Plant Dis. 2016, 100, 10. [Google Scholar] [CrossRef]
- Afechtal, M.; Kharmach, E.; Bibi, I. Survey, and molecular characterization of hop stunt viroid (HSVd) sequence variants from citrus groves in Morocco. Moroc. J. Agric. Sci. 2020, 1, 145–148. [Google Scholar]
- Bibi, I.; Afechtal, M.; Chafik, Z.; Bamouh, A.; Benyazid, J.; Bousamid, A.; Kharmach, E. Occurrence and distribution of virus and virus-like diseases of citrus in North-East of Morocco—Moulouya perimeter. In Proceedings of the Onzième Congrès de l’Association Marocaine de Protection des Plantes, Rabat, Marocco, 26–27 March 2019; pp. 119–134. [Google Scholar]
- Bibi, I.; Kharmach, E.; Chafik, Z.; Benyazid, J.; Bousamid, A. Incidence of citrus exocortis viroid and hopstund viroid in commercial citrus groves from Morocco. Moroc. J. Agric. Sci. 2020, 1, 142–144. [Google Scholar]
- Smith, I.M.; McNamara, D.G.; Scott, P.R.; Harris, K.M. (Eds.) Quarantine pests for Europe. In Data Sheets on Quarantine Pests for the European Communities and for the European and Mediterranean Plant Protection Organization; CAB International: Wallingford, UK; European and Mediterranean Plant Protection Organization: Paris, France, 1992. [Google Scholar]
- Ouantar, M.; Chebli, B.; Afechtal, M. First report of citrus bent leaf viroid from Morocco. Australas. Plant Dis. Notes 2018, 13, 35. [Google Scholar] [CrossRef]
- Abualrob, A.; Alabdallah, O.; Abou Kubaa, R.; Naser, S.; Alkowni, R. Molecular detection of citrus exocortis viroid (CEVd), Citrus viroid-III (CVd-III), and citrus viroid-IV (CVd-IV) in Palestine. Sci. Rep. 2024, 14, 423. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, M.H.; Treiber, C.; Born, U.; Schrader, G.; Stampfl, J.; Jakše, J.; Radišek, S. Risk potential of international fruit trade for viroid spreading—Case study on hop viroids in Europe. J. Plant Pathol. 2023, 105, 1335–1346. [Google Scholar] [CrossRef]
- Mehle, N.; Seljak, G.; Verhoeven, J.; Jansen, C.; Prezelj, N.; Ravnikar, M. Chrysanthemum stunt viroid newly reported in Slovenia. Plant Pathol. 2010, 59, 1159. [Google Scholar] [CrossRef]
- Marn, M.V.; Plěsko, I.M. First report of potato spindle tuber viroid in cape gooseberry in Slovenia. Plant Dis. 2012, 96, 150. [Google Scholar] [CrossRef] [PubMed]
- Badenes, M.L.; Llácer, G. Occurrence of peach latent mosaic viroid in American peach and nectarine cultivars. Acta Hortic. 1998, 472, 565–570. [Google Scholar] [CrossRef]
- Rubio-Cabetas, M.J.; Montañés, M.; Alonso, J.M.; Pallás, V.; Martínez, G.; Gómez, G. Incidence of peach latent mosaic viroid (PLMVd) in a Prunus persica bastch germplasm collection in Spain. Acta Hortic. 2012, 940, 687–692. [Google Scholar] [CrossRef]
- Cañizares, M.C.; Marcos, J.F.; Pallás, V. Studies on the incidence of hop stunt viroid in apricottrees (Prunus armeniaca) by using an easy and short extraction method to analyze a large number of samples. Acta Hortic. 1998, 472, 581–585. [Google Scholar] [CrossRef]
- Duran-Vila, N.; Romero-Durban, J.; Hernandez, M. Detection and eradication of chrysanthemum stunt viroid in Spain. EPPO Bull. 1996, 26, 399–405. [Google Scholar] [CrossRef]
- Vernière, C.; Perrier, X.; Dubois, C.; Dubois, A.; Botella, L.; Chabrier, C.; Bové, J.M.; Duran-Vila, N. Interactions between citrus viroids affect symptom expression and field performance of clementine trees grafted on trifoliate orange. Phytopathology 2006, 96, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Fagoaga, C.; Semancik, J.S.; Duran-Vila, N. A citrus exocortis viroid variant from broad bean (Vicia faba L.): Infectivity and pathogenesis. J. Gen. Virol. 1995, 76, 2271–2277. [Google Scholar] [CrossRef]
- Duran-Vila, N. (IVIA, Valencian Institute for Agricultural Research, Valencia, Spain). Unpublished results.
- Ismaeil, F.; Mando, J. Viruses, viroids and phytoplasmas reported to infect fruit trees in Syria: A Review. Arab J. Plant Protec. 2022, 40, 78–88. [Google Scholar]
- Abou Kubaa, R.; Saponari, M.; El-Khateeb, A.; Djelouah, K. First identification of citrus exocortis viroid (CEVd) and citrus dwarf viroid (CVd-III) in citrus orchards in Syria. J. Plant Pathol. 2016, 98, 171–185. [Google Scholar]
- Abou Kubaa, R.; El-Khateeb, A.; D’Onghia, A.M.; Djelouah, K. First report of hop stunt viroid infecting citrus orchards in Syria. J. Plant Pathol. 2011, 93 (Suppl. S4), S67. [Google Scholar]
- Najar, A.; Hamdi, I.; Varsani, A.; Duran-Vila, N. Citrus viroids in Tunisia: Prevalence and molecular characterization. J. Plant Pathol. 2017, 99, 787–792. [Google Scholar]
- Najar, A.; Duran-Vila, N. Viroid prevalence in Tunisian citrus. Plant Dis. 2004, 88, 1286. [Google Scholar] [CrossRef] [PubMed]
- Fekih Hassen, I.; Kummert, J.; Marbot, S.; Fakhfakh, H.; Marrakchi, M.; Jijakli, M. First Report of pear blister canker viroid, peach latent mosaic viroid, and hop stunt viroid infecting fruit trees in Tunisia. Plant Dis. 2004, 88, 1164. [Google Scholar] [CrossRef]
- Elleuch, A.; Hamdi, I.; Ellouze, O.; Ghrab, M.; Fakhfakh, H.; Darira, N. Pistachio (Pistacia vera L.) is a new natural host of hop stunt viroid. Virus Genes 2013, 47, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, I.; Soltani, R.; Baraket, G.; Varsani, A.; Najar, A. First report of apple hammerhead viroid infecting ‘Richared Delicious’ apple (Malus domestica) in Tunisia. J. Plant Pathol. 2022, 104, 811–812. [Google Scholar] [CrossRef]
- Elleuch, A.; Marrakchi, M.; Perreault, J.-P.; Fakhfakh, H. First report of Australian grapevine viroid from the Mediterranean region. J. Plant Pathol. 2003, 85, 51–55. [Google Scholar]
- Tuncel, F.; Tekkaş, N.; Türk, G.; Oksal, H.D.; Sipahioğlu, H.M. First detection and molecular characterization of peach latent mosaic viroid in a new natural host: Walnut (Juglans regia L.). Alinteri J. Agric. Sci. 2020, 35, 37–44. [Google Scholar]
- Gazel, M.; Ulubaş Serçe, Ç.; Çăglayan, K.; Luigi, M.; Faggioli, F. Incidence and genetic diversity of peach latent mosaic viroid ısolates in Turkey. Turk. J. Plant Pathol. 2008, 90, 495–503. [Google Scholar]
- Önelge, N.; Çinar, A.; Szychowski, J.A.; Vidalakis, G.; Semancik, J.S. Citrus viroid II variants associated with “Gummy Bark” disease. Eur. J. Plant Pathol. 2004, 110, 1047–1052. [Google Scholar] [CrossRef]
- Moreira, S. Report to Government of Turkey on Virus Diseases of Citrus; FAO/UNDP Report, No: 1982; Food and Agriculture Organization of the United Nations (FAO/UNDP): Rome, Italy, 1965; Volume 19. [Google Scholar]
- Gökçek, B. Investigation of Grape Yellow SPECKLE (GYSVd-1 and -2) Disease in Gaziantep. Ph.D. Dissertation, Çukurova University, Science Institute, Adana, Turkey, 2007; 94p. [Google Scholar]
- Gökçek, B.; Önelge, B. Gaziantep ili Bağlarında bağsarıbenek (GYSVd-1 ve 2) hastalığınınaraştırılması. In Proceedings of the Türkiye II Bitki Koruma Kongresi, Isparta, Turkey, 27–29 August 2007; p. 118. [Google Scholar]
- Önder, S.; Ozturk Gokce, N.; Ulubaş Serçe, Ç. Diagnosis of viruses and viroids via next generation sequencing in local grapevine, Turkey. In Proceedings of the Workshop on the Use of NGS Technologies for Plant Pest Diagnostics, Abstracts of Oral Communications and Posters, Bari, Italy, 23 November 2017; pp. 18, 22. [Google Scholar]
- Önelge, N.; Çınar, A. Virus and virus-like diseases in Turkey. Citriculture. In Proceedings of the 17th Conference of International Organization of Citrus Virologists (IOCV); IOCV: Riverside, CA, USA, 2010. Short Communications. pp. 233–236. [Google Scholar]
- Diener, T.O. Circular RNAs: Relics of precellular evolution? Proc. Natl. Acad. Sci. USA 1989, 86, 9370–9374. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.; Pelchat, M. Origin of hepatitis δ virus. Future Microbiol. 2010, 5, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Di Serio, F.; Navarro, B.; Flores, R. Origin and evolution of viroids. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press: Oxford, UK, 2017; pp. 125–134. [Google Scholar]
- Lee, B.D.; Neri, U.; Roux, S.; Gophna, U.; Dolja, V.V.; Koonin, E.V. Mining metatranscriptomes reveals a vast world of viroid-like circular RNAs. Cell 2023, 186, 646–661. [Google Scholar] [CrossRef] [PubMed]
- Forgia, M.; Navarro, B.; Daghino, S.; Cervera, A.; Gisel, A.; Perotto, S.; Aghayeva, D.N.; Akinyuwa, M.F.; Gobbi, E.; Zheludev, I.N.; et al. Hybrids of RNA viruses and viroid-like elements replicate in fungi. Nat. Commun. 2023, 14, 2591. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Wei, S.; Bian, R.; Luo, J.; Khan, H.A.; Tai, H.; Kondo, H.; Hadidi, A.; Andika, I.B.; Sun, L. Natural cross-kingdom spread of apple scar skin viroid from apple trees to fungi. Cells 2022, 11, 3686. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, A.; Sun, L.; Randles, J.W. Modes of viroid transmission. Cells 2022, 11, 719. [Google Scholar] [CrossRef]
- Sun, L.; Hadidi, A. Mycoviroids: Fungi as hosts and vectors of viroids. Cells 2022, 11, 1335. [Google Scholar] [CrossRef] [PubMed]
- Plants of the World Online. Royal Botanic Gardens, Kew. Available online: https://powo.science.kew.org (accessed on 6 March 2024).
- Guy, P.L. Ancient RNAs? RT-PCR of 50-year-old RNA identifies peach latent mosaic viroid. Arvh. Virol. 2013, 158, 691–694. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viroid Acronym | Country | Host Range | Reference | |
|---|---|---|---|---|
| ADFVd | Italy | apple, fig | [6,7,8,9] | |
| Lebanon | apple | [10] | ||
| Israel, Spain | pomegranate | [11] | ||
| Greece, Turkey | fig | [12,13,14] | ||
| AHVd | Italy, Spain | apple, loquat | [15,16,17] | |
| ASSVd | Greece | apple, sweet cherry, pear, wild pear, wild apple | [18,19,20,21,22,23] | |
| Italy | pear | [19] | ||
| Turkey | apple | [24] | ||
| AGVd | Greece, Italy, Tunisia, Turkey | grapevine | [25,26,27,28] | |
| ASBVd | Greece, Israel, Spain, Turkey | avocado | [29,30,31,32] | |
| CChMVd | France | chrysanthemum | [33] | |
| CSVd | Egypt, Turkey | chrysanthemum | [34,35] | |
| Slovenia, Spain | mainly on chrysanthemum but also on other ornamentals | [36] | ||
| France, Italy | chrysanthemum, marguerite daisy, common ragwort, common periwinkle, petunia, trailing petunia, climbing nightshade | [36] | ||
| CBCVd (CVd-IV) | Egypt, Greece, Israel, The Palestinian Authority, Turkey | citrus | [37,38,39,40,41,42] | |
| Slovenia | hop | [43] | ||
| CBLVd (CVd-I) | Egypt, Greece, Israel, Lebanon, The Palestinian Authority, Syria | citrus | [38,39,41,44,45,46] | |
| CDVd (CVd-III) | Croatia, Egypt, Greece, Israel, Lebanon, The Palestinian Authority, Syria, Tunisia, Turkey | citrus | [37,38,39,41,42,46,47,48,49] | |
| CEVd | Algeria, Egypt, Israel, Lebanon, Libya, Morocco, The Palestinian Authority, Syria, Tunisia, Turkey | citrus, tomato, fig | [38,41,45,46,49,50,51,52,53,54] | |
| Prevalent in Europe | citrus, climbing nightshade | [36,39,47,55,56,57,58] | ||
| Montenegro | common verbena | [59] | ||
| Italy | night blooming jessamine, Paraguay nightshade | [60] | ||
| Spain | turnip, carrot, eggplant | [61] | ||
| Italy, Spain, Turkey | grapevine | [36] | ||
| CVd-V | Spain, Tunisia, Turkey | citrus | [62,63,64] | |
| CbVd-1 | Croatia, Spain | spurflower | [36,65,66] | |
| CbVd-3 | Croatia | spurflower | [36,66] | |
| CLVd | France, Italy | tomato | [36,67,68] | |
| ELVd | Spain | eggplant | [36,69,70] | |
| GHVd | France, Italy, Greece | grapevine | [71,72,73] | |
| GLVd | Italy | grapevine | [15,74] | |
| GYSVd-1 | Albania, Croatia, Cyprus, France, Greece, Italy, Spain, Tunisia, Turkey | grapevine | [19,20,36,46,54,57,75,76,77] | |
| GYSVd-2 | Greece, Italy, Turkey | grapevine | [26,76,77,78,79] | |
| HLVd | France, Spain | hop | [36,57,80] | |
| HSVd | Countries of the Mediterranean Basin | citrus, grapevine | [36,46,54,57,81,82] | |
| Egypt | apple, pear, plum, peach, apricot, citrus, grapevine, mango | [54,83,84,85,86] | ||
| Croatia, Bosnia and Herzegovina, Slovenia, | apricot, plum, grapevine | [47,87] | ||
| Albania, Algeria, Cyprus, France, Libya, Italy, Israel, Spain | apricot, plum, peach, citrus, grapevine | [36,54,56,88,89] | ||
| Greece | almond, wild almond, plum, sweet cherry, Japanese plum, peach, apricot, apple, wild apple, grapevine, citrus | [36,89,90,91] | ||
| Lebanon | apricot, fig, mulberry, citrus, grapevine | [54,92,93,94] | ||
| Syria | fig, citrus, apricot, grapevine | [54,88,95,96] | ||
| Tunisia | pomegranate, citrus, fig, pistachio, almond, peach, pear, grapevine | [54,97,98] | ||
| Turkey | pistachio, citrus, pomegranate, grapevine, apricot, plum | [54,88,89,99,100,101] | ||
| IrVd-1 | Italy | featherhead amaranth, cockscomb | [36,102] | |
| Italy, Slovenia | common purslane | [36,103] | ||
| PLMVd | Widespread in Europe | peach | [36,57] | |
| Bosnia and Herzegovina, Croatia | peach | [87,104,105] | ||
| Egypt | apple, apricot, mango, plum, pear, peach | [46,52,54] | ||
| France, Algeria | apricot, sweet cherry, plum, peach | [106,107] | ||
| Albania | apricot, plum, peach | [107] | ||
| Greece | peach, wild pear, pear, hawthorn, plum, apricot, cherry, Japanese plum, quince | [19,22,36,106,107,108] | ||
| Italy | apricot, cherry, Japanese plum, plum, Nanjing cherry, pear | [36,57,106,107,109,110,111,112,113,114] | ||
| Lebanon, Morocco, Syria | apricot, sweet cherry, stone fruits | [46,54,95,106,107,115] | ||
| Slovenia | plum | [107] | ||
| Tunisia | almond, peach, pear | [54,97,106,107,116] | ||
| Turkey | apricot, peach | [54,117,118] | ||
| PBCVd | Albania, Bosnia and Herzegovina, France, Spain, Italy, Malta | pear | [36,119,120,121,122,123] | |
| Greece | apple, pear, quince, wild pear, wild apple | [19,124,125,126] | ||
| Tunisia, Turkey | pear, quince | [54,97,127] | ||
| PlVd-I | Israel | pomegranate | [11,128] | |
| PSTVd | Egypt, Israel, Turkey | potato, tomato | [35,46,54,129,130,131] | |
| Italy | tomato | [36,132] | ||
| Prevalent in Europe | ornamentals | [36,133,134,135] | ||
| TASVd | Israel, Tunisia | tomato | [36,46,54,136] | |
| Croatia, France, Italy, Slovenia | angel trumpets, climbing nightshade, Paraguay nightshade, night blooming jessamine, marmalade bush | [36,133,137] | ||
| France, Israel, Italy | tomato | [36] | ||
| TCDVd | France, Israel, Tunisia | tomato | [36,54,138] | |
| Tunisia | mock orange | [54] | ||
| Slovenia | petunia | [36,139] | ||
| Spain | eggplant | [140] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaponi, M.; Kyriakopoulou, P.E.; Hadidi, A. Viroids of the Mediterranean Basin. Viruses 2024, 16, 612. https://doi.org/10.3390/v16040612
Kaponi M, Kyriakopoulou PE, Hadidi A. Viroids of the Mediterranean Basin. Viruses. 2024; 16(4):612. https://doi.org/10.3390/v16040612
Chicago/Turabian StyleKaponi, Maria, Panayota E. Kyriakopoulou, and Ahmed Hadidi. 2024. "Viroids of the Mediterranean Basin" Viruses 16, no. 4: 612. https://doi.org/10.3390/v16040612
APA StyleKaponi, M., Kyriakopoulou, P. E., & Hadidi, A. (2024). Viroids of the Mediterranean Basin. Viruses, 16(4), 612. https://doi.org/10.3390/v16040612